
High Purity of α-Lactalbumin from Binary Protein Mixture by Charged UF Membrane Far from the Isoelectric Point to Limit Fouling

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Protein Quantification
2.3. Protein Size and Charge Measurement
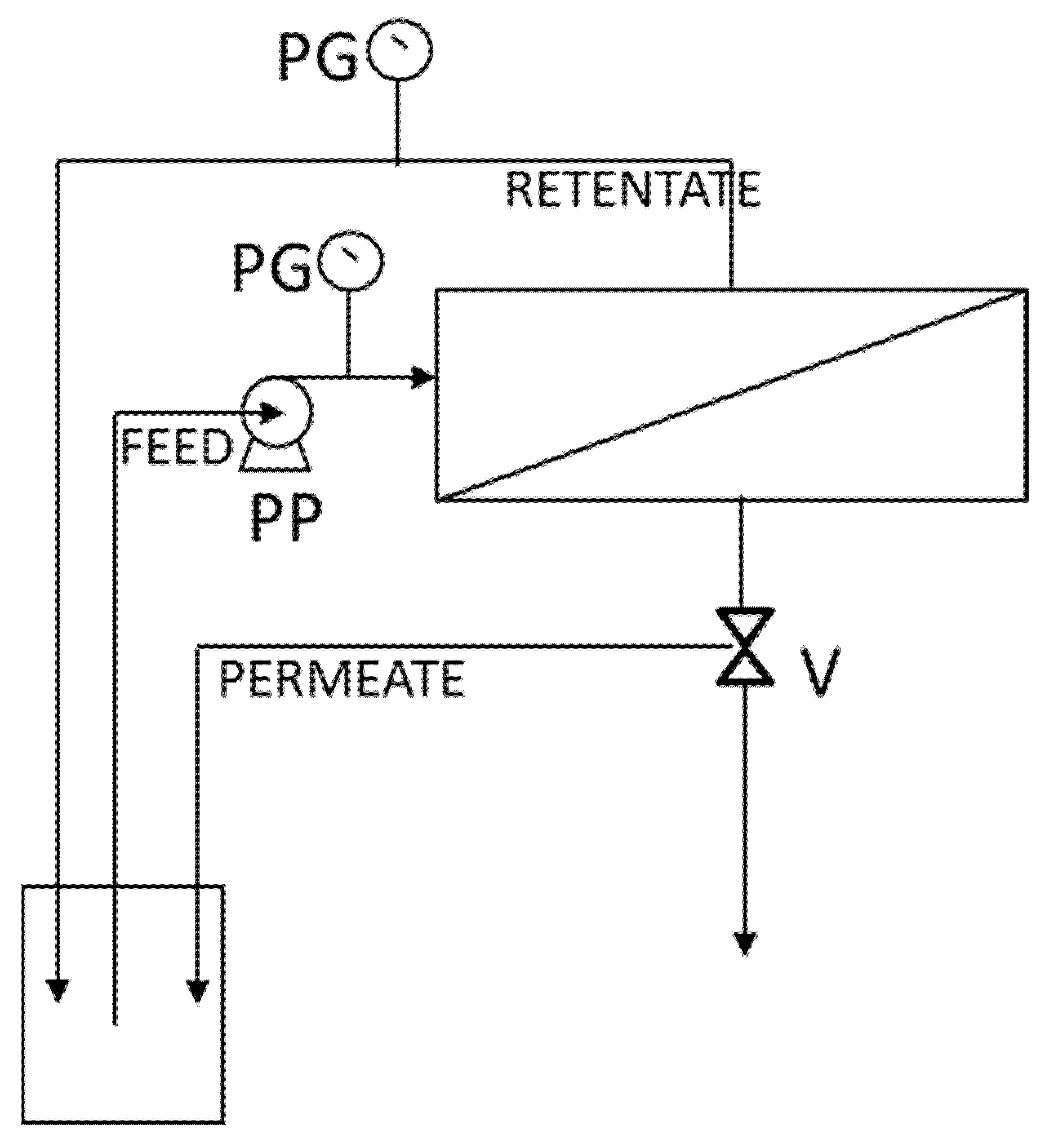
2.4. Experimental Setup
2.5. Ultrafiltration Experiments
2.6. Imparting Positive Charge to Regenerated Cellulose Membranes
3. Results
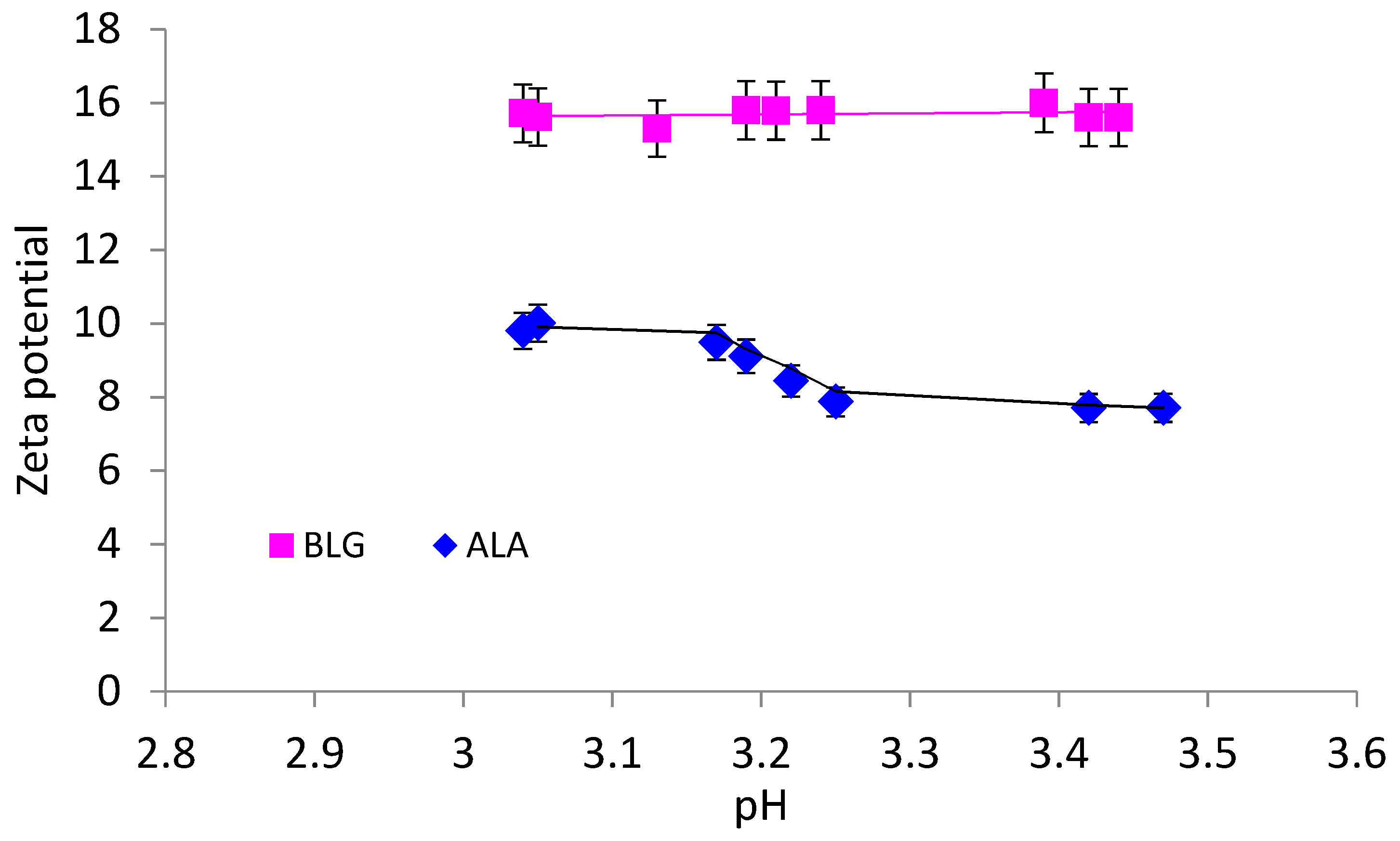
3.1. Properties of Individual Proteins in Bulk Solution
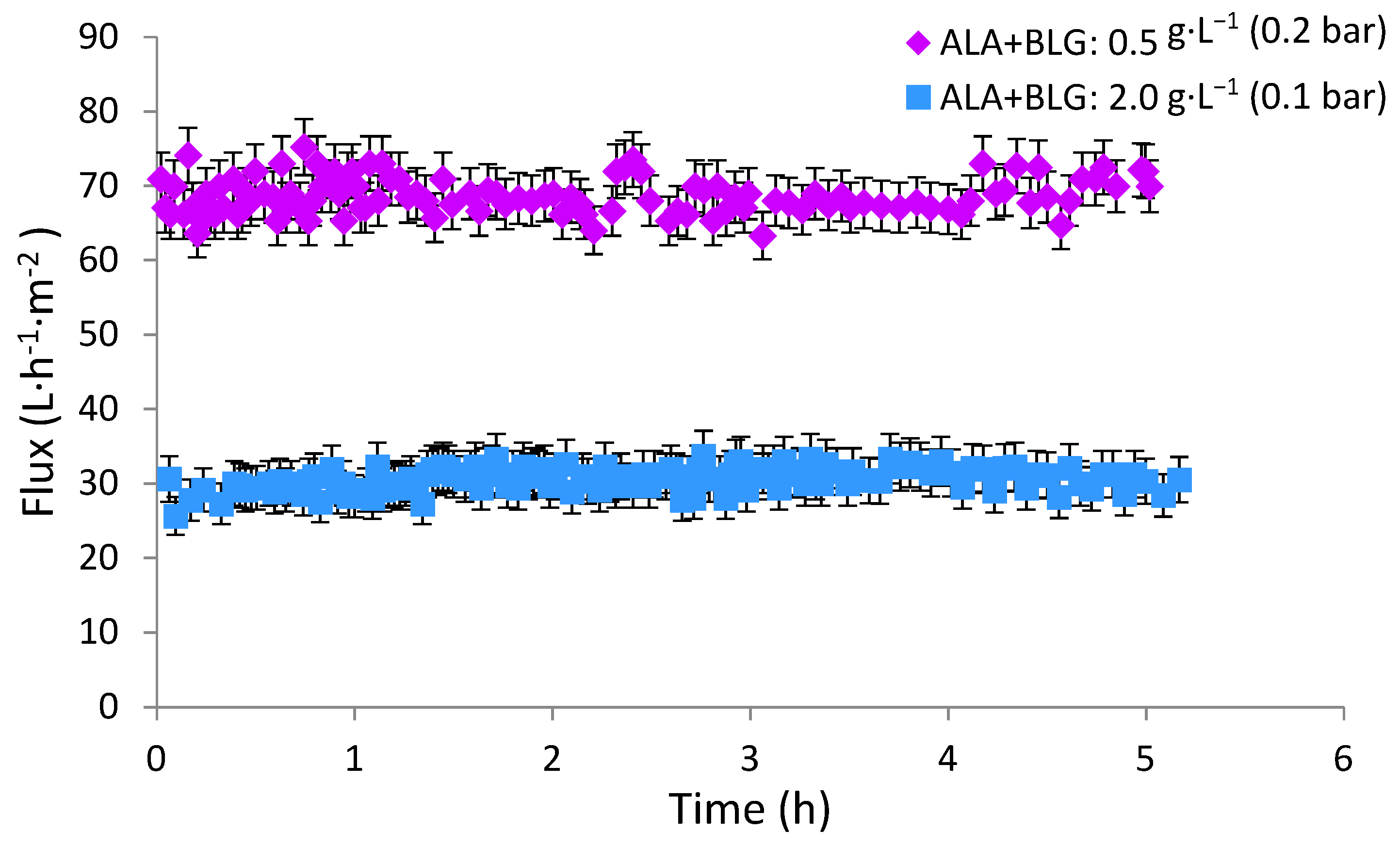
3.2. Determination of Critical Pressure
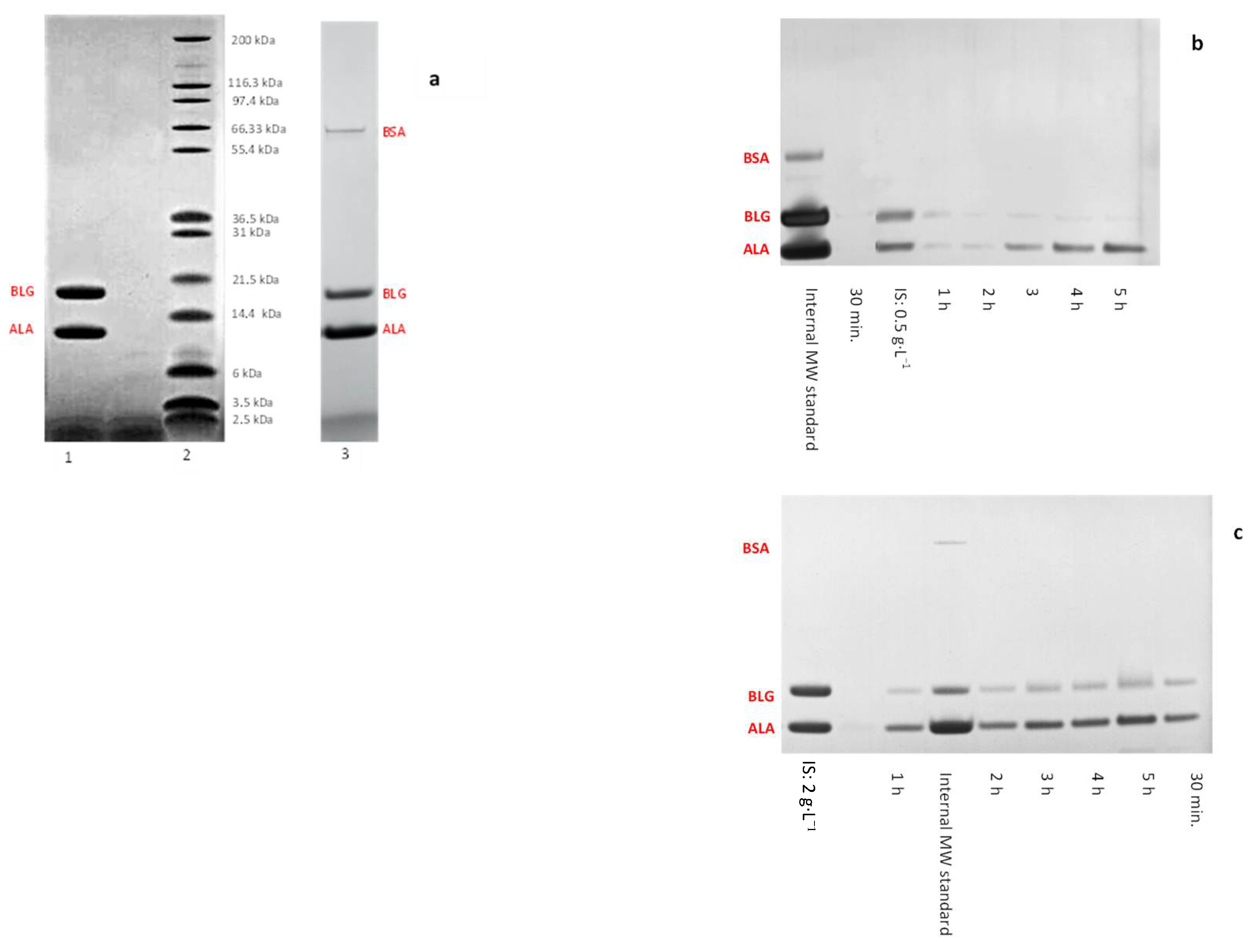
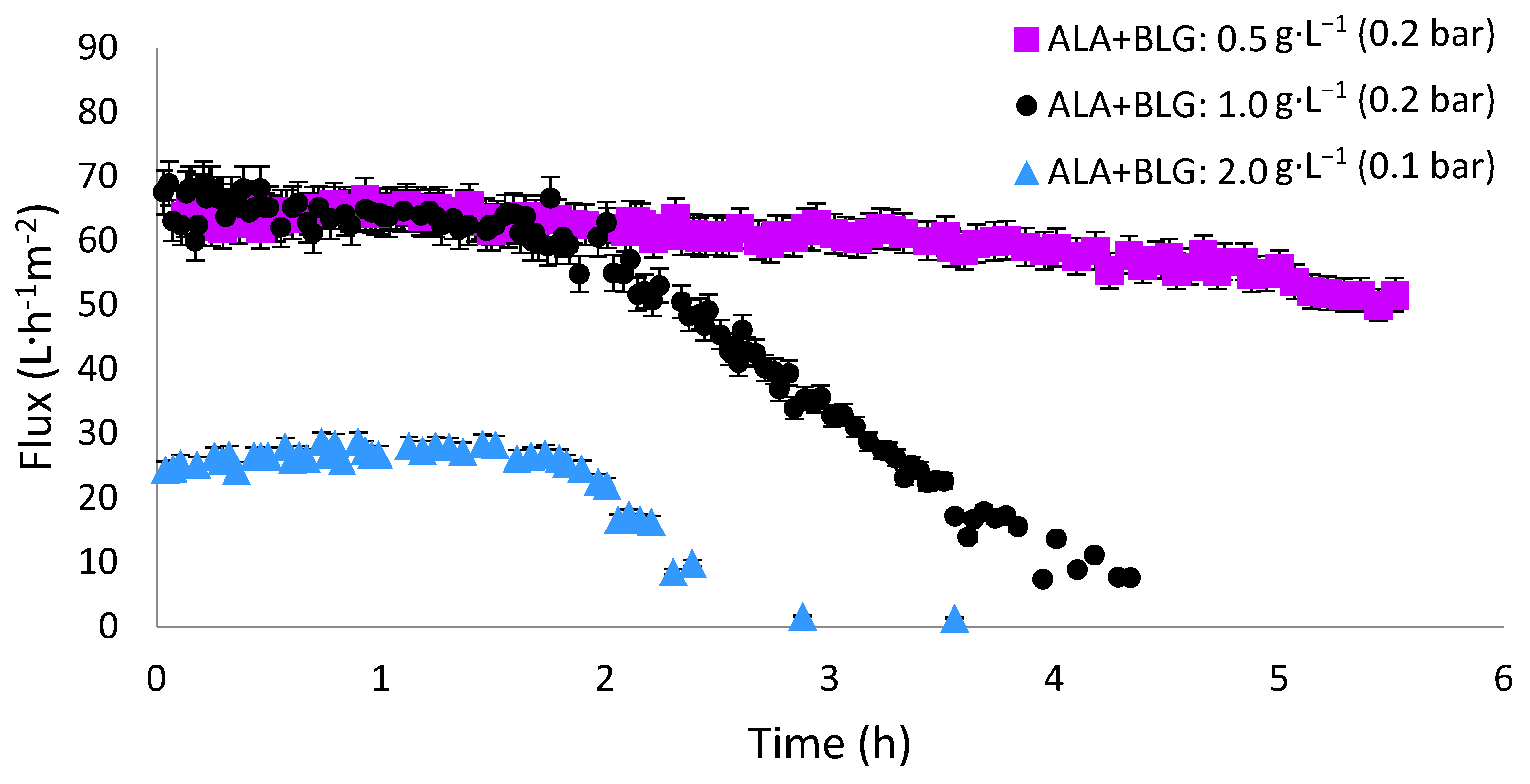
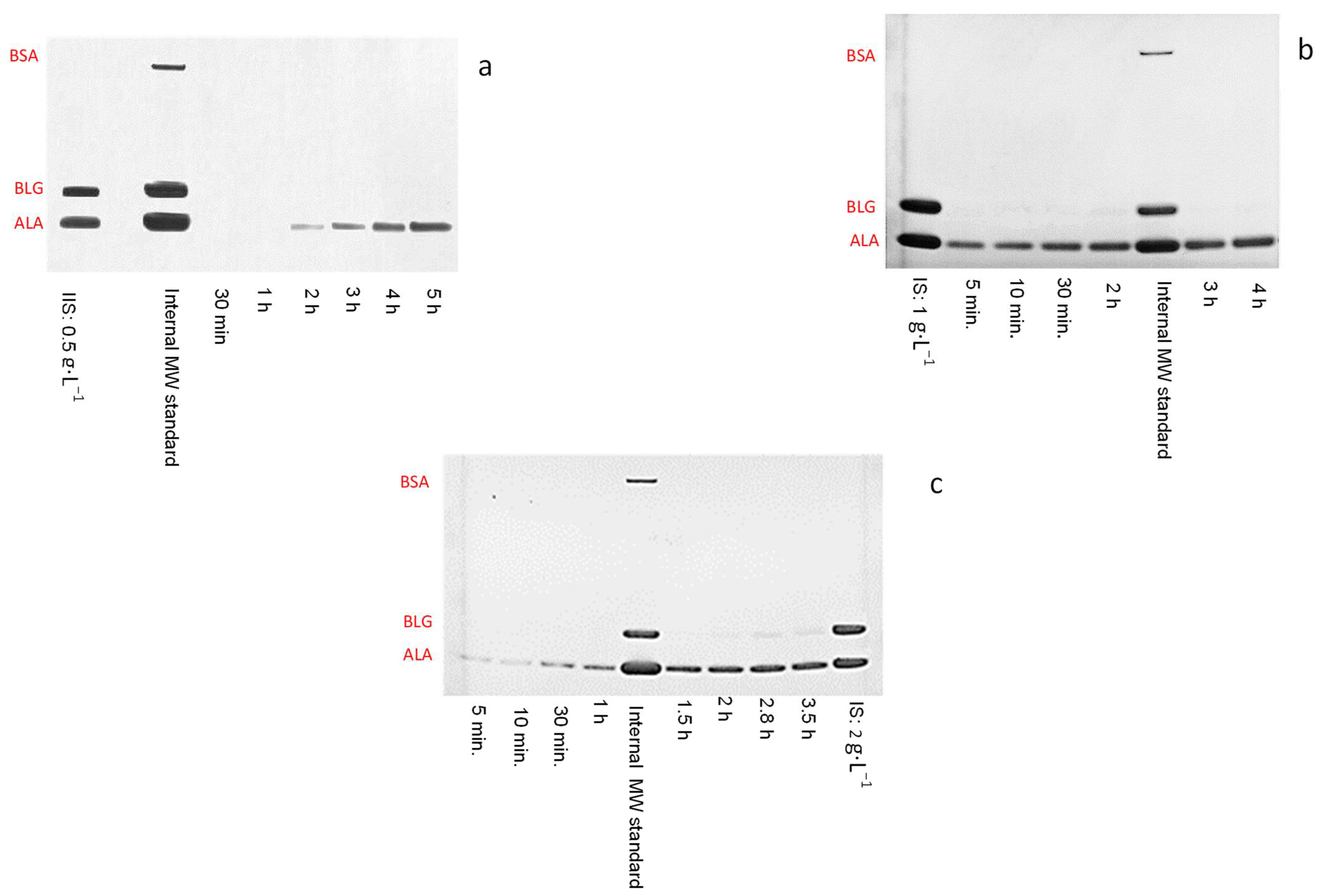
3.3. Binary Protein Mixture Ultrafiltration at pH 3 in Concentration Mode
3.4. Binary Protein Mixture Ultrafiltration at pH 3.4
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhushan, S.; Etzel, M.R. Charged ultrafiltration membranes increase the selectivity of whey protein separations. J. Food Sci. 2009, 74, E131–E139. [Google Scholar] [CrossRef]
- Saxena, A.; Tripathi, B.P.; Kumar, M.; Shahi, V.K. Membrane-based techniques for the separation and purifi cation of proteins: An overview. Adv. Colloid Interface Sci. 2009, 145, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Rohani, M.M.; Mehta, A.; Zydney, A.L. Development of high performance charged ligands to control protein transport through charge-modified ultrafiltration membranes. J. Membr. Sci. 2010, 362, 434–443. [Google Scholar] [CrossRef]
- Arunkumar, A.; Etzel, M.R. Fractionation of α-lactalbumin from β-lactoglobulin using positively charged tangential flow ultrafiltration membranes. Sep. Purif. Technol. 2013, 105, 121–128. [Google Scholar] [CrossRef]
- Cheang, B.; Zydney, A.L. Separation of α-lactalbumin and β-lactoglobulin using membrane ultrafiltration. Biotechnol. Bioeng. 2003, 83, 201–209. [Google Scholar] [CrossRef]
- Toro-Sierra, J.; Schumann, J.; Kulozik, U. Impact of spray-drying conditions on the particle size of microparticulated whey protein fractions. Dairy Sci. Technol. 2013, 93, 487–503. [Google Scholar] [CrossRef] [Green Version]
- Ratnaningsih, E.; Reynard, R.; Khoiruddin, K.; Wenten, I.G.; Boopathy, R. Recent Advancements of UF-Based Separation for Selective Enrichment of Proteins and Bioactive Peptides—A Review. Appl. Sci. 2021, 11, 1078. [Google Scholar] [CrossRef]
- van Reis, R.; Brake, J.; Charkoudian, J.; Burns, D.; Zydney, A. High-performance tangential flow filtration using charged membranes. J. Membr. Sci. 1999, 159, 133–142. [Google Scholar] [CrossRef]
- Rabiller-Baudry, M.; Chaufer, B.; Lucas, D.; Michel, F. Ultrafiltration of mixed protein solutions of lysozyme and lactoferrin: Role of modified inorganic membranes and ionic strength on the selectivity. J. Membr. Sci. 2001, 184, 137–148. [Google Scholar] [CrossRef]
- Chun, K.-Y.; Stroeve, P. Protein transport in nanoporous membranes modified with self-assembled monolayers of functionalized thiols. Langmuir 2002, 18, 4653–4658. [Google Scholar] [CrossRef]
- Ebersold, M.F.; Zydney, A.L. Separation of protein charge variants by ultrafiltration. Biotechnol. Prog. 2008, 20, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Striemer, C.C.; Gaborski, T.R.; McGrath, J.L.; Fauchet, P.M. Charge- and size-based separation of macromolecules using ultrathin silicon membranes. Nature 2006, 445, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Mahlicli, F.Y.; Altinkaya, S.A.; Yurekli, Y. Preparation and cha racterization of polyacrylonitrile membranes modified with polyelec trolytedepositionfor separatin g simil ar sized proteins. J. Membr. Sci. 2012, 415–416, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Sorci, M.; Gu, M.; Heldt, C.L.; Grafeld, E.; Belfort, G. A multi-dimensional approach for fractionating proteins using charged membranes. Biotechnol. Bioeng. 2013, 110, 1704–1713. [Google Scholar] [CrossRef] [PubMed]
- Valiño, V.; San Román, M.F.; Ibañez, R.; Ortiz, I. Improved separation of bovine serum albumin and lactoferrin mixtures using charged ultrafiltration membranes. Sep. Purif. Technol. 2014, 125, 163–169. [Google Scholar] [CrossRef]
- Fane, A.G.; Fell, C.J.D.; Suki, A. The effect of the pH and ionic environment on the ultrafiltration of protein solutions with retentive membranes. J. Membr. Sci. 1983, 16, 195–210. [Google Scholar] [CrossRef]
- Fane, A.G.; Fell, C.J.D.; Waters, A.G. Ultrafiltration of protein solutions through partially permeable membranes—The effect of adsorption and solution environment. J. Membr. Sci. 1983, 16, 211–224. [Google Scholar] [CrossRef]
- Kelly, S.T.; Zydney, A.L. Mechanisms for BSA fouling during microfiltration. J. Membr. Sci. 1995, 107, 115–124. [Google Scholar] [CrossRef]
- Ricq, L.; Narçon, S.; Reggiani, J.-C.; Pagetti, J. Streaming potential and protein transmission ultrafiltration of single proteins and proteins in mixture: B-lactoglobulin and lysozyme. J. Membr. Sci. 1999, 156, 81–96. [Google Scholar] [CrossRef]
- Martín, A.; Martinez, F.; Calvo, J.I.; Prádanos, P.; Palacio, L.; Hernández, A. Protein adsorption onto na inorganic microfiltration membrane solute-solid interactions and surface coverage. J. Membr. Sci. 2002, 207, 199–207. [Google Scholar] [CrossRef]
- Kujundzic, E.; Greenberg, A.R.; Fong, R.; Hernandez, M. Monitoring Protein Fouling on Polymeric Membranes Using Ultrasonic Frequency-Domain Reflectometry. Membranes 2011, 1, 195–216. [Google Scholar] [CrossRef] [Green Version]
- Portugal, C.A.M. Functional membranes for control of protein-surface interactions and transport properties. Curr. Org. Chem. 2017, 21, 1725–1739. [Google Scholar] [CrossRef]
- Aguero, R.; Bringas, E.; Román, M.S.; Ortiz, I.; Ibañez, R. Membrane processes for whey proteins separation and purification. A review. Curr. Org. Chem. 2017, 21, 1740–1752. [Google Scholar] [CrossRef]
- Bramaud, C.; Aimar, P.; Daufin, G. Whey protein fractionation: Isoelectric precipitation of a-lactalbumin under gentle heat treatment. Biotechnol. Bioeng. 1997, 56, 391–397. [Google Scholar] [CrossRef]
- Salgin, S.; Salgin, U.; Bahadir, S. Zeta potentials and isoelectric points of biomolecules: The effect of ion types and ionic strengths. Int. J. Electrochem. Sci. 2012, 7, 12404–12414. [Google Scholar]
- Yan, Y.; Seeman, D.; Zheng, B.; Kizilay, E.; Xu, Y.; Dubin, P.L. pH-dependent aggregation and disaggregation of native β-lactoglobulin in low salt. Langmuir 2013, 29, 4584–4593. [Google Scholar] [CrossRef]
- Arunkumar, A.; Etzel, M.R. Fractionation of α-lactalbumin and β-lactoglobulin from bovine milk serum using staged, positively charged, tangential flow ultrafiltration membranes. J. Membr. Sci. 2014, 454, 488–495. [Google Scholar] [CrossRef]
- Chayen, N.; Dieckmann, M.; Dierks, K.; Fromme, P. Size and shape determination of proteins in solution by a noninvasive depolarized dynamic light scattering instrument. Ann. N. Y. Acad. Sci. 2004, 1027, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Mazzei, R.; Drioli, E.; Giorno, L. Biocatalytic membrane reactor and membrane emulsification concepts combined in a single unit to assist production and separation of water unstable reaction products. J. Membr. Sci. 2010, 352, 166–172. [Google Scholar] [CrossRef]
- Mazzei, R.; Drioli, E.; Giorno, L. Comprehensive Membrane Science and Engineering. Biocatal. Membr. Membr. Bioreact. Compr. Membr. Sci. Eng. 2010, 3, 195–212. [Google Scholar]
- van Reis, R.D. Charged Filtration Membranes and Uses Therefor. U.S. Patent 7,001,550B2, 26 December 2006. Available online: https://app.dimensions.ai/details/patent/US-7153426-B2 (accessed on 21 September 2021).
- Filipe, C.D.; Ghosh, R. Effects of protein–protein interaction in ultrafiltration based fractionation processes. Biotechnol. Bioeng. 2005, 91, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Holland, B.; Kackmar, J.; Corredig, M. Short communication: Isolation of a whey fraction rich in α-lactalbumin from skim milk using tangential flow ultrafiltration. J. Dairy Sci. 2012, 95, 5604–5607. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Protein Diameter (nm) | Molecular Weight (kDa) | Pd (%) * | |
|---|---|---|---|---|
| ALA | 3.0 | 3.62 (±0.60) | 13.5 (±4.5) | 14.7 |
| 3.2 | 3.62 (±0.48) | 13.5 (±3.9) | 15.2 | |
| 3.4 | 3.62 (±0.36) | 13.5 (±5.3) | 16.0 | |
| BLG | 3.0 | 4.19 (±0.71) | 19.0 (±5.4) | 15.4 |
| 3.2 | 4.19 (±0.99) | 26.7 (±10.1) | 19.5 | |
| 3.4 | 4.89 (±1.36) | 26.7 (±6.5) | 27.1 |
| Protein Mixture (g·L−1) | pH | Critical Pressure (bar) | Critical Flux (L·h−1·m−2) | Rtot (m−1) | Rm (m−1) | Rfrev (m−1) | Rfirr (m−1) |
|---|---|---|---|---|---|---|---|
| 0.5 | 3.0 | 0.2 | 68 (±5) | 1.00 × 1012 (±3.00 × 1010) | 9.67 × 1011 (±5.80 × 1010) | 3.34 × 1010 (±2.34 × 109) | 0 |
| 3.4 | 0.2 | 64 (±5) | 1.24 × 1012 (±7.44 × 1010) | 1.01 × 1012 (±5.00 × 1010) | 1.64 × 1011 (±2.40 × 1010) | 2.20 × 1010 * | |
| 1 | 3 | 0.2 | 70 (±3) | 1.18 × 1012 (±9.44 × 1010) | 8.29 × 1011 (±5.80 × 1010) | 3.50 × 1011 (±2.45 × 1010) | 0 |
| 3.4 | 0.2 | 68 (±3) | 1.68 × 1012 (±1.01 × 1010) | 8.7 × 1011 (±5.22 × 1010) | 8.10 × 1011 (±4.86 × 1010) | 0 | |
| 2 | 3.0 | 0.1 | 35 (±5) | 1.16 × 1012 (±6.96 × 1010) | 9.20 × 1011 (±4.60 × 1010) | 2.15 × 1011 (±2.60 × 1010) | 2.56 × 1010 * |
| 3.4 | 0.1 | 25 (±5) | 1.68 × 1012 (±8.40 × 1010) | 9.73 × 1011 (±5.84 × 1010) | 6.79 × 1011 (±4.75 × 1010) | 2.62 × 1010 * |
| Initial Binary Protein Mixture Concentration (mg/mL) | ALA Recovery Factor (%) | ALA Purity (%) |
|---|---|---|
| 0.5 | 33 | 95 |
| 1.0 | 32 | |
| 2.0 | 33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzei, R.; Szymczak, A.M.; Drioli, E.; Al-Fageeh, M.; Aljohi, M.A.; Giorno, L. High Purity of α-Lactalbumin from Binary Protein Mixture by Charged UF Membrane Far from the Isoelectric Point to Limit Fouling. Appl. Sci. 2021, 11, 9167. https://doi.org/10.3390/app11199167
Mazzei R, Szymczak AM, Drioli E, Al-Fageeh M, Aljohi MA, Giorno L. High Purity of α-Lactalbumin from Binary Protein Mixture by Charged UF Membrane Far from the Isoelectric Point to Limit Fouling. Applied Sciences. 2021; 11(19):9167. https://doi.org/10.3390/app11199167
Chicago/Turabian StyleMazzei, Rosalinda, Anna Maria Szymczak, Enrico Drioli, Mohamed Al-Fageeh, Mohammed A. Aljohi, and Lidietta Giorno. 2021. "High Purity of α-Lactalbumin from Binary Protein Mixture by Charged UF Membrane Far from the Isoelectric Point to Limit Fouling" Applied Sciences 11, no. 19: 9167. https://doi.org/10.3390/app11199167