
Water and Sediment Bacterial Communities in a Small Mediterranean, Oxygen-Stratified, Saline Lake (Lake Alboraj, SE Spain)

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Water and Sediment Samples for Chemistry and Bacterial Analysis
2.3. Physico-Chemical and Pigment Profiling
2.4. Chemical Analysis
2.5. DNA Extraction and MiSeq Illumina Sequencing
3. Results
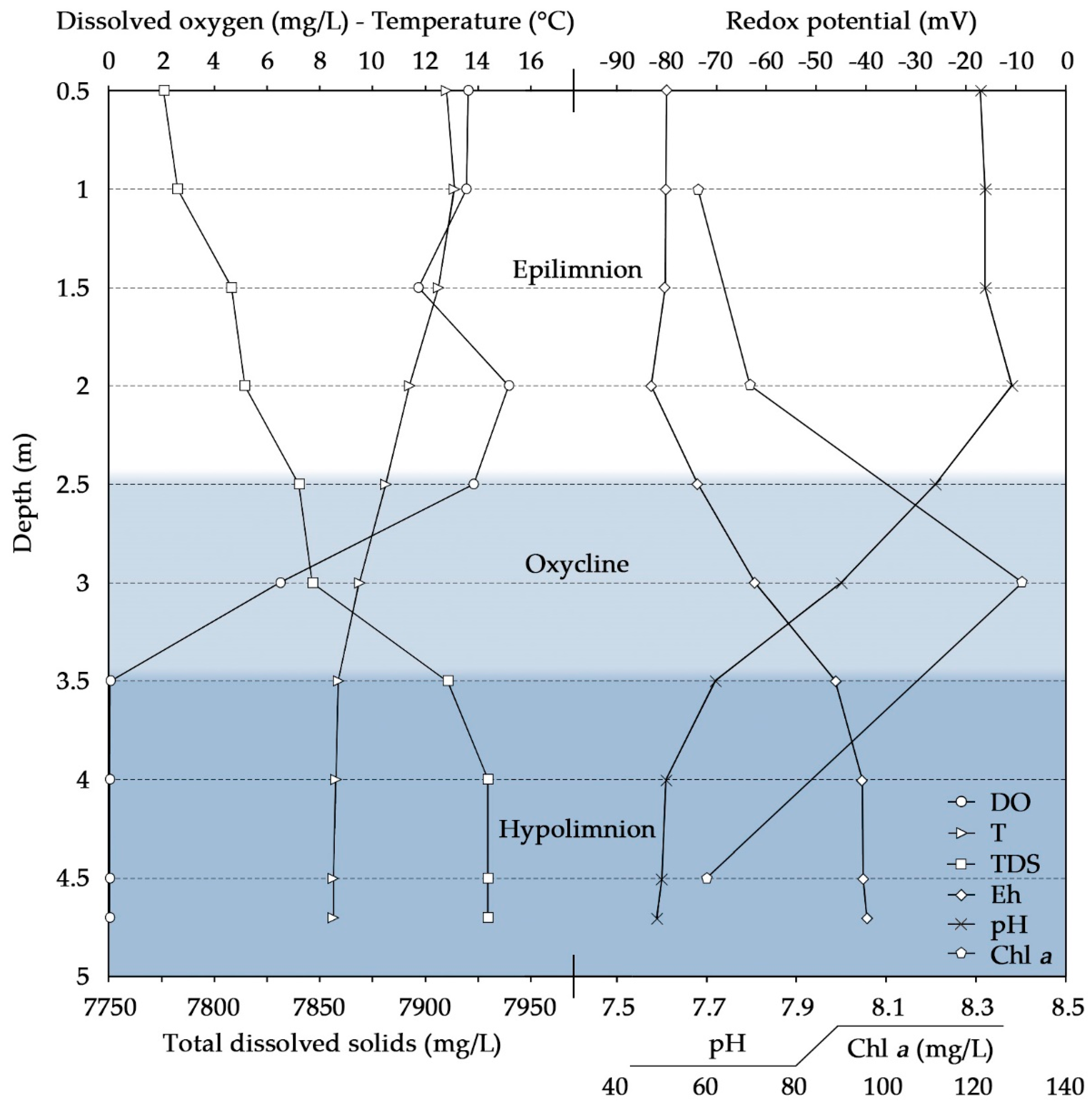
3.1. Chemical and Ecological Changes in the Water Column
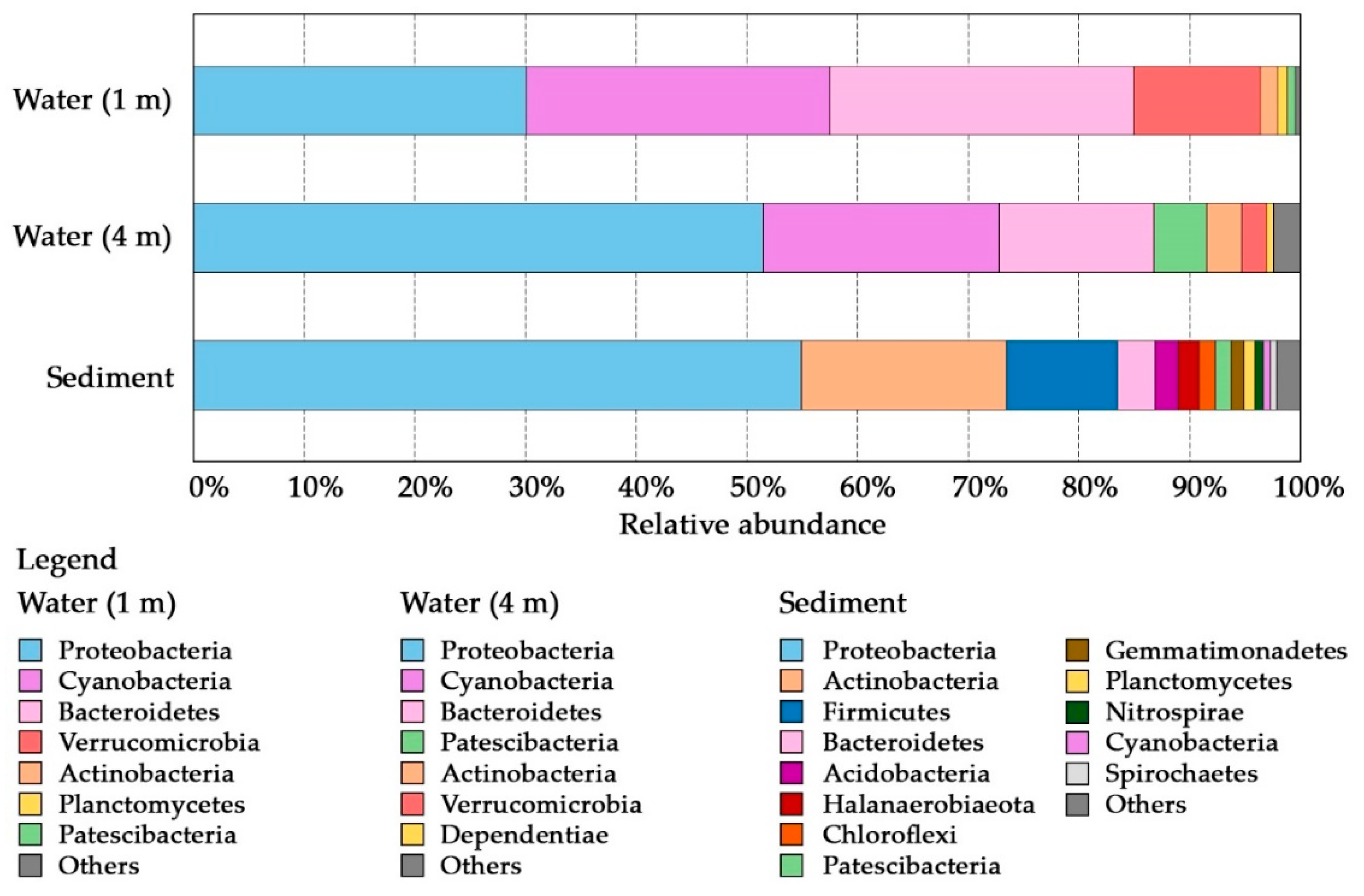
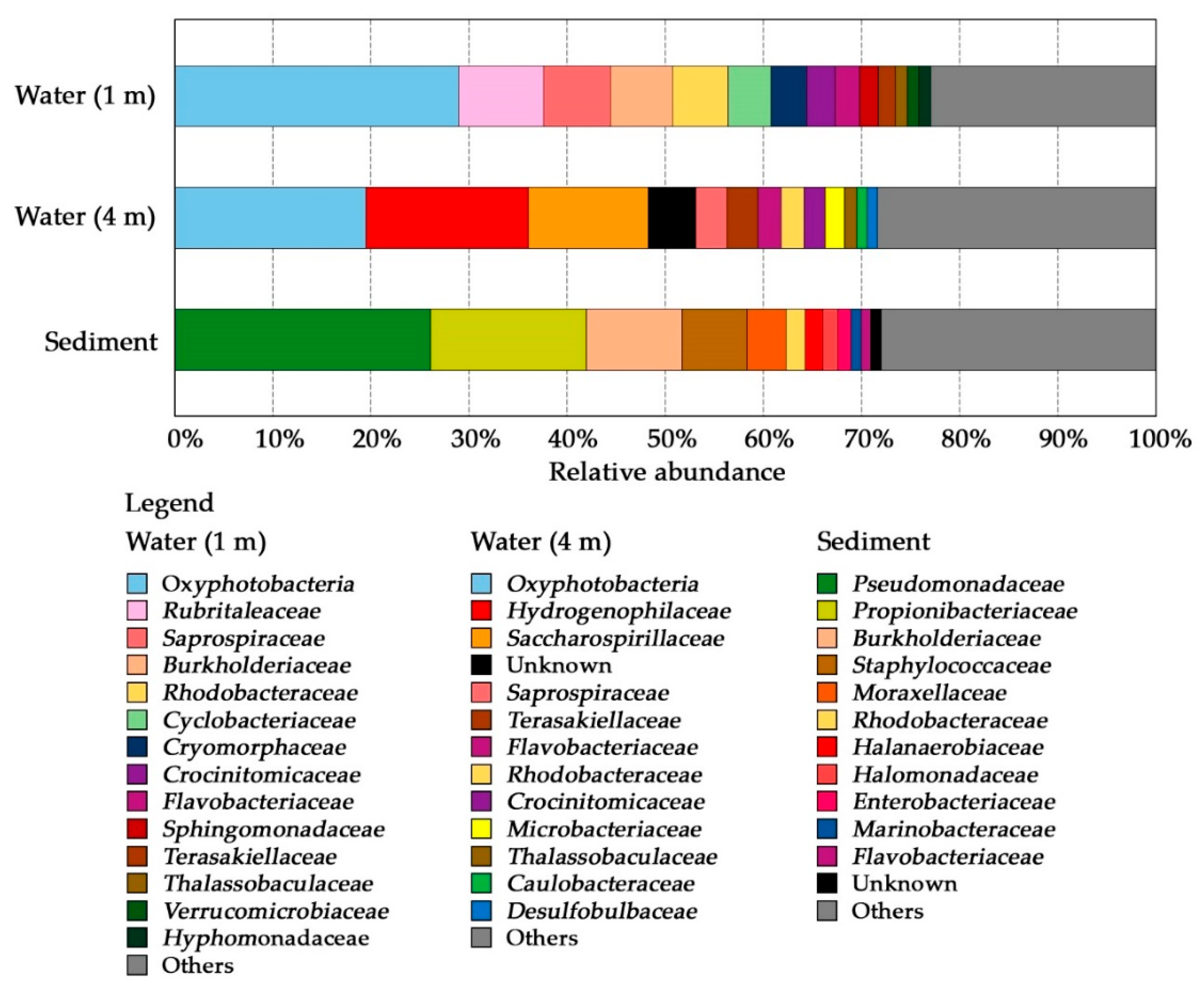
3.2. Comparative Analysis of the Microbial Structure
3.2.1. Water Associated Bacterial Communities
3.2.2. Sediment Associated Microbial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kelly-Quinn, M.; Biggs, J.; von Fumetti, S. Preface: The importance of small water bodies. Hydrobiologia 2017, 793, 1–2. [Google Scholar] [CrossRef]
- Bolpagni, R.; Poikane, S.; Laini, A.; Bagella, S.; Bartoli, M.; Cantonati, M. Ecological and Conservation Value of Small Standing-Water Ecosystems: A Systematic Review of Current Knowledge and Future Challenges. Water 2019, 11, 402. [Google Scholar] [CrossRef] [Green Version]
- Cebrián, A. El patrimonio natural de Albacete como recurso turístico. Gran Tour Rev. Investig. Turísticas 2010, 2, 62–82. [Google Scholar]
- Camacho, A.; Borja, C.; Valero-Garcés, B.; Sahuquillo, M.; Cirujano, S.; Soria, J.M.; Rico, E.; De La Hera, A.; Santamans, A.C.; García De Domingo, A.; et al. 3190 Lagos y lagunas kársticas sobre yesos. In Bases Ecológicas Preliminares para la Conservación de los Tipos de Hábitat de Interés Comunitario en España; Ministerio de Medio Ambiente, y Medio Rural y Marino: Madrid, Spain, 2009. [Google Scholar]
- Boronat, L.; Miracle, M.R.; Armengol, X. Cladoceran assemblages in a mineralization gradient. Hydrobiologia 2001, 442, 75–88. [Google Scholar] [CrossRef]
- Millán, A.; Moreno, J.L.; Velasco, J. Estudio faunístico y ecológico de los coleópteros y heterópteros acuáticos de las lagunas y humedales de Albacete (Alboraj, Los Patos, Ojos de Villaverde, Ontalafia y Pétrola). Sabuco 2001, 1, 43–94. [Google Scholar]
- Soria, J.; Vicente, E.; Camacho, A.; Miracle, M.; Rico, E.; García-Avilés, J.; Rueda, J.; Armengol, X.; Roca, J. Estudio Limnológico de 28 Humedales de Castilla-La Mancha Como Base para la Elaboración del Plan de Ordenación de Recursos Naturales; Junta de Comunidades de Castilla-La Mancha: Valencia, Spain, 1998. [Google Scholar] [CrossRef]
- Cirujano, S. Flora y Vegetación de las Lagunas y Humedales de la Provincia de Albacete; Instituto de Estudios Albacetenses: Albacete, Spain, 1990; p. 52. ISBN 978-84-8713-617-7. [Google Scholar]
- Moreno, J.L.; Menchén, A.; Toledo, B.; Sanz, D.; Gómez-Alday, J.J. Formación de oxiclina estacional, anoxia y dinámica de nutrientes en una pequeña laguna kárstica del SE ibérico (Laguna de Alboraj, Albacete). In Proceedings of the Congress of the Iberian Association of Limnology & III Iberoamerican Congress of Limnology, Murcia, Spain, 26–29 October 2020. [Google Scholar]
- Moreno, J.L.; Ortega, J.F.; Moreno, M.A.; Ballesteros, R. Using Unmanned aerial vehicles (UAV) for lake management: Ecological status, lake state shifts and biogeochemical processes in a small Mediterranean lake. In Proceedings of the Congress of the Iberian Association of Limnology & III Iberoamerican Congress of Limnology, Murcia, Spain, 26–29 October 2020. [Google Scholar]
- Zeglin, L.H. Stream microbial diversity in response to environmental changes: Review and synthesis of existing research. Front. Microbiol. 2015, 6, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, L.R.; Buxton, A.S.; Rees, H.C.; Bruce, K.; Brys, R.; Halfmaerten, D.; Read, D.S.; Watson, H.V.; Sayer, C.D.; Jones, E.P.; et al. Prospects and challenges of environmental DNA (eDNA) monitoring in freshwater ponds. Hydrobiologia 2019, 826, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Boehrer, B.; Schultze, M. Stratification of lakes. Rev. Geophys. 2008, 46. [Google Scholar] [CrossRef] [Green Version]
- Vegas, J.; Rodríguez-Pascua, M.; Perucha, M.A.; Pérez-López, R.; Israde-Alcántara, I.; Galán, L.; Mediato, J. Upper Holocene seismites in the Alboraj lake (I to IV centuries, Albacete, Spain). GeoTemas 2012, 13, 1543–1546. [Google Scholar]
- European Union. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Moreno, J.L.; Monteagudo, L.; Florin, M.; Chicote, A.; Ortega, J.F.; Ballesteros, R.; Moreno, M.A.; De Las Heras, J. Evolución histórica y reciente de la de laguna de Alboraj (Tobarra, Albacete): Bases para su estudio. In Proceedings of the III Jornadas Sobre el Medio Natural Albacetense, Albacete, Spain, 5–8 October 2016; Blanco, D., Fajardo, J., Ferrandis, P., Gómez, J., Picazo, J., Sanz, D., Valdés, A., Verde, A., Eds.; Instituto de Estudios Albacetenses: Albacete, Spain, 2018; pp. 73–81. ISBN 978-84-948930-0-1. [Google Scholar]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equation for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Plant Physiol. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Wood, E.D.; Armstrong, F.A.J.; Richards, F.A. Determination of nitrate in sea water by cadmium-copper reduction to nitrite. J. Mar. Biolog. Assoc. U. K. 1967, 47, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Koroleff, F. Direct determination of ammonia in natural waters as indophenol blue. Int. Con. Explor. Sea 1969, 9, 152–155. [Google Scholar]
- Petrosino, J.F.; Highlander, S.; Luna, R.A.; Gibbs, R.A.; Versalovic, J. Metagenomic Pyrosequencing and Microbial Identification. Clin. Chem. 2009, 55, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E. The Family Rubritaleaceae. In The Prokaryotes; Springer: Berlin, Germany, 2014; pp. 861–862. [Google Scholar] [CrossRef]
- Zárate, A.; Dorador, C.; Araya, R.; Guajardo, M.; Florez, J.Z.; Icaza, G.; Cornejo, D.; Valdés, J. Connectivity of bacterial assemblages along the Loa River in the Atacama Desert, Chile. PeerJ 2020, 8, e9927. [Google Scholar] [CrossRef]
- Oh, S.; Choi, D. Microbial Community Enhances Biodegradation of Bisphenol A Through Selection of Sphingomonadaceae. Microb. Ecol. 2018, 77, 631–639. [Google Scholar] [CrossRef]
- Basta, T.; Keck, A.; Klein, J.; Stolz, A. Detection and Characterization of Conjugative Degradative Plasmids in Xenobiotic-Degrading Sphingomonas Strains. J. Bacteriol. 2004, 186, 3862–3872. [Google Scholar] [CrossRef] [Green Version]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Hydrogenophilaceae fam. nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; Bergey’s Manual Trust and John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar] [CrossRef]
- Weiler, B.A.; Verhoeven, J.T.P.; Dufour, S.C. Bacterial Communities in Tissues and Surficial Mucus of the Cold-Water Coral Paragorgia arborea. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Oh, Y.S.; Roh, D.H. Phenylobacterium muchangponense sp. nov., isolated from beach soil, and emended description of the genus Phenylobacterium. Int. J. Syst. Evol. Microbiol. 2011, 62, 977–983. [Google Scholar] [CrossRef]
- Pujalte, M.J.; Lucena, T.; Ruvira, M.A.; Arahal, D.R.; Macián, M.C. The Family Rhodobacteraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin, Germany, 2014; pp. 439–512. [Google Scholar] [CrossRef]
- Klepac-Ceraj, V.; Bahr, M.; Crump, B.C.; Teske, A.P.; Hobbie, J.E.; Polz, M.F. High overall diversity and dominance of microdiverse relationships in salt marsh sulphate-reducing bacteria. Environ. Microbiol. 2004, 6, 686–698. [Google Scholar] [CrossRef]
- Gaytán, I.; Sánchez-Reyes, A.; Burelo, M.; Vargas-Suárez, M.; Liachko, I.; Press, M.; Sullivan, S.; Cruz-Gómez, M.J.; Loza-Tavera, H. Degradación del poliuretano recalcitrante y aditivos xenobióticos por una comunidad microbiana de vertederos seleccionada y su potencial biodegradativo revelado por análisis metagenómico basado en ligadura de proximidad. Front. Microbiol. 2019, 10, 2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evtushenko, L.I.; Takeuchi, M. The Family Microbacteriaceae. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006. [Google Scholar] [CrossRef]
- Ye, J.; Singh, A.; Ward, O.P. Biodegradation of nitroaromatics and other nitrogen-containing xenobiotics. World J. Microbiol. Biotechnol. 2004, 20, 117–135. [Google Scholar] [CrossRef]
- Rieger, P.G.; Meier, H.M.; Gerle, M.; Vogt, U.; Groth, T.; Knackmuss, H.J. Xenobiotics in the environment: Present and future strategies to obviate the problem of biological persistence. J. Biotechnol. 2002, 94, 101–123. [Google Scholar] [CrossRef]
- Compant, S.; Nowak, J.; Coenye, T.; Clément, C.; Ait Barka, E. Diversity and occurrence of Burkholderia spp. in the natural environment. FEMS Microbiol. Rev. 2008, 32, 607–626. [Google Scholar] [CrossRef] [Green Version]
- Shchegolkova, N.M.; Krasnov, G.S.; Belova, A.A.; Dmitriev, A.A.; Kharitonov, S.L.; Klimina, K.M.; Melnikova, N.V.; Kudryavtseva, A.V. Microbial Community Structure of Activated Sludge in Treatment Plants with Different Wastewater Compositions. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Lin, H.; Zhang, X. Simultaneous Ammonia Nitrogen and Phosphorus Removal from Micro-Polluted Water by Biological Aerated Filters with Different Media. Water Air Soil Pollut. 2020, 231. [Google Scholar] [CrossRef]
- Stackebrandt, E. The Family Propionibacteriaceae: Genera other than Propionibacterium. Prokaryotes 2014, 725–741. [Google Scholar] [CrossRef]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [Green Version]
- Velásquez-Puentes, F.J.; Bacon, C.D. An introduction to phylogenetic community structure: A case study in Bolivian palms. Ecol. Boliv. 2016, 51, 126–140. [Google Scholar]
- Peterson, A.T. Conservatism of ecological niches in evolutionary time. Science 1999, 285, 1265–1267. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Foley, B.; Jones, I.D.; Maberly, S.C.; Rippey, B. Long-term changes in oxygen depletion in a small temperate lake: Effects of climate change and eutrophication. Freshw. Biol. 2012, 57, 278–289. [Google Scholar] [CrossRef]
- Meyer, R.L.; Risgaard-Petersen, N.; Allen, D.E. Correlation between anammox activity and microscale distribution of nitrite in a subtropical mangrove sediment. Appl. Environ. Microbiol. 2005, 71, 6142–6149. [Google Scholar] [CrossRef] [Green Version]
- Risgaard-Petersen, N.; Nielsen, L.P.; Rysgaard, S.; Dalsgaard, T.; Meyer, R.L. Application of the isotope pairing technique in sediments where anammox and denitrification coexist. Limnol. Oceanogr. Meth. 2003, 1, 63–73. [Google Scholar] [CrossRef]
- Su, X.; He, Q.; Mao, Y.; Chen, Y.; Hu, Z. Dissolved oxygen stratification changes nitrogen speciation and transformation in a stratified lake. Environ. Sci. Pollut. Res. 2019, 26, 2898–2907. [Google Scholar] [CrossRef]
- Kirf, M.K.; Hans, R.; Holtappels, M.; Fischer, J.P.; Schubert, C.J.; Wehrli, B. Redox gradients at the low oxygen boundary of lakes. Aquat. Sci. 2015, 77, 81–93. [Google Scholar] [CrossRef]
- Degermendzhy, A.G.; Zadereev, E.S.; Rogozin, D.Y.; Prokopkin, I.G.; Barkhatov, Y.V.; Tolomeev, A.P.; Khromechek, E.B.; Janse, J.H.; Mooij, W.M.; Gulati, R.D. Vertical stratification of physical, chemical and biological components in two saline lakes Shira and Shunet (South Siberia, Russia). Aquat. Ecol. 2010, 44, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Tiedje, J. Ecology of denitrification and dissimilatory nitrate reduction to ammonium. In Biology of Anaerobic Microorganisms; Zehnder, J.M., Ed.; Wiley: New York, NY, USA, 1988; pp. 179–244. [Google Scholar]
- Jetten, M.S.; Niftrik, L.V.; Strous, M.; Kartal, B.; Keltjens, J.T.; Op den Camp, H.J. Biochemistry and molecular biology of anammox bacteria. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.W.; Farnsley, S.E.; LeCleir, G.R.; Layton, A.C.; Satchwell, M.F.; DeBruyn, J.M.; Boyer, G.L.; Zhu, G.; Paerl, H.W. The relationships between nutrients, cyanobacterial toxins and the microbial community in Taihu (Lake Tai), China. Harmful Algae 2011, 10, 207–215. [Google Scholar] [CrossRef]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, B.B. Ecology of the sulphur cycle: Oxidative pathways in the sediment. In The Nitrogen and Sulphur Cycles; Cole, J.A., Ferguson, S.J., Eds.; Cambridge University Press: Cambridge, UK, 1988; pp. 31–63. [Google Scholar]
- Holmer, M.; Storkholm, P. Sulphate reduction and sulphur cycling in lake sediments: A review. Freshw. Biol. 2001, 46, 431–451. [Google Scholar] [CrossRef]
- Rodríguez-Pascua, M.A.; Bischoff, J.; Garduño-Monroy, V.H.; Pérez-López, R.; Giner-Robles, J.L.; Israde-Alcántara, I.; Calvo, J.P.; Williams, R.W. Estimation of the tectonic slip-rate from Quaternary lacustrine facies within the intraplate Albacete province (SE of Spain). Sediment. Geol. 2009, 222, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Tarnovetskii, I.Y.; Merkel, A.Y.; Kanapatskii, T.A.; Ivanova, E.A.; Gulin, M.B.; Toshchannkov, S.; Pimenov, N.V. Decoupling between sulfate reduction and the anaerobic oxidation of methane in the shallow methane seep of the Black sea. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Tourova, T.P.; Slobodova, N.V.; Bumazhkin, B.K.; Kolganova, T.V.; Muyzer, G.; Sorokin, D.Y. Analysis of community composition of sulfur-oxidizing bacteria in hypersaline and soda lakes using soxB as a functional molecular marker. FEMS Microbiol. Ecol. 2013, 84, 280–289. [Google Scholar] [CrossRef] [Green Version]
- AnilKumar, P.; Srinivas, T.N.R. The family Cyclobacteriaceae. In The Prokaryotes: Other Major Lineages of Bacteria and the Archaea; Springer: Berlin, Germany, 2014; pp. 551–575. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Epilimnion | Hypolimnion | ||
|---|---|---|---|
| Depth | 1 | 4 | 4.5 |
| NH4+ | 0.04 | 2.2 | 0.06 |
| NO3− | 10.3 | 0.6 | 4.7 |
| TIN | 2.4 | 1.1 | |
| TON | 1.3 | 0.5 | |
| Ntotal | 3.7 | 3.20 | 1.6 |
| Ptotal | 0.1 | 0.12 | |
| DOC | 12.9 | 14.9 | |
| HCO3− | 379 | 398 | - |
| SO42− | 6310 | 5426 | - |
| Cl− | 1376 | 1187 | - |
| F− | 8.7 | 7.5 | - |
| Br− | 2.7 | 2.4 | - |
| Sample | Depth (m) | Habitat | Sequences Generated | OTUs Generated | Shannon | Phylogenetic Diversity |
|---|---|---|---|---|---|---|
| W1 | 1 | Water, epilimnion | 133,167 | 558 | 6.24 | 51.89 |
| W4 | 4 | Water, hypolimnion | 115,563 | 507 | 5.75 | 51.00 |
| S | 4.7 | Sediment | 130,596 | 308 | 6.12 | 43.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espín, Y.; Menchén, A.; Moreno, J.L.; Sanz, D.; Álvarez-Ortí, M.; Fernández, J.A.; Gómez-Alday, J.J. Water and Sediment Bacterial Communities in a Small Mediterranean, Oxygen-Stratified, Saline Lake (Lake Alboraj, SE Spain). Appl. Sci. 2021, 11, 6309. https://doi.org/10.3390/app11146309
Espín Y, Menchén A, Moreno JL, Sanz D, Álvarez-Ortí M, Fernández JA, Gómez-Alday JJ. Water and Sediment Bacterial Communities in a Small Mediterranean, Oxygen-Stratified, Saline Lake (Lake Alboraj, SE Spain). Applied Sciences. 2021; 11(14):6309. https://doi.org/10.3390/app11146309
Chicago/Turabian StyleEspín, Yolanda, Alfonso Menchén, José Luis Moreno, David Sanz, Manuel Álvarez-Ortí, José Antonio Fernández, and Juan José Gómez-Alday. 2021. "Water and Sediment Bacterial Communities in a Small Mediterranean, Oxygen-Stratified, Saline Lake (Lake Alboraj, SE Spain)" Applied Sciences 11, no. 14: 6309. https://doi.org/10.3390/app11146309