Identification of Genes Associated with Sensitivity to Ultraviolet A (UVA) Irradiation by Transposon Mutagenesis of Vibrio parahaemolyticus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Cell Preparation
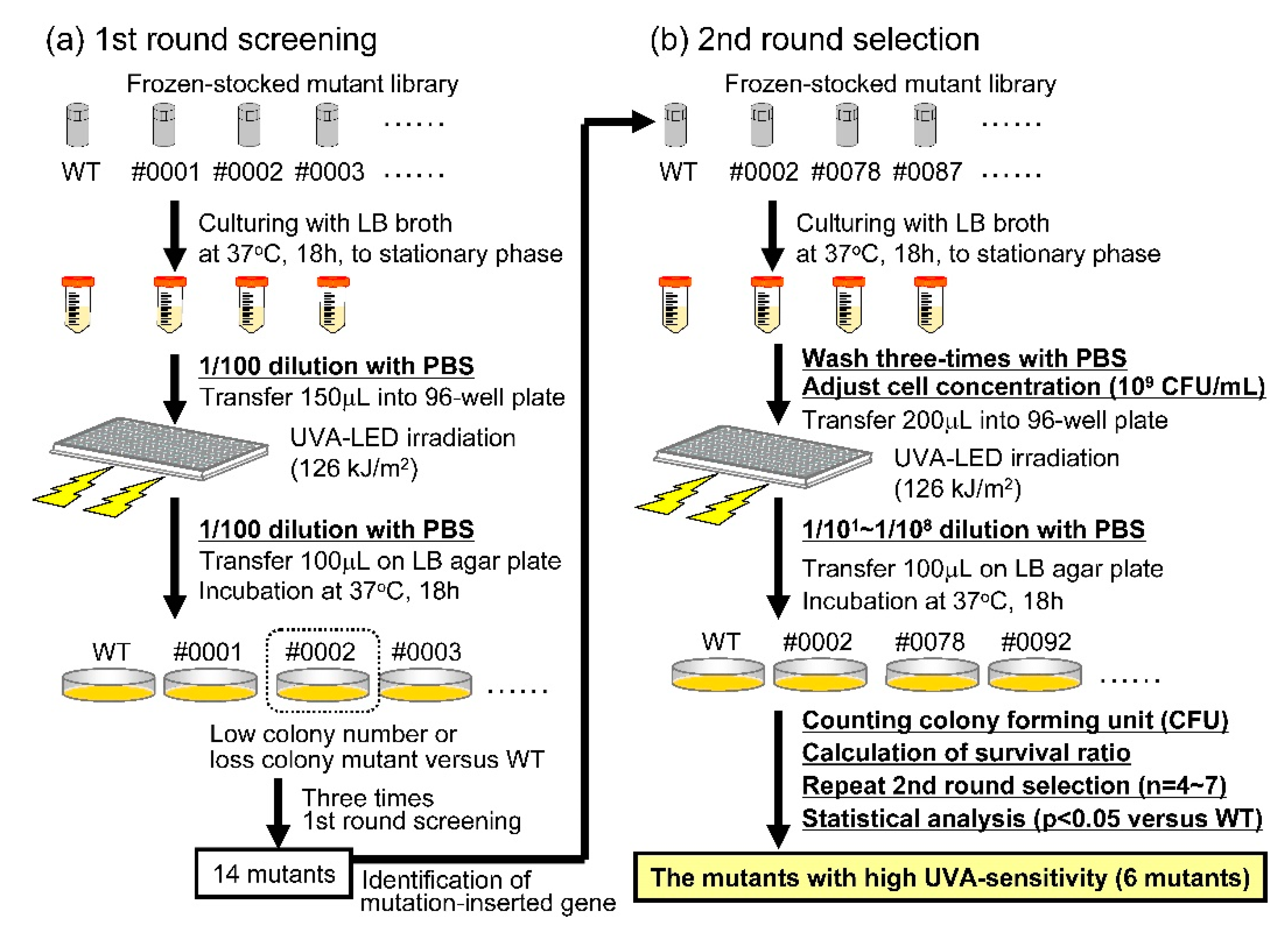
2.2. Construction of the V. parahaemolyticus Mutant Library and Screening of the Mutants for High Sensitivity to UVA
2.3. Gene Complementation in the Mutants
2.4. UV Irradiation
2.5. Measurement of Sensitivities to UV Irradiation
2.6. Intracellular ATP Concentration
2.7. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Silver Staining
2.8. Ubiquinone-8 Measurements
2.9. Statistical Analysis
3. Results
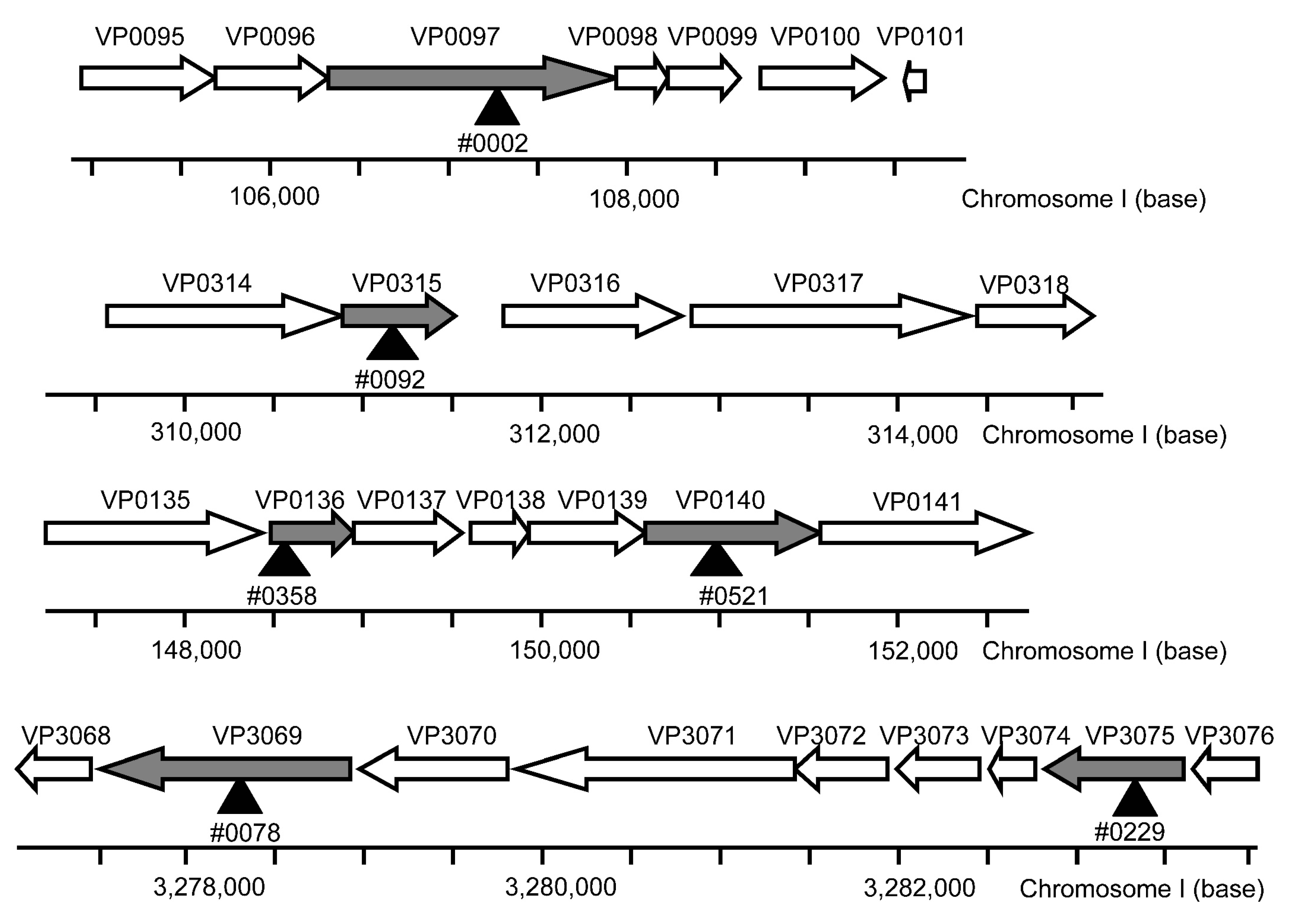
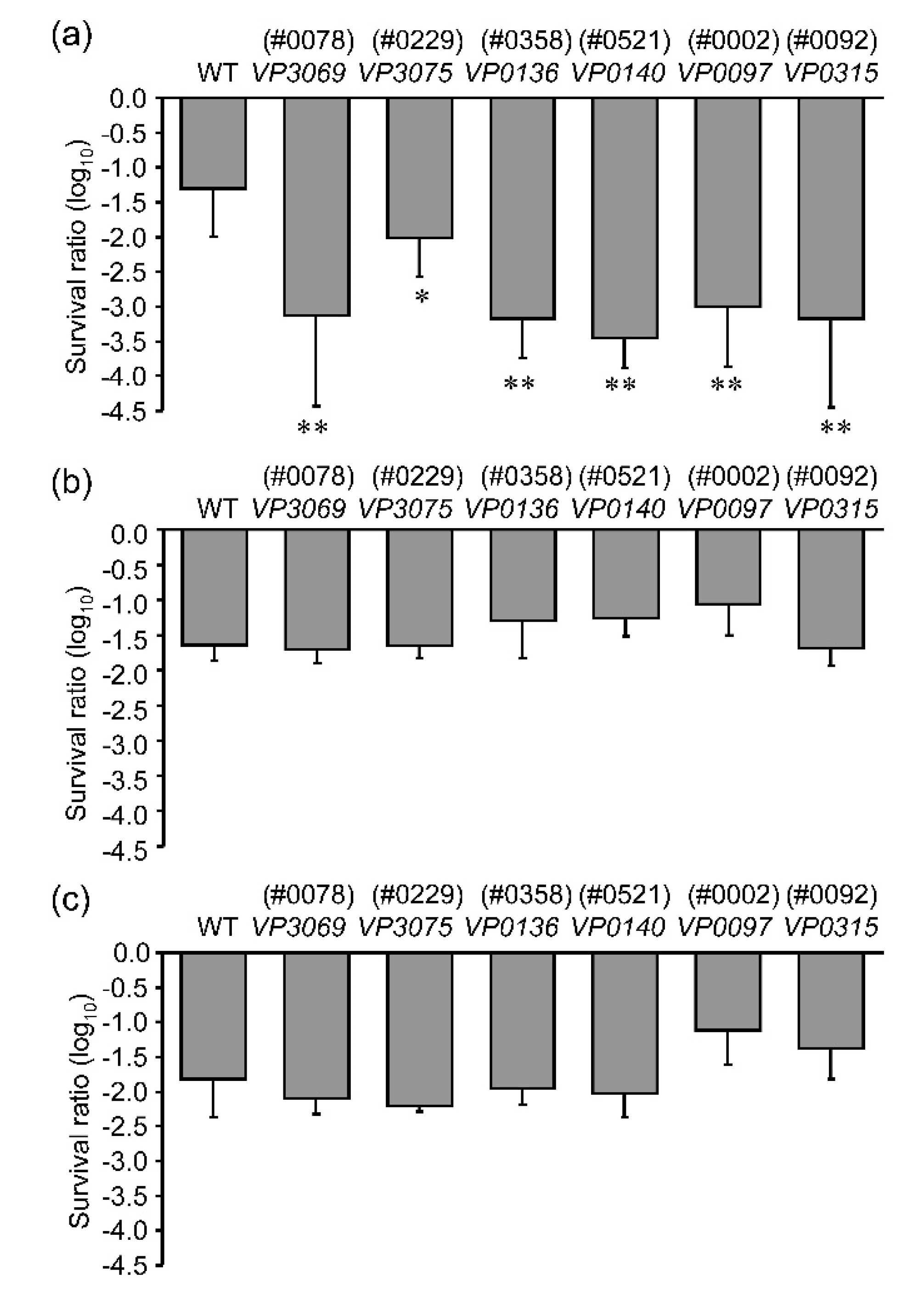
3.1. Screening of the Mutants with High UVA Sensitivity
3.2. Sensitivities to Different UV Wavelengths
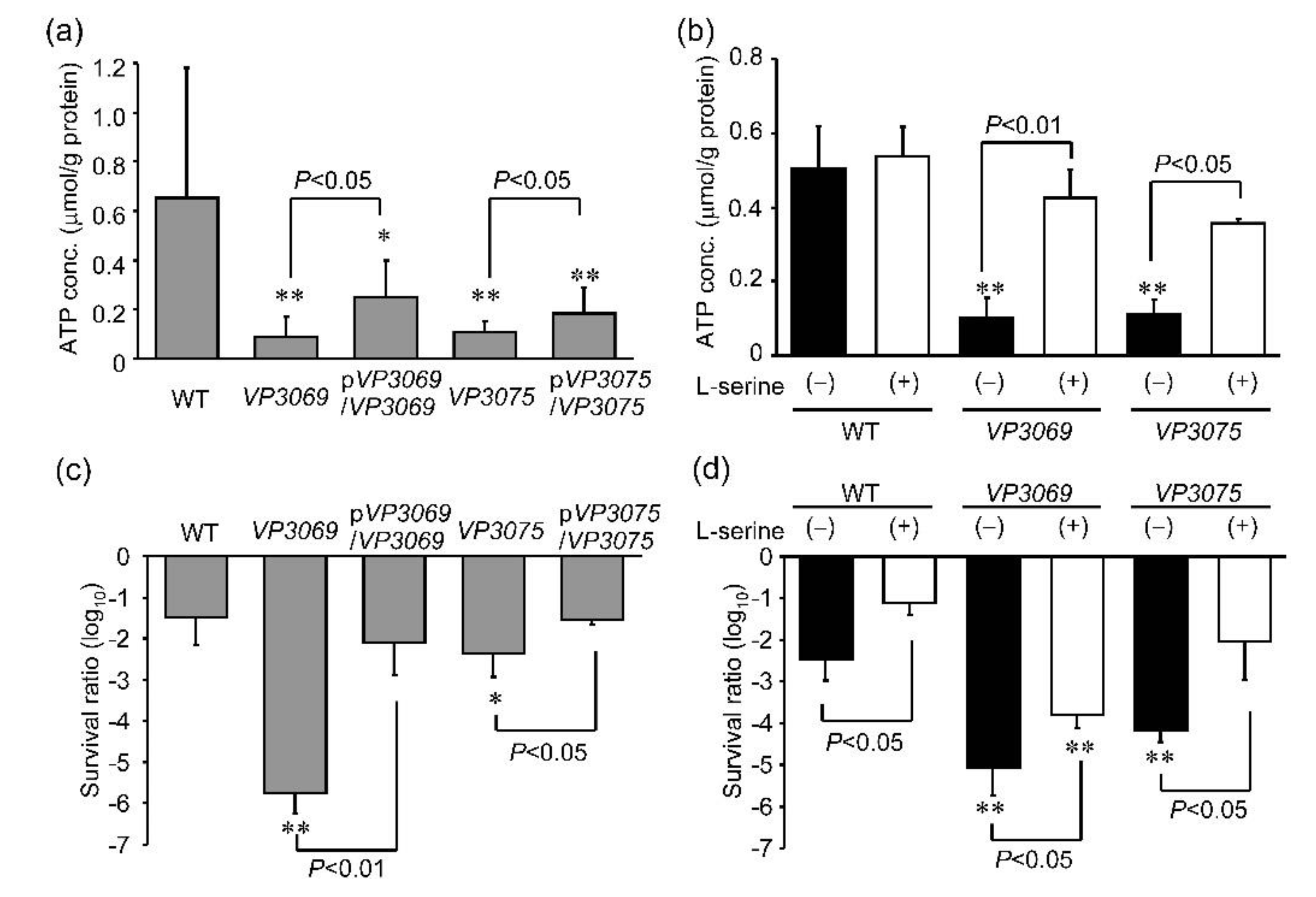
3.3. Evaluation of Intracellular ATP Concentration and Sensitivity to UVA Irradiation in Strains with Mutations in F0F1-Type ATP Synthase Genes
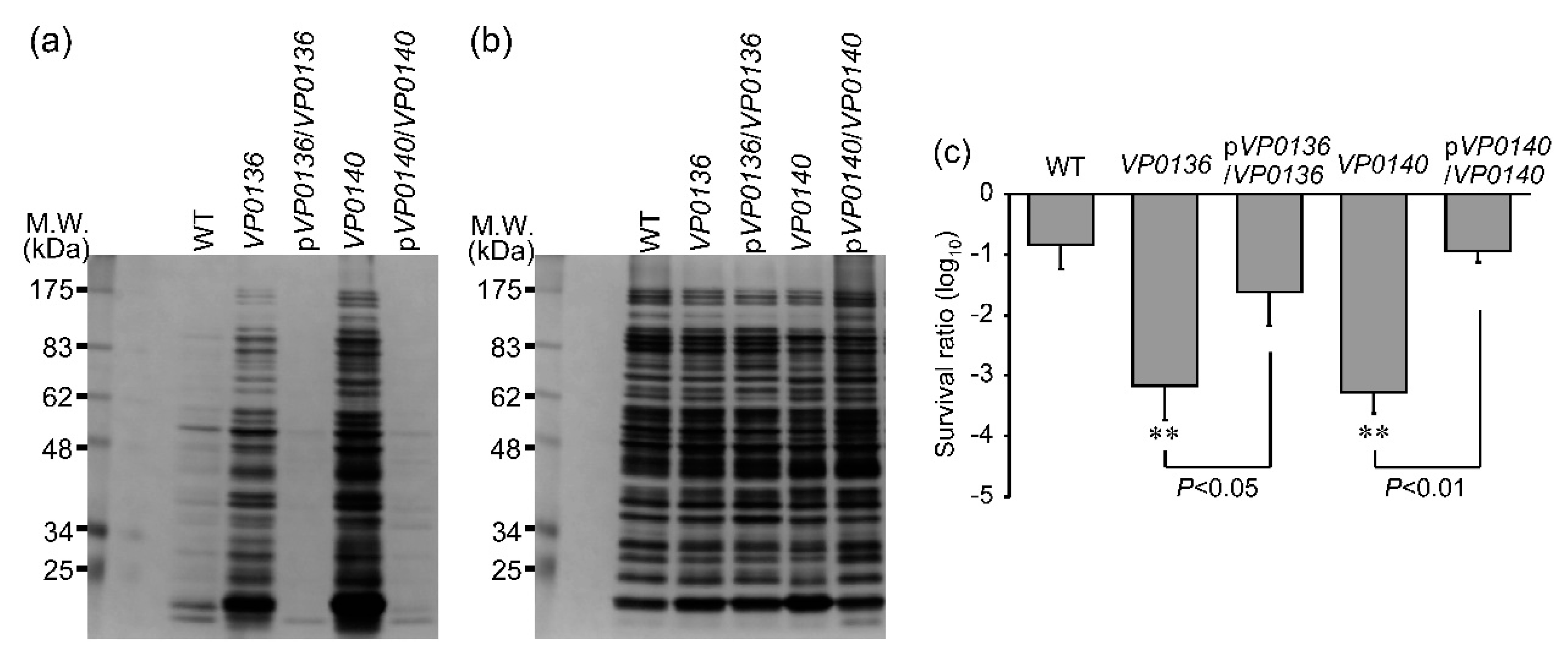
3.4. Evaluation of Extracellular Protein Content and Sensitivity to UVA among Mutants of General Secretion Pathway Genes
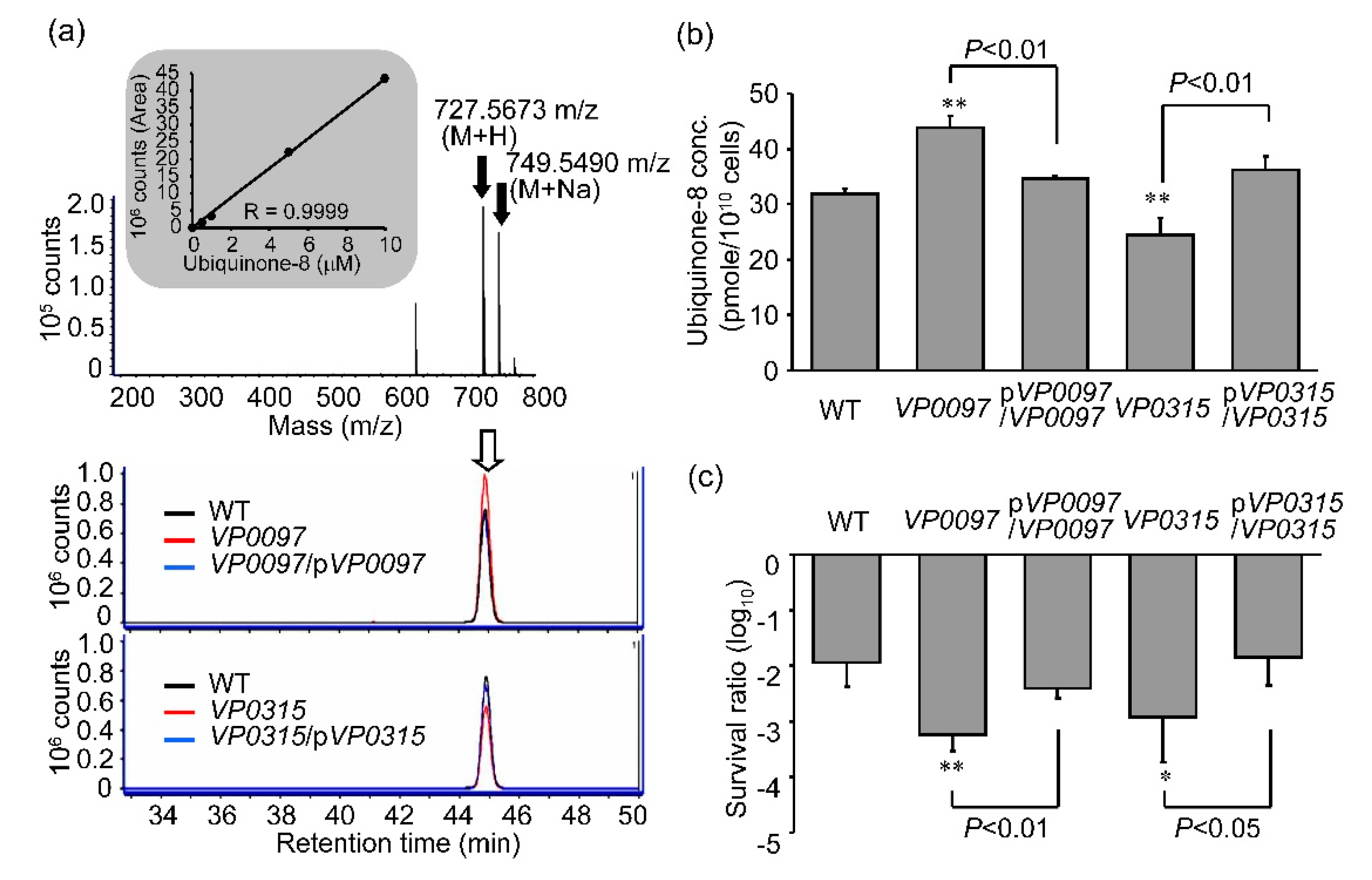
3.5. Intracellular Concentrations of Ubiquinone-8 and Sensitivity to UVA Irradiation among Strains with Mutations in Ubiquinone Biosynthesis Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DePaola, A.; Kaysner, C.A.; Bowers, J.; Cook, D.W. Environmental investigations of Vibrio parahaemolyticus in oysters after outbreaks in Washington, Texas, and New York (1997 and 1998). Appl. Environ. Microbiol. 2000, 66, 4649–4654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, J.B.; DePaola, A.; Bopp, C.A.; Martinek, K.A.; Napolilli, N.P.; Allison, C.G.; Murray, S.L.; Thompson, E.C.; Bird, M.M.; Middaugh, J.P. Outbreak of Vibrio parahaemolyticus gastroenteritis associated with Alaskan oysters. N. Engl. J. Med. 2005, 353, 1463–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, H.C.; Chen, M.C.; Liu, S.H.; Liu, D.P. Incidence of highly genetically diversified Vibrio parahaemolyticus in seafood imported from Asian countries. Int. J. Food Microbiol. 1999, 52, 181–188. [Google Scholar] [CrossRef]
- Levine, W.C.; Griffin, P.M. Vibrio infections on the Gulf Coast: Results of first year of regional surveillance. Gulf Coast Vibrio Working Group. J. Infect. Dis. 1993, 167, 479–483. [Google Scholar] [CrossRef]
- Nishibuchi, M.; Kaper, J.B. Thermostable direct hemolysin gene of Vibrio parahaemolyticus: A virulence gene acquired by a marine bacterium. Infect. Immun. 1995, 63, 2093–2099. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Ni, Y.X.; Miwatani, T. Purification and characterization of a hemolysin produced by a clinical isolate of Kanagawa phenomenon-negative Vibrio parahaemolyticus and related to the thermostable direct hemolysin. Infect. Immun. 1988, 56, 961–965. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Ono, T.; Rokuda, M.; Jang, M.H.; Okada, K.; Iida, T.; Honda, T. Functional characterization of two type III secretion systems of Vibrio parahaemolyticus. Infect. Immun. 2004, 72, 6659–6665. [Google Scholar] [CrossRef] [Green Version]
- Velazquez-Roman, J.; León-Sicairos, N.; de Jesus Hernández-Díaz, L.; Canizalez-Roman, A. Pandemic Vibrio parahaemolyticus O3:K6 on the American continent. Front. Cell Infect. Microbiol. 2014, 110, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Tack, D.M.; Ray, L.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Rissman, T.; Jervis, R.; Lathrop, S.; Muse, A.; Duwell, M.; et al. Preliminary incidence and trends of infections with pathogens transmitted commonly through food-foodborne diseases active surveillance network, 10 U.S. Sites, 2016–2019. MMWR Morb. Mortal Wkly. Rep. 2020, 69, 509–514. [Google Scholar] [CrossRef]
- Fawell, J.; Robinson, D.; Bull, R.; Birnbaum, L.; Butterworth, B.; Daniel, P.; Galal-Gorchev, H.; Hauchman, F.; Julkunen, P.; Klaassen, C.; et al. Disinfection byproducts in drinking water: Critical issues in health effects research. Environ. Health Perspect. 1997, 105, 108–109. [Google Scholar] [CrossRef]
- von Gunten, U. Ozonation of drinking water: Part I. Oxidation kinetics and product formation. Water Res. 2003, 37, 1443–1467. [Google Scholar] [CrossRef]
- Wang, D.; Bolton, J.R.; Andrews, S.A.; Hofmann, R. Formation of disinfection by-products in the ultraviolet/chlorine advanced oxidation process. Sci. Total Environ. 2015, 518–519, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hijnen, W.A.M.; Beerendonk, E.F.; Medema, G.J. Inactivation credit of UV radiation for viruses, bacteria and protozoan (oo)cysts in water: A review. Water Res. 2006, 40, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, A.; Mori, M.; Takahashi, A.; Nakano, M.; Wakikawa, N.; Akutagawa, M.; Ikehara, T.; Nakaya, Y.; Kinouchi, Y. New water disinfection system using UVA light-emitting diodes. J. Appl. Microbiol. 2007, 103, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Hamamoto, A.; Takahashi, A.; Nakano, M.; Wakikawa, N.; Tachibana, S.; Ikehara, T.; Nakaya, Y.; Akutagawa, M.; Kinouchi, Y. Development of a new water sterilization device with a 365 nm UV-LED. Med. Biol. Eng. Comput. 2007, 45, 1237–1241. [Google Scholar] [CrossRef] [PubMed]
- Erill, I.; Campoy, S.; Barbe, J. Aeons of distress: An evolutionary perspective on the bacterial SOS response. FEMS Microbiol. Rev. 2007, 31, 637–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selby, C.P.; Sancar, A. A cryptochrome/photolyase class of enzymes with single-stranded DNA-specific photolyase activity. Proc. Natl. Acad. Sci. USA 2006, 103, 17696–17700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamamoto, A.; Bandou, C.; Nakano, M.; Mawatari, K.; Lian, X.; Yamato, M.; Harada, N.; Akutagawa, M.; Kinouchi, Y.; Nakaya, Y.; et al. Differences in stress response after UVC or UVA irradiation in Vibrio parahaemolyticus. Environ. Microbiol. Rep. 2010, 2, 660–666. [Google Scholar] [CrossRef]
- Nakahashi, M.; Mawatari, K.; Hirata, A.; Maetani, M.; Shimohata, T.; Uebanso, T.; Hamada, Y.; Akutagawa, M.; Kinouchi, Y.; Takahashi, A. Simultaneous irradiation with different wavelengths of ultraviolet light has synergistic bactericidal effect on Vibrio parahaemolyticus. Photochem. Photobiol. 2014, 90, 1397–1403. [Google Scholar] [CrossRef]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of V. cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- Nakano, M.; Takahashi, A.; Su, Z.; Harada, N.; Mawatari, K.; Nakaya, Y. Hfq regulates the expression of the thermostable direct hemolysin gene in Vibrio parahaemolyticus. BMC Microbiol. 2008, 8, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aussel, L.; Loiseau, L.; Hajj Chehade, M.; Pocachard, B.; Fontecave, M.; Pierrel, F.; Barras, F. ubiJ, a new gene required for aerobic growth and proliferation in macrophage, is involved in coenzyme Q biosynthesis in Escherichia coli and Salmonella enterica serovar Typhimurium. J. Bacteriol. 2014, 196, 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexei, V.; Elke, K.M.; Zoltan, N. Serine biosynthesis with one carbon catabolism and the glycine cleavage system represents a novel pathway for ATP generation. PLoS ONE 2011, 6, e25881. [Google Scholar] [CrossRef] [Green Version]
- Lim, B.R.; Ahn, K.H.; Songprasert, P.; Cho, J.W.; Lee, S.H. Microbial community structure of membrane fouling film in an intermittently and continuously aerated submerged membrane bioreactor treating domestic wastewater. Water Sci. Technol. 2004, 49, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Aussel, L.; Pierrel, F.; Loiseau, L.; Lombard, M.; Fontecave, M.; Barras, F. Biosynthesis and physiology of coenzyme Q in bacteria. Biochim. Biophys. Acta 2014, 1837, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Usukura, E.; Suzuki, T.; Furuike, S.; Soga, N.; Saita, E.; Hisabori, T.; Kinosita, K., Jr.; Yoshida, M. Torque generation and utilization in motor enzyme F0F1-ATP synthase: Half-torque F1 with short-sized pushrod helix and reduced ATP Synthesis by half-torque F0F1. J. Biol. Chem. 2012, 287, 1884–1891. [Google Scholar] [CrossRef] [Green Version]
- Bosshard, M.; Bucheli, M.; Meur, Y.; Egli, T. The respiratory chain is the cell’s Achilles’ heel during UVA inactivation in Escherichia coli. Microbiology 2010, 150, 2006–2015. [Google Scholar] [CrossRef] [Green Version]
- Bosshard, F.; Riedel, K.; Schneider, T.; Geiser, C.; Bucheli, M.; Egli, T. Protein oxidation and aggregation in UVA-irradiated Escherichia coli cells as signs of accelerated cellular senescence. Environ. Microbiol. 2010, 12, 2931–2945. [Google Scholar] [CrossRef]
- Akhova, A.V.; Tkachenko, A.G. ATP/ADP alteration as a sign of the oxidative stress development in Escherichia coli cells under antibiotic treatment. FEMS. Microbiol. Lett. 2014, 353, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Green, E.R.; Mecsas, J. Bacterial secretion systems: An overview. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Sikora, A.E.; Lybarger, S.R.; Sandkvist, M. Compromised outer membrane integrity in Vibrio cholerae Type II secretion mutants. J. Bacteriol. 2007, 189, 8484–8495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandkvist, M.; Michel, L.O.; Hough, L.P.; Morales, V.M.; Bagdasarian, M.; Koomey, M.; DiRita, V.J.; Bagdasarian, M. General secretion pathway (eps) genes required for toxin secretion and outer membrane biogenesis in Vibrio cholerae. J. Bacteriol. 1997, 179, 6994–7003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikora, A.E.; Beyhan, S.; Bagdasarian, M.; Yildiz, F.H.; Sandkvist, M. Cell envelope perturbation induces oxidative stress and changes in iron homeostasis in Vibrio cholerae. J. Bacteriol. 2009, 191, 5398–5408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitter, W.; van Boxtel, R.; Groeneweg, M.; Carballo, P.S.; Zahringer, U.; Tommassen, J.; Koster, M. Species-specific functioning of the Pseudomonas XcpQ secretin: Role for the C-terminal homology domain and lipopolysaccharide. J. Bacteriol. 2007, 189, 2967–2975. [Google Scholar] [CrossRef] [Green Version]
- Michel, G.; Ball, G.; Goldberg, J.B.; Lazdunski, A. Alteration of the lipopolysaccharide structure affects the functioning of the Xcp secretory system in Pseudomonas aeruginosa. J. Bacteriol. 2000, 182, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.L.; Fong, J.C.; Rule, C.; Rogers, A.; Yildiz, F.H.; Sandkvist, M. The Type II secretion system delivers matrix proteins for biofilm formation by Vibrio cholerae. J. Bacteriol. 2014, 196, 4245–4252. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, F.H.; Visick, K.L. Vibrio biofilms: So much the same yet so different. Trends Microbiol. 2009, 17, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Mann, E.E.; Wozniak, D.J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Pezzoni, M.; Pizarro, R.A.; Costa, C.S. Protective role of extracellular catalase (KatA) against UVA radiation in Pseudomonas aeruginosa biofilms. J. Photochem. Photobiol. B 2014, 131, 53–64. [Google Scholar] [CrossRef]
- Søballe, B.; Poole, R.K. Microbial ubiquinones: Multiple roles in respiration, gene regulation and oxidative stress management. Microbiology 1999, 145, 1817–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, B.J.; Young, I.G. Role of quinones in electron transport to oxygen and nitrate in Escherichia coli. Studies with a ubiA- menA- double quinone mutant. Biochim. Biophys. Acta 1977, 461, 84–100. [Google Scholar] [CrossRef]
- Sharma, P.; Teixeira de Mattos, M.J.; Hellingwerf, K.J.; Bekker, M. On the function of the various quinone species in Escherichia coli. FEBS J. 2012, 279, 3364–3373. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, S.; Schoepp-Cothenet, B.; Ceccaldi, P.; Guigliarelli, B.; Magalon, A. The prokaryotic Mo/W-bisPGD enzymes family: A catalytic workhorse in bioenergetic. Biochim. Biophys. Acta 2013, 1827, 1048–1085. [Google Scholar] [CrossRef] [Green Version]
- Meganathan, R. Ubiquinone biosynthesis in microorganisms. FEMS Microbiol. Lett. 2001, 203, 131–139. [Google Scholar] [CrossRef]
- Søballe, B.; Poole, R.K. Ubiquinone limits oxidative stress in Escherichia coli. Microbiology 2000, 146, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Gulmezian, M.; Hyman, K.R.; Marbois, B.N.; Clarke, C.F.; Javor, G.T. The role of UbiX in Escherichia coli coenzyme Q biosynthesis. Arch. Biochem. Biophys. 2007, 467, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Poon, W.W.; Davis, D.E.; Ha, H.T.; Jonassen, T.; Rather, P.N.; Clarke, C.F. Identification of Escherichia coli ubiB, a gene required for the first monooxygenase step in ubiquinone biosynthesis. J. Bacteriol. 2000, 182, 5139–5146. [Google Scholar] [CrossRef] [Green Version]
- Sekiya, M.; Nakamoto, R.K.; Nakanishi-Matsui, M.; Futai, M. Binding of phytopolyphenol piceatannol disrupts β/γ subunit interactions and rate-limiting step of steady-state rotational catalysis in Escherichia coli F1-ATPase. J. Biol. Chem. 2012, 287, 22771–22780. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain and Plasmid | Description | Source or Reference |
|---|---|---|
| Vibrio parahaemolyticus strains | ||
| RIMD2210633 | KP-positive, serotype O3:K6; clinical isolate; Wild-type strain in this study | Makino et al. [20] |
| VP0097 | RIMD2210633 mutant #0002, Ez-Tn5 DHFR-1 insertion to VP0097, Tmpr | This study |
| VP0136 | RIMD2210633 mutant #0358, Ez-Tn5 DHFR-1 insertion to VP0136, Tmpr | This study |
| VP0140 | RIMD2210633 mutant #0521, Ez-Tn5 DHFR-1 insertion to VP0140, Tmpr | This study |
| VP0315 | RIMD2210633 mutant #0092, Ez-Tn5 DHFR-1 insertion to VP0315, Tmpr | This study |
| VP3069 | RIMD2210633 mutant #0078, Ez-Tn5 DHFR-1 insertion to VP3069, Tmpr | This study |
| VP3075 | RIMD2210633 mutant #0229, Ez-Tn5 DHFR-1 insertion to VP3075, Tmpr | This study |
| Plasmids | ||
| pSA19CP | Expression vector; Cmr | Nakano et al. [21] |
| pVP0097 | pSA19CP expressing VP0097, controlled by the tdhA promoter; Cmr | This study |
| pVP0136 | pSA19CP expressing VP0136, controlled by the tdhA promoter; Cmr | This study |
| pVP0140 | pSA19CP expressing VP0140, controlled by the tdhA promoter; Cmr | This study |
| pVP0315 | pSA19CP expressing VP0315, controlled by the tdhA promoter; Cmr | This study |
| pVP3069 | pSA19CP expressing VP3069, controlled by the tdhA promoter; Cmr | This study |
| Gene | Sequence (5′–3′) | |
|---|---|---|
| Forward | Reverse | |
| VP0097 | 5′–TCTAGA ATGACGCCAGCAGAATTAAAGC–3′ | 5′–GAATTC CTATTGACGATAAGCTCGCCAAC–3′ |
| VP0136 | 5′–TCTAGA ATGAAATTTAAGCGTAGTAAGC–3′ | 5′–GAATTC TTATTGGAAGTCTTGCATGTTCCA–3′ |
| VP0140 | 5′–TCTAGA ATGGCTAATCGTCAGCGCGGT–3′ | 5′–GAATTC CTACTCAGCCGAACGGTCAGAAA–3′ |
| VP0315 | 5′–TCTAGA ATGCACAACAAAATACAACCC–3′ | 5′–GGATCC TCACGAGCGCTGGTCATA–3′ |
| VP3069 | 5′–GGATCC ATGGCTACAGGTAAGATCGTAC–3′ | 5′–GAATTC TTATAGCTTCTTCGCATTCTCG–3′ |
| VP3075 | 5′–TCTAGA ATGGCTGCGCCAGGTGAA–3′ | 5′–GAATTC TTAATGATCAGAGTCTTCGTGTGC–3′ |
| Genes | Orthologous Genes | Product | Functional Classification |
|---|---|---|---|
| VP3075 | atpB | ATP synthase F0F1 subunit alpha | Oxidative phosphorylation F-type H+-transporting ATPase |
| VP3069 | atpD | ATP synthase F0F1 subunit beta | |
| VP0136 | gspG | general secretion pathway protein G | Bacterial secretion system Type II secretion system |
| VP0140 | gspK | general secretion pathway protein K | |
| VP0097 | ubiB | ubiquinone biosynthesis protein UbiB | Ubiquinone and other Terpenoid-quinone biosynthesis |
| VP0315 | ubiX | 3-polyprenyl-4-hydroxybenzoate carboxy-lyase UbiX |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maetani-Yasui, M.; Mawatari, K.; Honjo, A.; Bui, T.K.N.; Shimohata, T.; Uebanso, T.; Aihara, M.; Emoto, T.; Akutagawa, M.; Kinouchi, Y.; et al. Identification of Genes Associated with Sensitivity to Ultraviolet A (UVA) Irradiation by Transposon Mutagenesis of Vibrio parahaemolyticus. Appl. Sci. 2020, 10, 5549. https://doi.org/10.3390/app10165549
Maetani-Yasui M, Mawatari K, Honjo A, Bui TKN, Shimohata T, Uebanso T, Aihara M, Emoto T, Akutagawa M, Kinouchi Y, et al. Identification of Genes Associated with Sensitivity to Ultraviolet A (UVA) Irradiation by Transposon Mutagenesis of Vibrio parahaemolyticus. Applied Sciences. 2020; 10(16):5549. https://doi.org/10.3390/app10165549
Chicago/Turabian StyleMaetani-Yasui, Miki, Kazuaki Mawatari, Airi Honjo, Thi Kim Ngan Bui, Takaaki Shimohata, Takashi Uebanso, Mutsumi Aihara, Takahiro Emoto, Masatake Akutagawa, Yohsuke Kinouchi, and et al. 2020. "Identification of Genes Associated with Sensitivity to Ultraviolet A (UVA) Irradiation by Transposon Mutagenesis of Vibrio parahaemolyticus" Applied Sciences 10, no. 16: 5549. https://doi.org/10.3390/app10165549