
Upper Triassic Carbonate Records: Insights from the Most Complete Panthalassan Platform (Lime Peak, Yukon, Canada)


Abstract
:1. Introduction
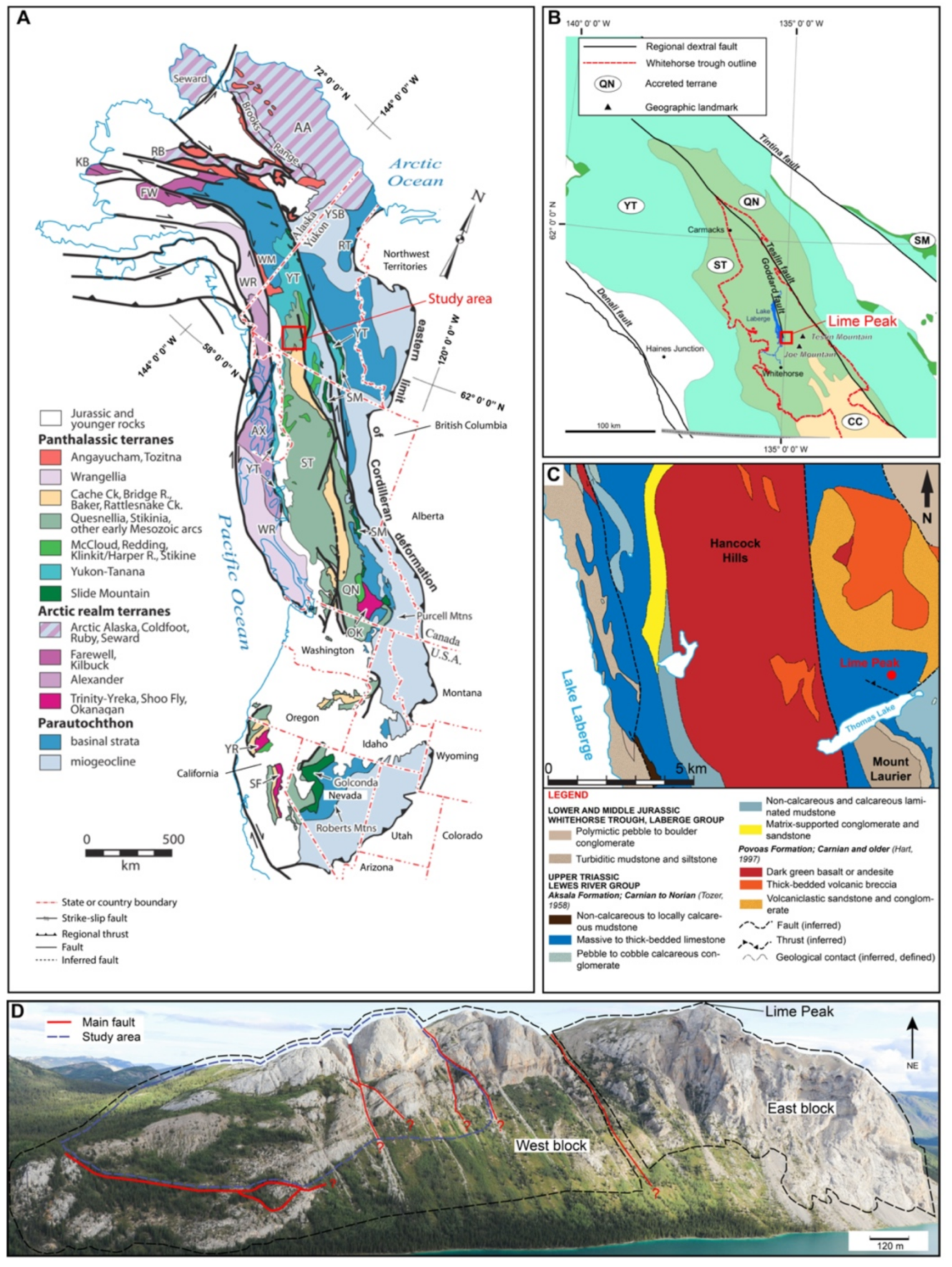
2. Geological Context

3. Area of Study, Mode of Occurrence and Lithofacies Description
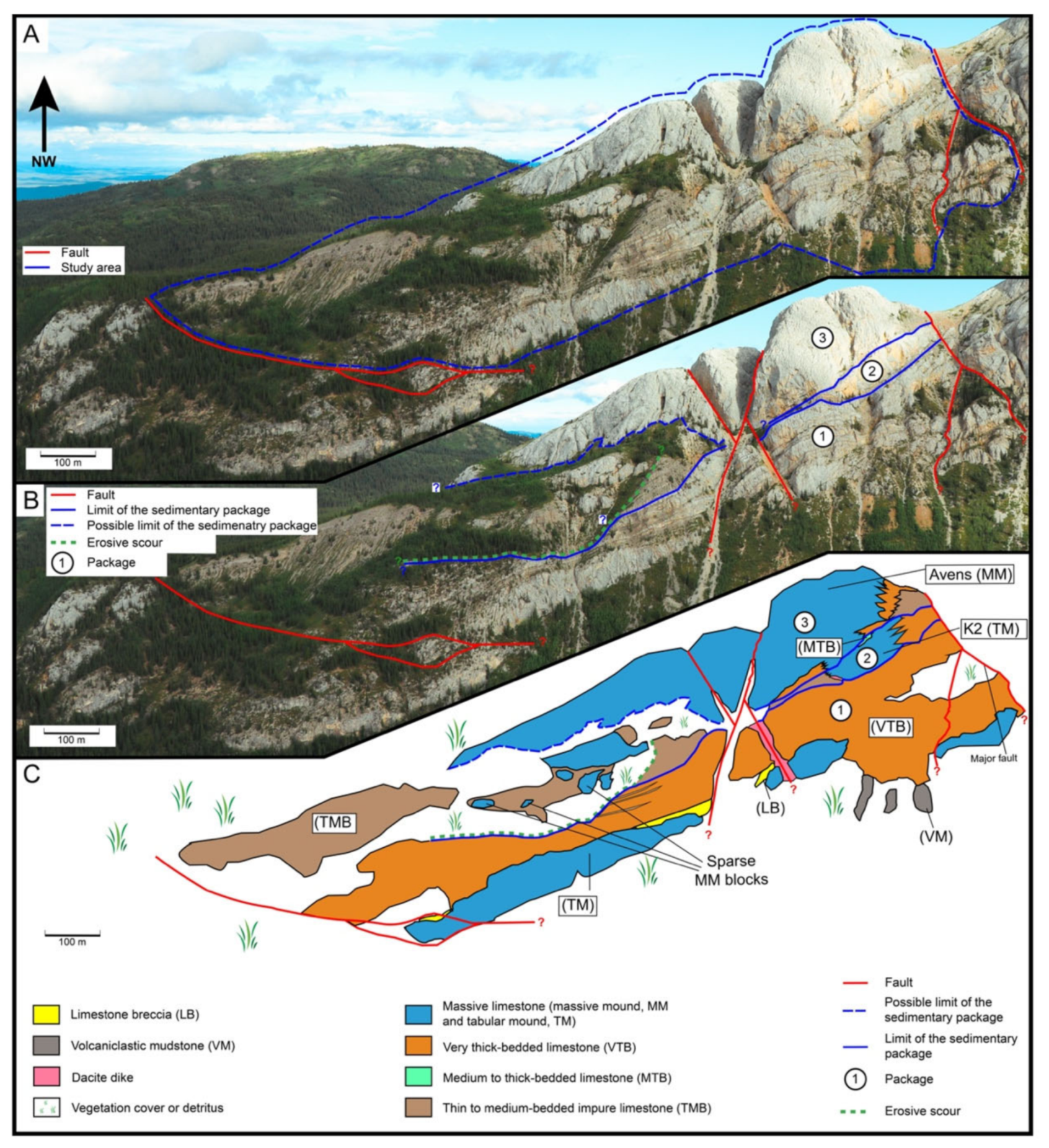
3.1. Area of Study
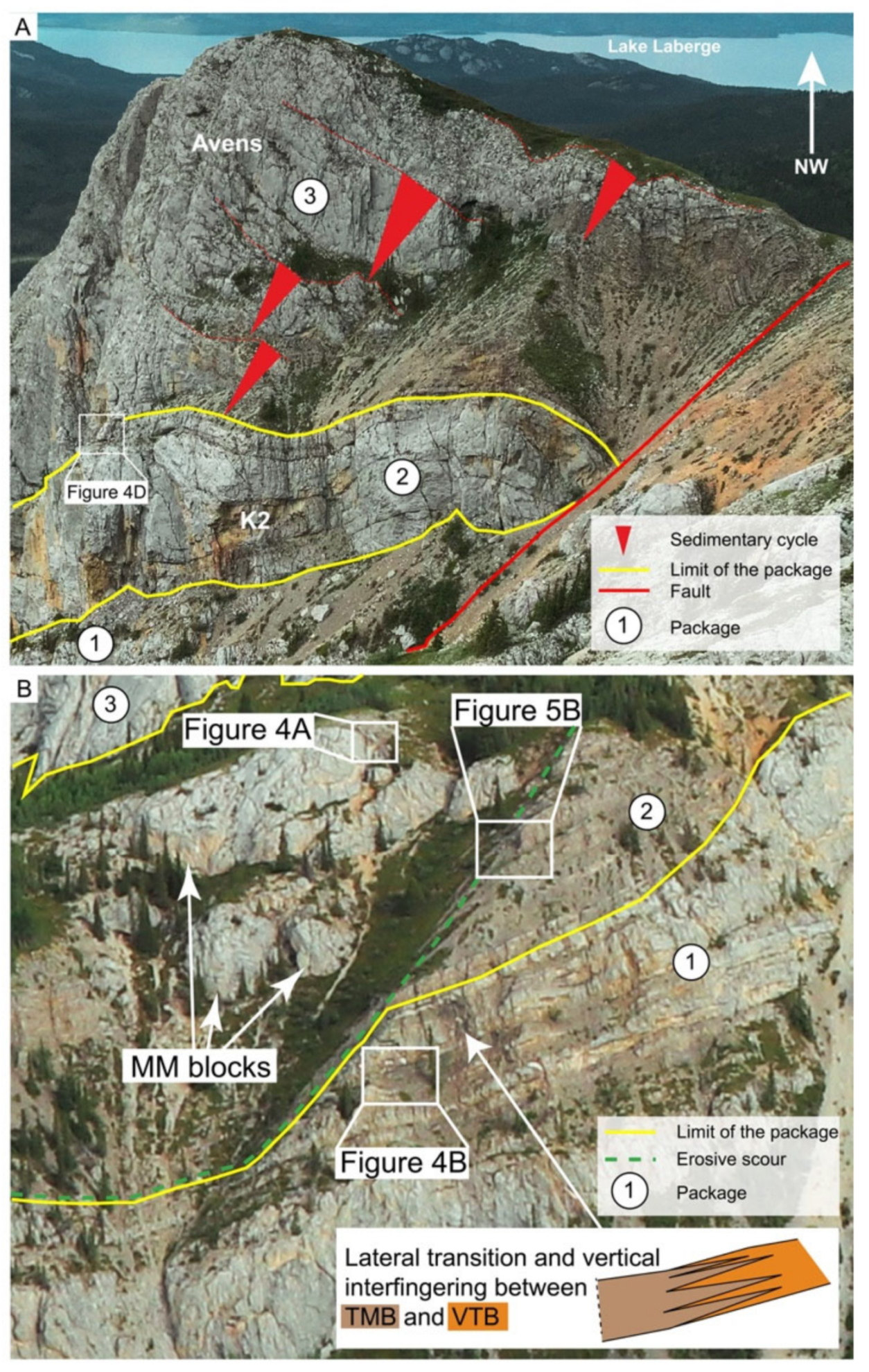
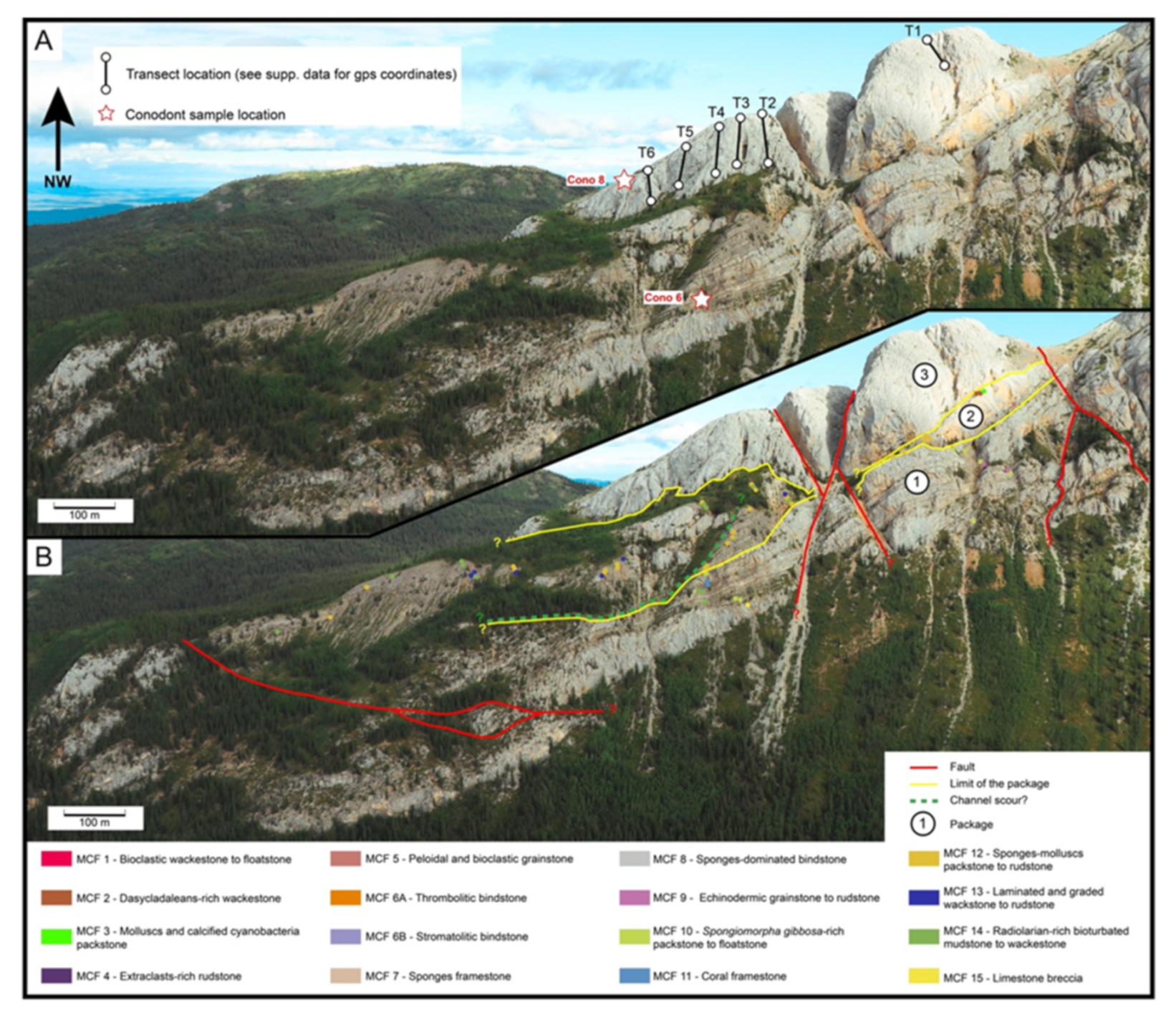
3.2. Mode of Occurrence and Geometries
3.3. Lithofacies Description and Spatial Occurrence
3.3.1. Lithofacies of the First Package
3.3.2. Lithofacies of the Second Package
3.3.3. Lithofacies of the Third Package
4. Methods
5. Age of the Deposits
6. Results
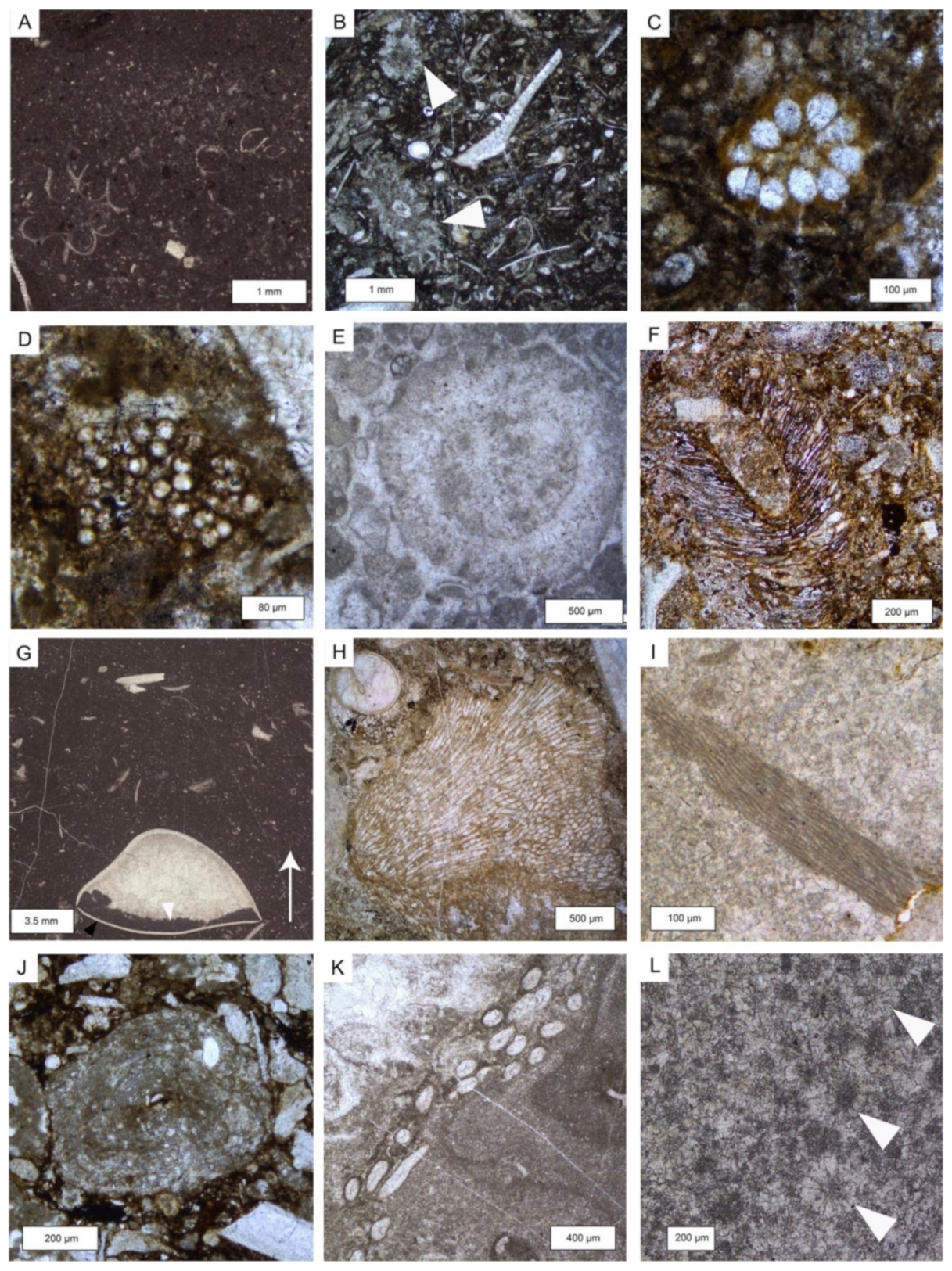
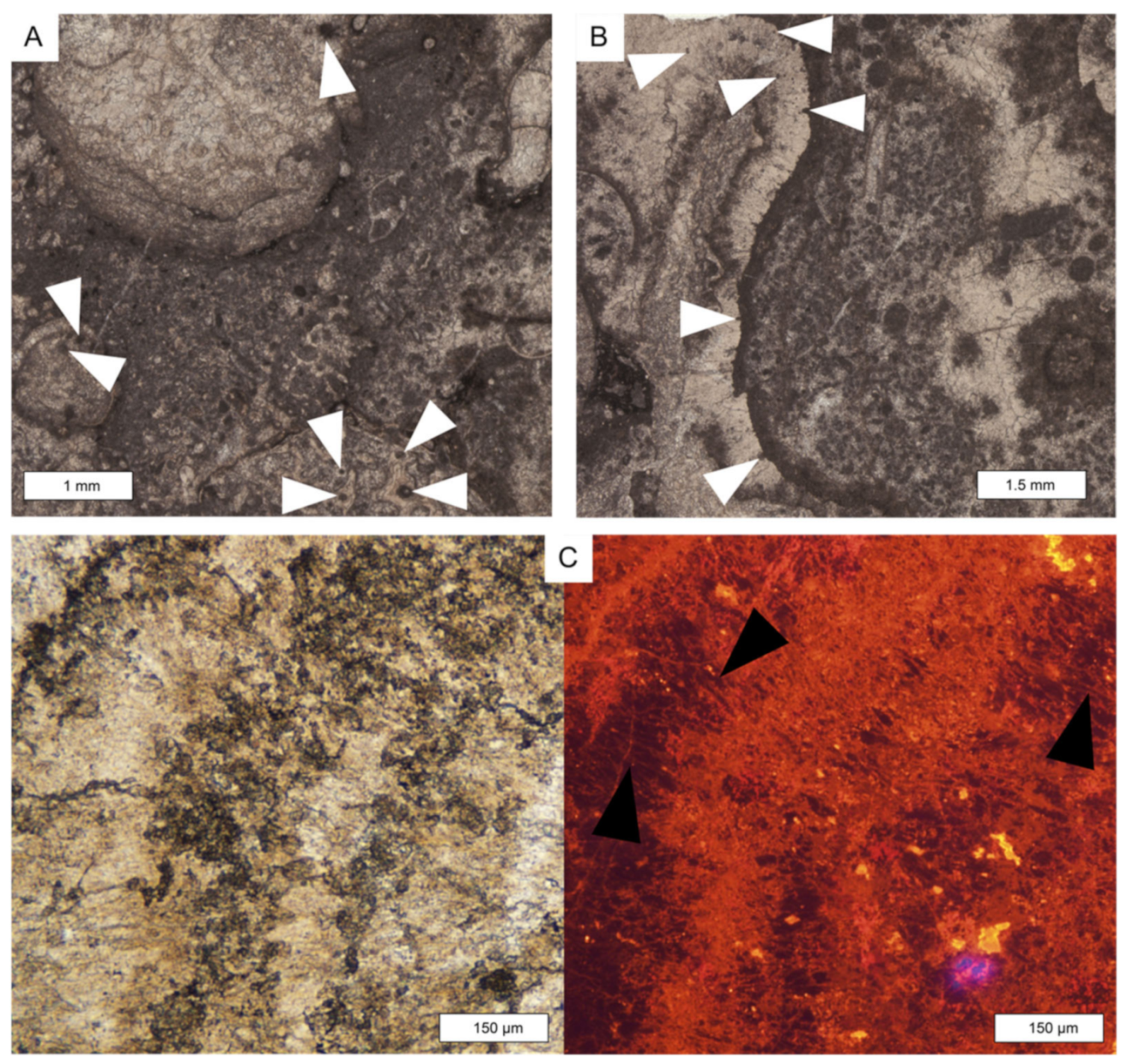
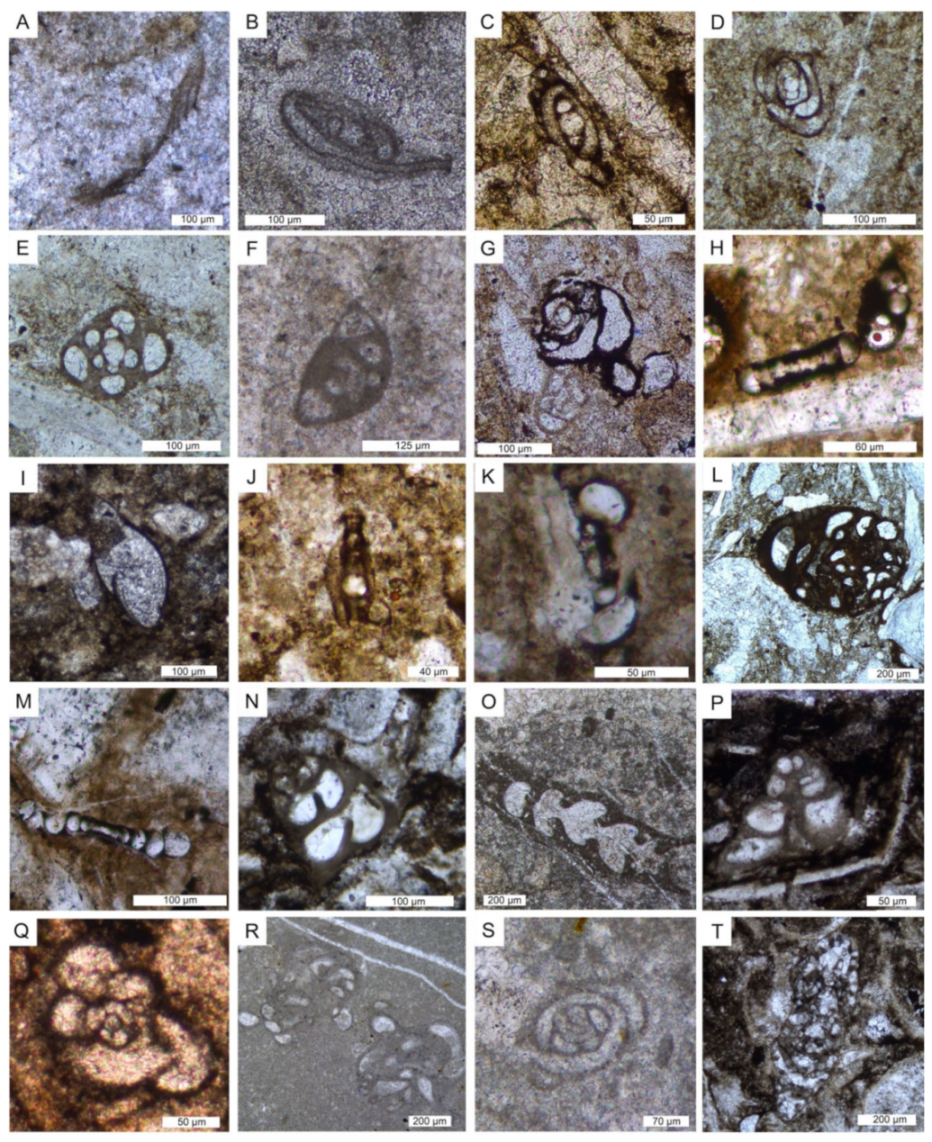
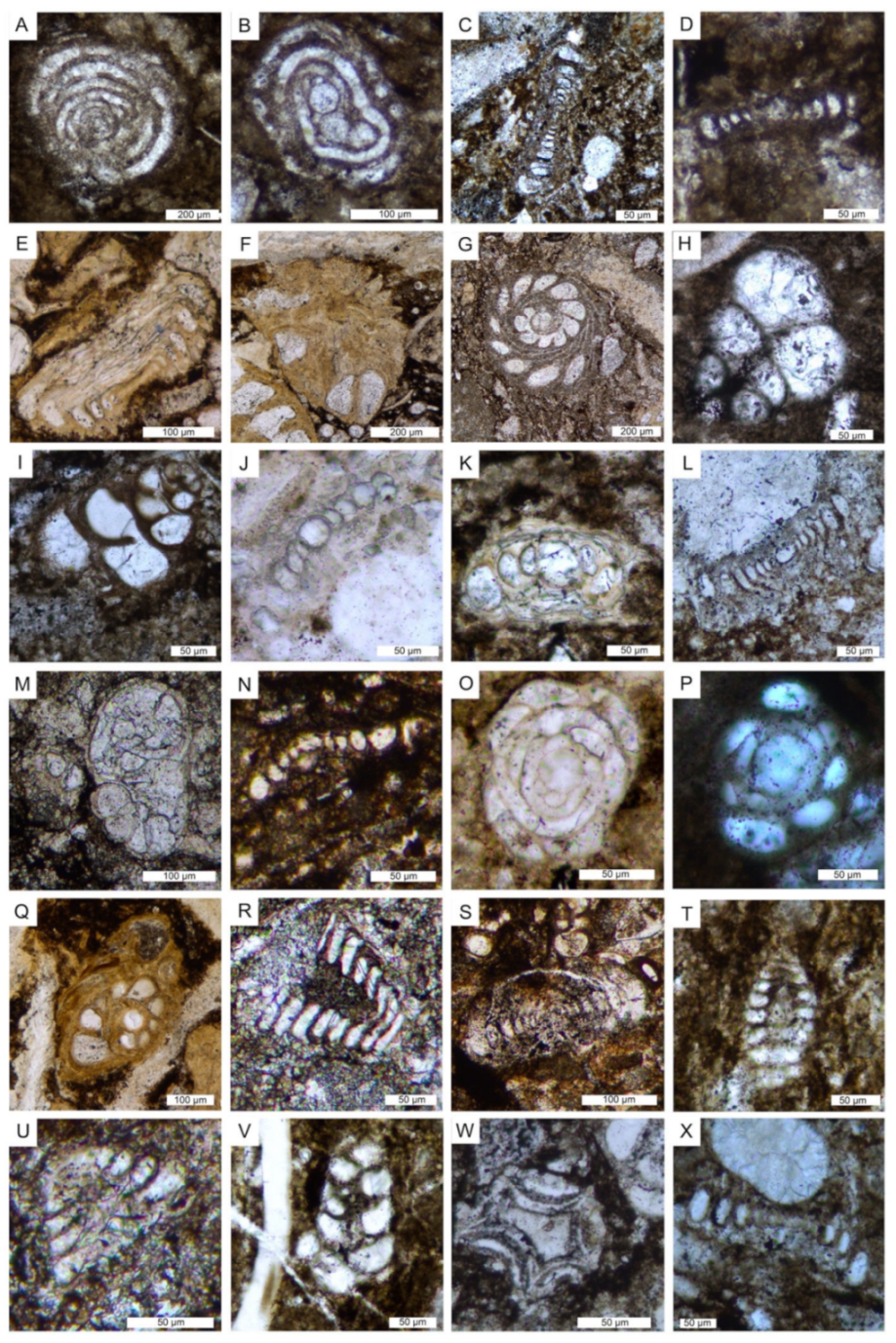
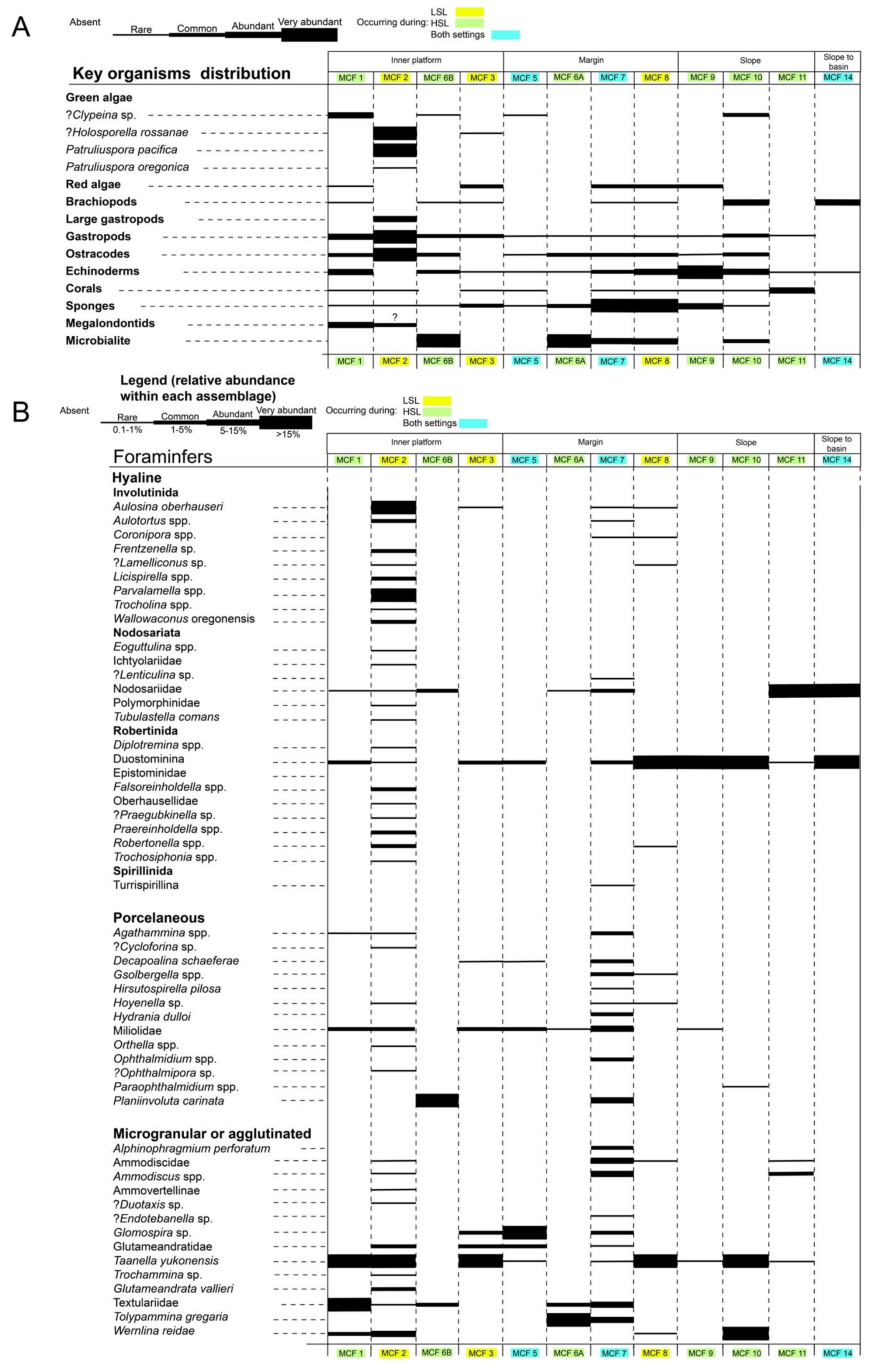
6.1. Microfacies Description and Biotic Content
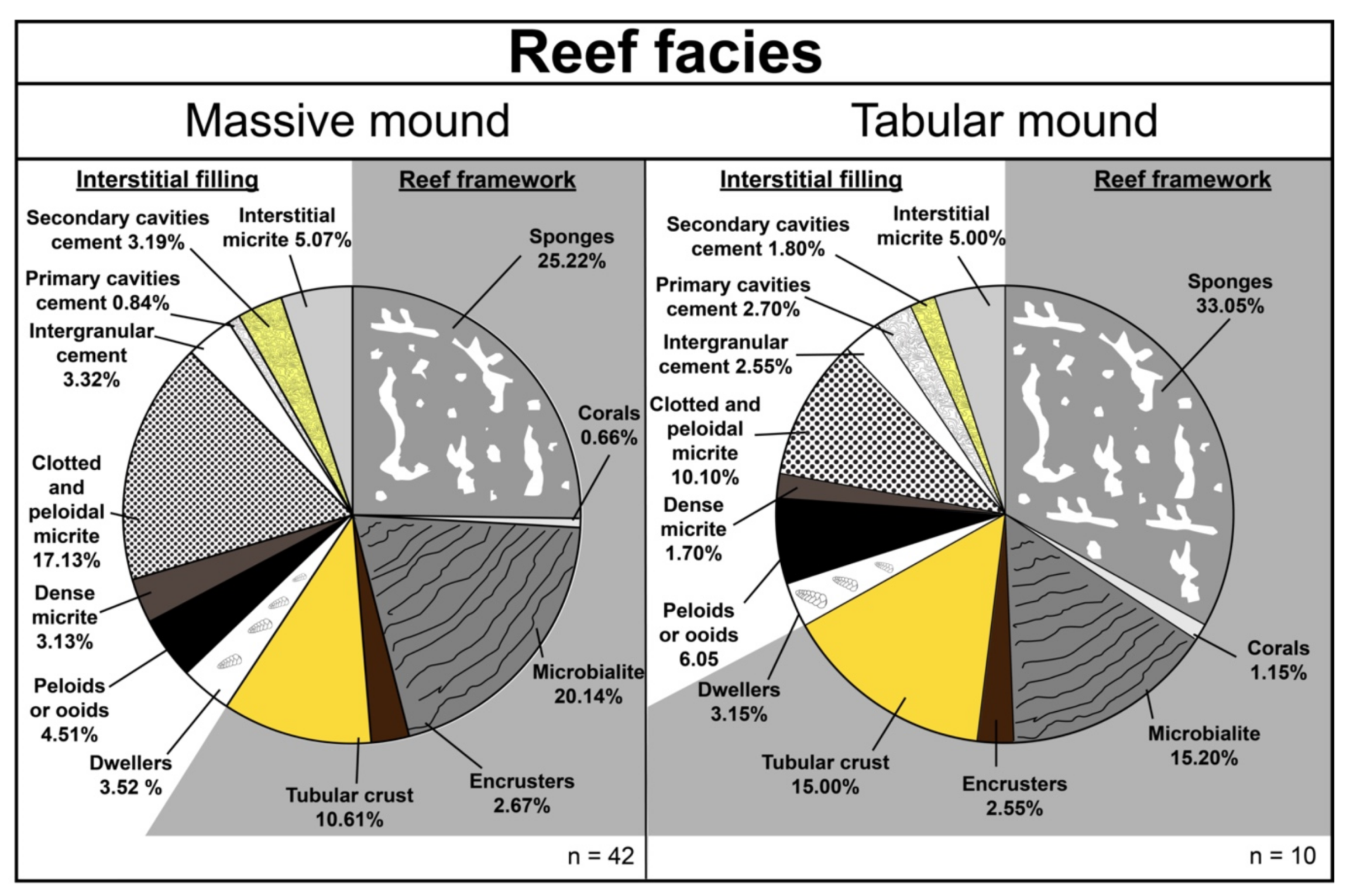
6.2. Composition of the Reef Facies
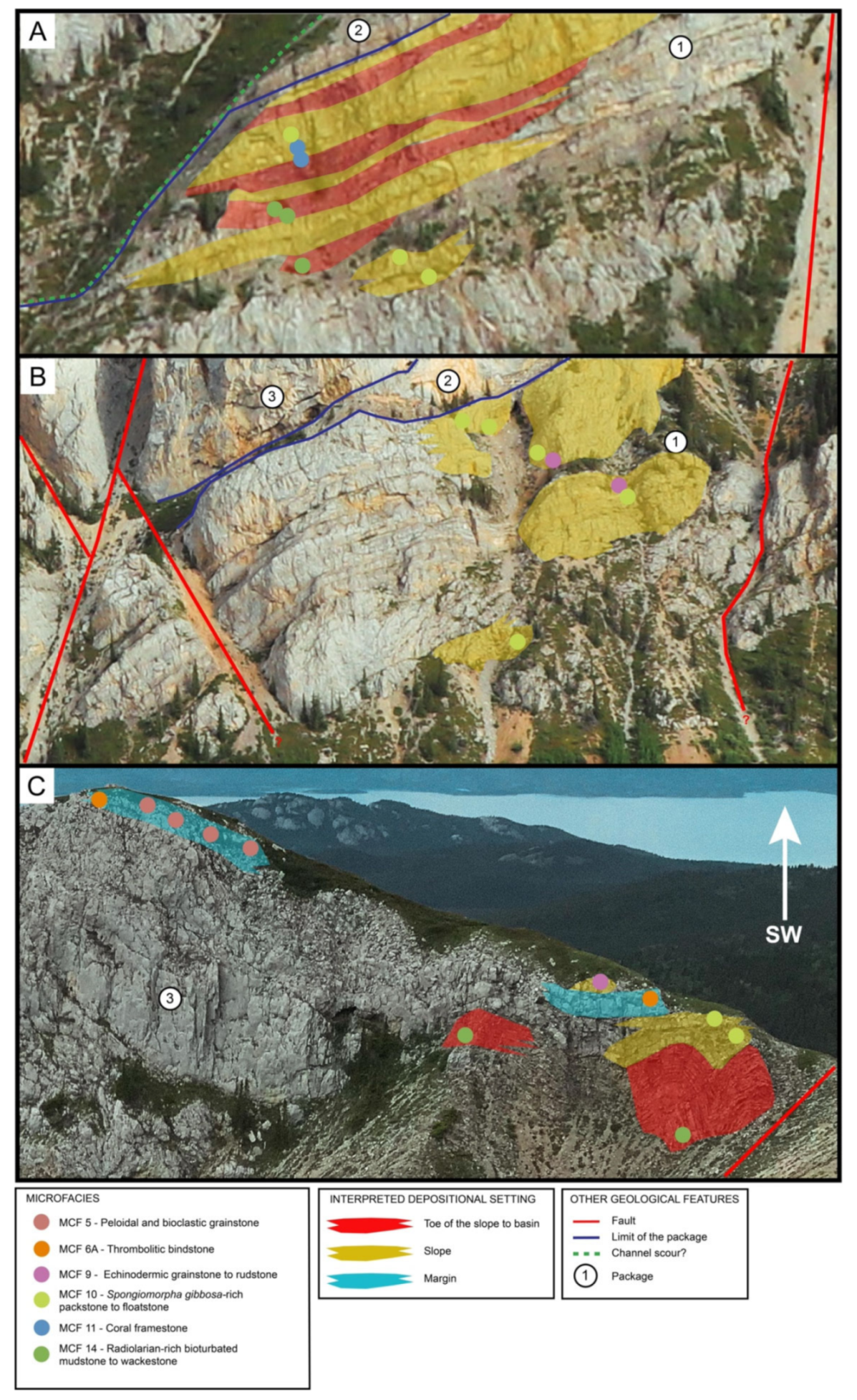
7. Interpretation of the Depositional Environments
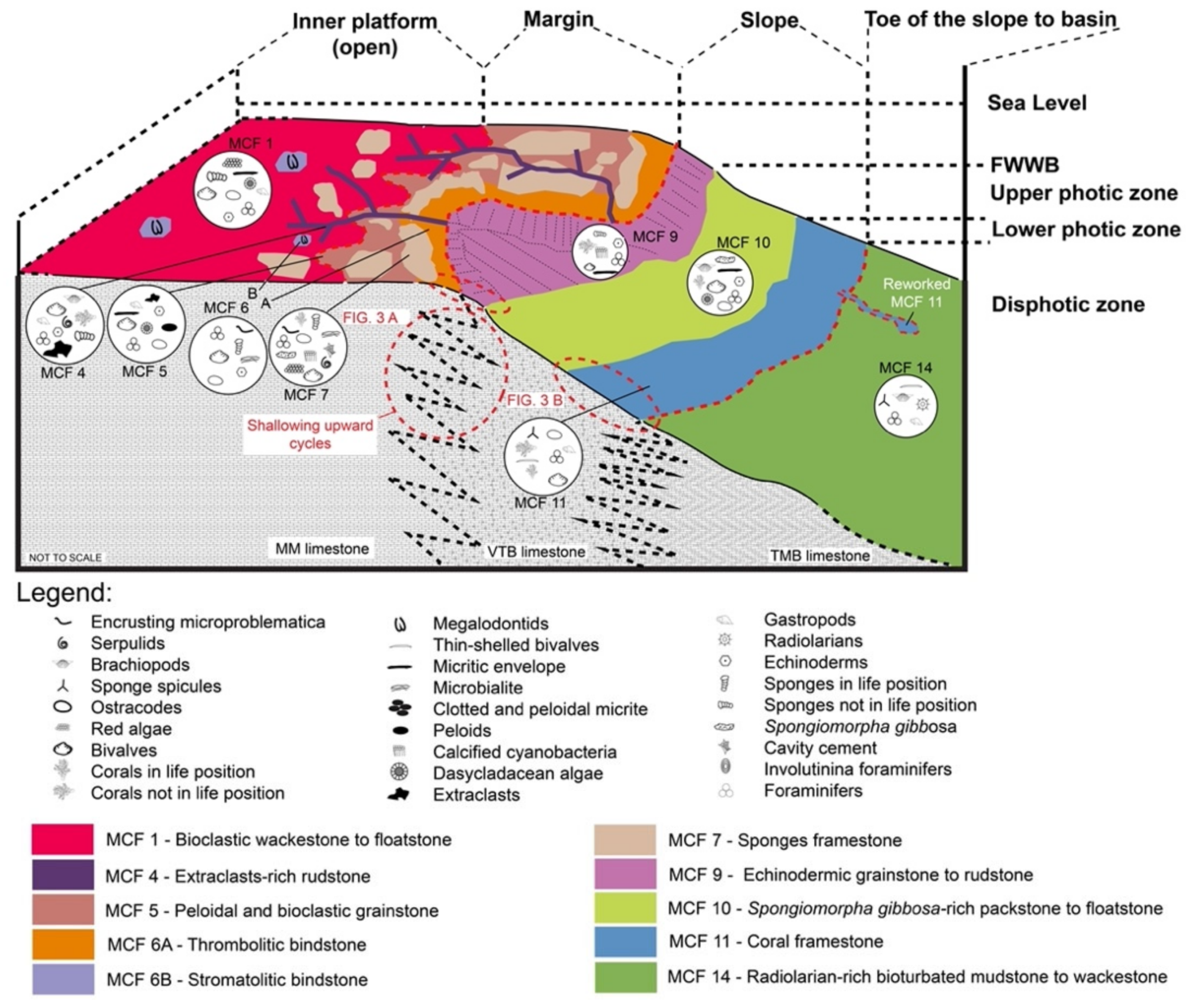
7.1. Depositional Environments during Sedimentation of the First and Third Packages
7.1.1. Toe of the Slope-to-Basin and Slope Environments
7.1.2. Platform Margin Environment
7.1.3. Inner Platform
7.1.4. Depositional Model for the First and Third Packages
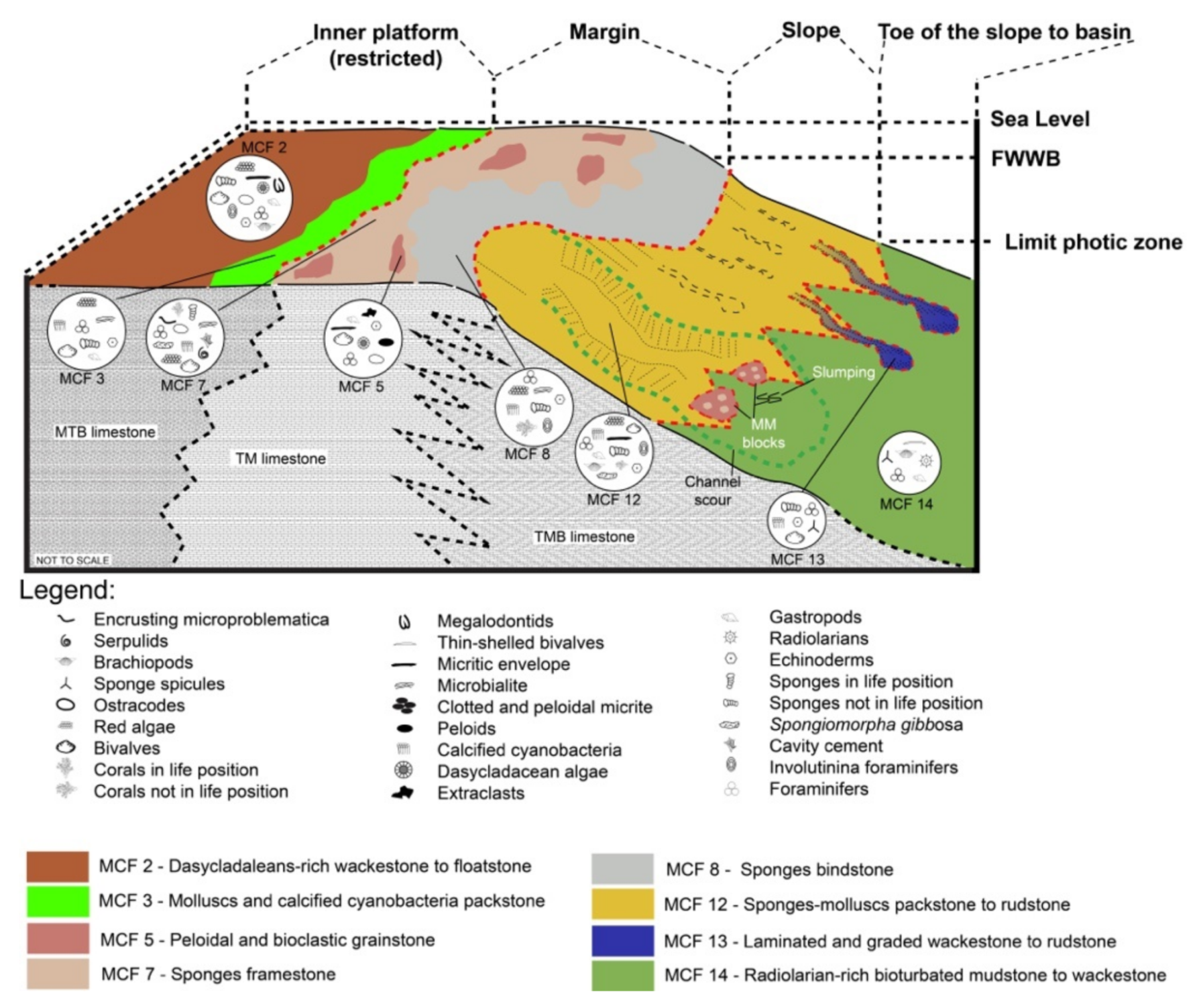
7.2. Depositional Environments during Deposition of the Second Package
7.2.1. Slope and Toe of the Slope-to-Basin Environments
7.2.2. Platform Margin
7.2.3. Inner Platform
7.2.4. Deposition of the Second Package: A Different Tale?
8. Discussion
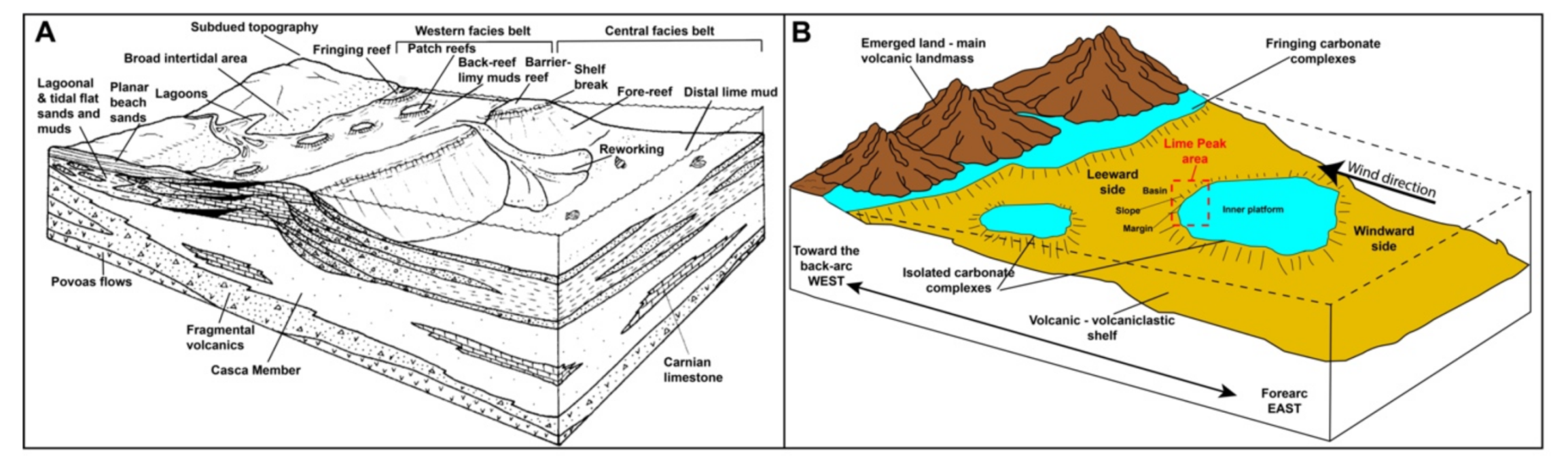
8.1. What Is Lime Peak? New Insights Regarding Its Nature and Spatial Relationship with Other Upper Triassic Carbonates in the Whitehorse Region
8.2. Tectonic Influence on the Sedimentation at Lime Peak
8.3. Evinosponges at Lime Peak
8.4. Carbonate Factory at Lime Peak
8.5. Organisms’ Distribution within the Lime Peak Carbonate System
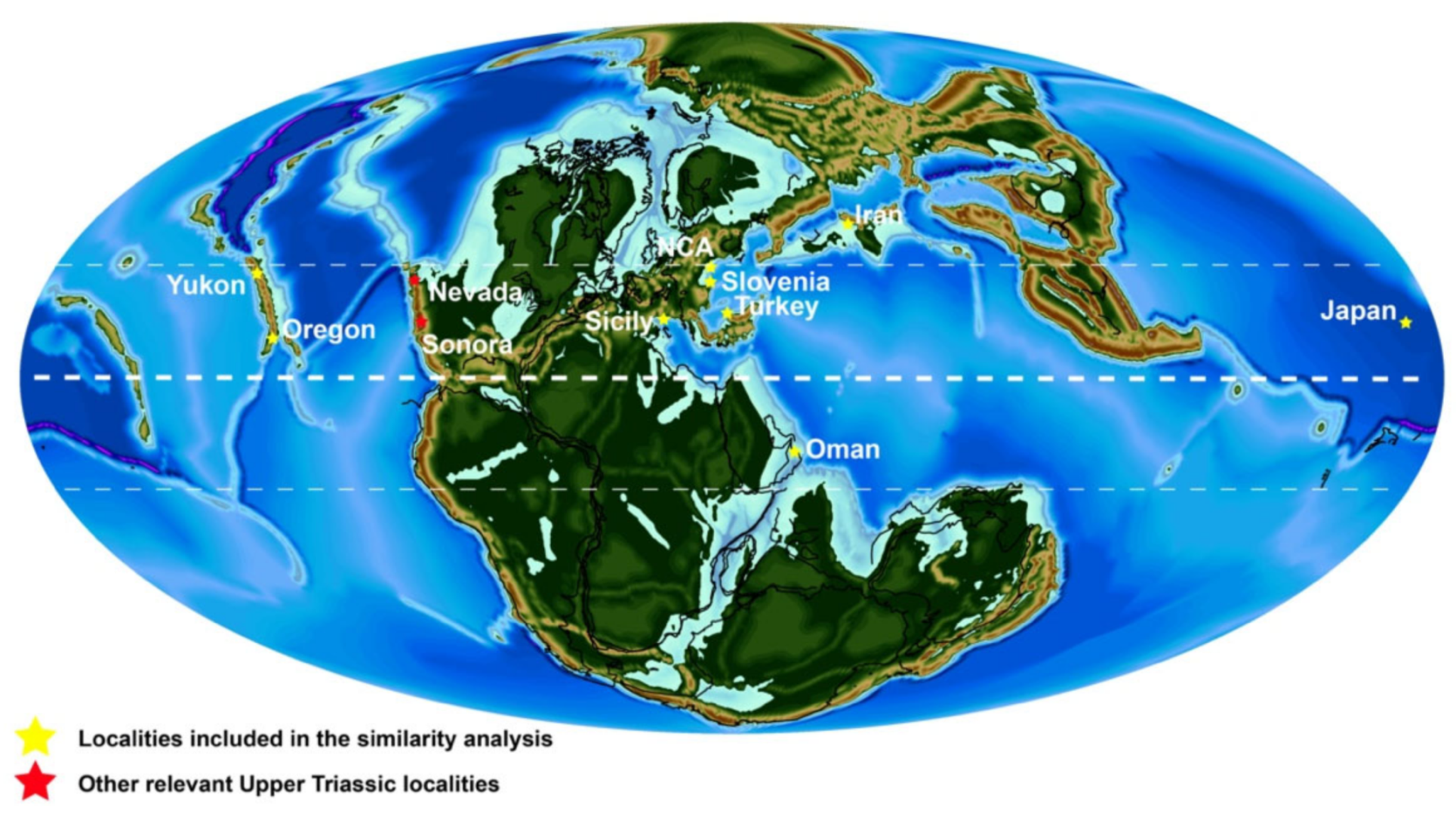
8.6. Reef Paleoecology and the Paleogeography of Stikinia

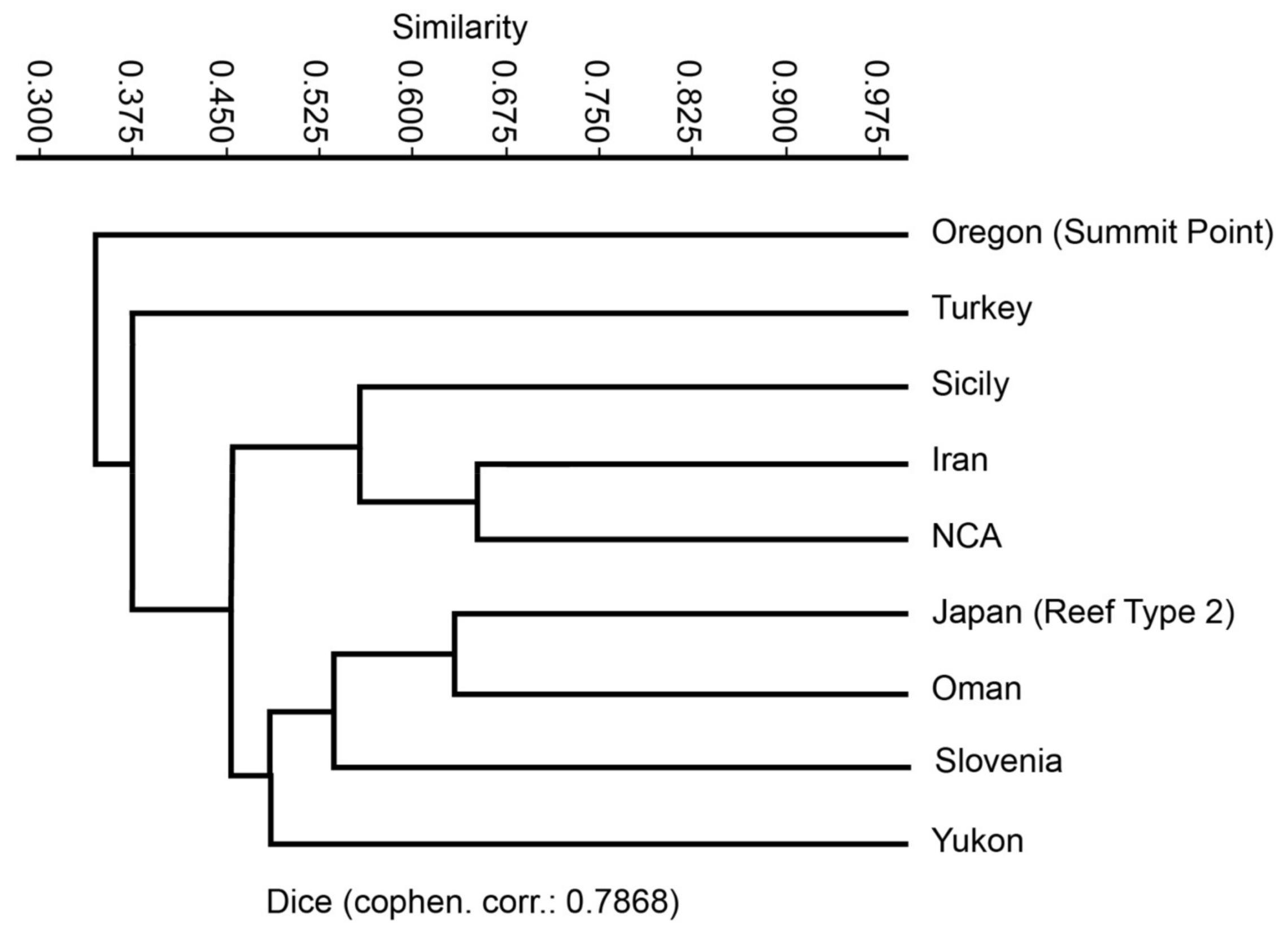
8.7. Comparison with Other Upper Triassic Carbonate Systems Studied during the REEFCaDe Project
8.8. Brief Comparison with Upper Triassic Systems from the Tethys
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral Reef Biodiversity. In Life in the World’s Oceans: Diversity, Distribution, and Abundance; McIntyre, A.D., Ed.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2010; pp. 65–77. ISBN 9781405192972. [Google Scholar]
- Flügel, E. Evolution of Triassic Reefs: Current Concepts and Problems. Facies 1982, 6, 297–327. [Google Scholar] [CrossRef]
- Stanton, R.J.; Flügel, E. An Accretionary Distally Steepened Ramp at an Intrashelf Basin Margin: An Alternative Explanation for the Upper Triassic Steinplatte “Reef” (Northern Calcareous Alps, Austria). Sediment. Geol. 1995, 95, 269–286. [Google Scholar] [CrossRef]
- Chiocchini, M.; Farinacci, A.; Mancinelli, A.; Molinari, V.; Potetti, M. Biostratigrafia a foraminiferi, dasicladali e calpionelle delle successioni carbonatiche mesozoiche dell’Appennino centrale (Italia). In Biostratigrafia dell’Italia Centrale; Mancinelli, A., Ed.; Studi Geologici Camerti, n. speciale; Università di Camerino: Camerino, Italy, 1994; pp. 9–129. [Google Scholar] [CrossRef]
- Galli, M.T.; Jadoul, F.; Bernasconi, S.M.; Cirilli, S.; Weissert, H. Stratigraphy and palaeoenvironmental analysis of the Triassic–Jurassic transition in the western Southern Alps (Northern Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 244, 52–70. [Google Scholar] [CrossRef]
- Romano, R.; Masetti, D.; Carras, N.; Barattolo, F.; Roghi, G. The Triassic/Jurassic boundary in a peritidal carbonate platform of the Pelagonian Domain: The Mount Messapion section (Chalkida, Greece). Riv. Ital. Paleontol. Stratigr. 2008, 114, 431–452. [Google Scholar] [CrossRef]
- Tunaboylu, B.; Altiner, D.; Isintek, I.; Demirci, D. Foraminiferal biostratigraphy and sequence stratigraphy of peritidal carbonates at the Triassic-Jurassic boundary (Karaburun Peninsula, Western Turkey). J. Asian Earth Sci. 2014, 90, 61–76. [Google Scholar] [CrossRef]
- Todaro, S.; Di Stefano, P.; Zarcone, G.; Randazzo, V. Facies stacking and extinctions across the Triassic–Jurassic boundary in a peritidal succession from western Sicily. Facies 2017, 63, 20. [Google Scholar] [CrossRef]
- Martindale, R.C.; Krystyn, L.; Bottjer, D.J.; Corsetti, F.A.; Senowbari-Daryan, B.; Martini, R. Depth Transect of an Upper Triassic (Rhaetian) Reef from Gosau, Austria: Microfacies and Community Ecology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 376, 1–21. [Google Scholar] [CrossRef]
- Marangon, A.; Gattolin, G.; Della Porta, G.; Preto, N. The Latemar: A Flat-Topped, Steep Fronted Platform Dominated by Microbialites and Synsedimentary Cements. Sediment. Geol. 2011, 240, 97–114. [Google Scholar] [CrossRef]
- Jin, X.; Shi, Z.; Rigo, M.; Franceschi, M.; Preto, N. Carbonate Platform Crisis in the Carnian (Late Triassic) of Hanwang (Sichuan Basin, South China): Insights from Conodonts and Stable Isotope Data. J. Asian Earth Sci. 2018, 164, 104–124. [Google Scholar] [CrossRef]
- Bernecker, M. Late Triassic Reefs from the Northwest and South Tethys: Distribution, Setting, and Biotic Composition. Facies 2005, 51, 442–453. [Google Scholar] [CrossRef]
- Schäfer, P.; Senowbari-Daryan, B. Facies Development and Paleoecologic Zonation of Four Upper Triassic Patch-Reefs, Northern Calcareous Alps near Salzburg, Austria. In European Fossil Reef Models; Toomey, D.F., Ed.; SEPM Society for Sedimentary Geology: Tulsa, OK, USA, 1981; Volume 30, pp. 241–259. [Google Scholar]
- Senowbari-Daryan, B.; Rashidi, K.; Torabi, H. Foraminifera and Their Associations of a Possibly Rhaetian Section of the Nayband Formation in Central Iran, Northeast of Esfahan. Facies 2010, 56, 567–596. [Google Scholar] [CrossRef]
- Ziegler, A.M.; Eshel, G.; McAllister Rees, P.; Rothfus, T.A.; Rowley, D.B.; Sunderlin, D. Tracing the Tropics across Land and Sea: Permian to Present. Lethaia 2003, 36, 227–254. [Google Scholar] [CrossRef] [Green Version]
- Flügel, E. Triassic Reef Patterns. In Phanerozoic Reef Patterns; Kiessling, W., Flugel, E., Golonka, J., Eds.; Society of Economic Paleontologists and Mineralogists: Tulsa, OK, USA, 2002; pp. 391–464. [Google Scholar]
- Peyrotty, G.; Rigaud, S.; Kemkin, I.; Martini, R. Sedimentology and Biostratigraphy of Upper Triassic Atoll-Type Carbonates from the Dalnegorsk Area, Taukha Terrane, Far East Russia. Glob. Planet. Chang. 2020, 184, 103072. [Google Scholar] [CrossRef]
- Peybernes, C.; Chablais, J.; Onoue, T.; Martini, R. Mid-Oceanic Shallow-Water Carbonates of the Panthalassa Domain: New Microfacies Data from the Sambosan Accretionary Complex, Shikoku Island, Japan. Facies 2016, 62, 1–27. [Google Scholar] [CrossRef]
- Mitchell, A.H.G. Facies of an Early Miocene Volcanic Arc, Malekula Island, New Hebrides. Sedimentology 1970, 14, 201–243. [Google Scholar] [CrossRef]
- Hamilton, W.B. Plate Tectonics and Island Arcs. Geol. Soc. Am. Bull. 1988, 100, 1503–1527. [Google Scholar] [CrossRef]
- Peyrotty, G.; Brigaud, B.; Martini, R. Δ18O, Δ13C, Trace Elements and REE in Situ Measurements Coupled with U–Pb Ages to Reconstruct the Diagenesis of Upper Triassic Atoll-Type Carbonates from the Panthalassa Ocean. Mar. Pet. Geol. 2020, 120, 104520. [Google Scholar] [CrossRef]
- Peybernes, C.; Chablais, J.; Onoue, T.; Escarguel, G.; Martini, R. Paleoecology, Biogeography, and Evolution of Reef Ecosystems in the Panthalassa Ocean during the Late Triassic: Insights from Reef Limestone of the Sambosan Accretionary Complex, Shikoku, Japan. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 457, 31–51. [Google Scholar] [CrossRef] [Green Version]
- Chablais, J.; Martini, R.; Onoue, T. Aulotortus Friedli from the Upper Triassic Gravitational Flow Deposits of the Kumagawa River (Kyushu, Southwest Japan). Paleontol. Res. 2010, 14, 151–160. [Google Scholar] [CrossRef]
- Chablais, J.; Onoue, T.; Martini, R. Upper Triassic Reef-Limestone Blocks of Southwestern Japan: New Data from a Panthalassan Seamount. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 293, 206–222. [Google Scholar] [CrossRef]
- Chablais, J.; Martini, R.; Samankassou, E.; Onoue, T.; Sano, H. Microfacies and Depositional Setting of the Upper Triassic Mid-Oceanic Atoll-Type Carbonates of the Sambosan Accretionary Complex (Southern Kyushu, Japan). Facies 2010, 56, 249–278. [Google Scholar] [CrossRef]
- Chablais, J.; Martini, R.; Kobayashi, F.; Stampfli, G.M.; Onoue, T. Upper Triassic Foraminifers from Panthalassan Carbonate Buildups of Southwestern Japan and Their Paleobiogeographic Implications. Micropaleontology 2011, 57, 93–124. [Google Scholar] [CrossRef]
- Peybernes, C.; Peyrotty, G.; Chablais, J.; Onoue, T.; Yamashita, D.; Martini, R. Birth and Death of Seamounts in the Panthalassa Ocean: Late Triassic to Early Jurassic Sedimentary Record at Mount Sambosan, Shikoku, Southwest Japan. Glob. Planet. Chang. 2020, 192, 103250. [Google Scholar] [CrossRef]
- Peybernes, C.; Chablais, J.; Martini, R. Upper Triassic (Ladinian?-Carnian) Reef Biota from the Sambosan Accretionary Complex, Shikoku, Japan. Facies 2015, 61, 20. [Google Scholar] [CrossRef]
- Peyrotty, G.; Ueda, H.; Peybernes, C.; Rettori, R.; Martini, R. Upper Triassic Shallow–Water Carbonates from the Naizawa Accretionary Complex, Hokkaido (Japan): New Insights from Panthalassa. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 554, 109832. [Google Scholar] [CrossRef]
- Rigaud, S.; Blau, J.; Martini, R.; Rettori, R. Taxonomy and Phylogeny of the Trocholinidae (Involutinina). J. Foraminifer. Res. 2013, 43, 317–339. [Google Scholar] [CrossRef]
- Rigaud, S.; Vachard, D.; Schlagintweit, F.; Martini, R. New Lineage of Triassic Aragonitic Foraminifera and Reassessment of the Class Nodosariata. J. Syst. Palaeontol. 2016, 14, 919–938. [Google Scholar] [CrossRef]
- Khalil, H.; Baumgartner, P.O.; Onoue, T.; Del Piero, N.; Stanley, G.; Rigaud, S.; Martini, R. Middle-Late Triassic Radiolarian Assemblages from Chert Clasts of the Excelsior Gulch Conglomerate (Wallowa Terrane, Oregon, U.S.A.). Rev. Paleobiol. 2020, 39, 565–579. [Google Scholar] [CrossRef]
- Fucelli, A.; Golding, M.; Martini, R. Downslope Re-Sedimentation from a Short-Living Carbonate Platform: Record from the Upper Triassic Hosselkus Limestone (Northern California). Sediment. Geol. 2021, 422, 105967. [Google Scholar] [CrossRef]
- Rigaud, S.; Martini, R.; Rettori, R.; Stanley, G.D.J. Stratigraphic Potential of the Upper Triassic Benthic Foraminifers. Albertiana 2010, 38, 34–39. [Google Scholar]
- Rigaud, S.; Martini, R.; Rettori, R. Parvalamellinae, a New Subfamily for Triassic Glomospiroid Involutinidae. J. Foraminifer. Res. 2012, 42, 245–256. [Google Scholar] [CrossRef]
- Rigaud, S.; Martini, R.; Rettori, R. A New Genus of Norian Involutinid Foraminifers—Its Morphological, Biostratigraphic, and Evolutionary Significance. Acta Palaeontol. Pol. 2013, 58, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Rigaud, S.; Martini, R.; Vachard, D. Early Evolution and New Classification of the Order Robertinida (Foraminifera). J. Foraminifer. Res. 2015, 45, 3–28. [Google Scholar] [CrossRef]
- Rigaud, S.; Blau, J.; Martini, R.; Rettori, R. Taxonomy, Phylogeny, and Functional Morphology of the Foraminiferal Genus Involutina. Acta Palaeontol. Pol. 2015, 60, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Rigaud, S.; Vachard, D.; Martini, R. Agglutinated versus Microgranular Foraminifers: End of a Paradigm? J. Syst. Palaeontol. 2015, 13, 75–95. [Google Scholar] [CrossRef]
- Rigaud, S.; Martini, R. Agglutinated or Porcelaneous Tests: Where To Draw the Line? J. Foraminifer. Res. 2016, 46, 333–344. [Google Scholar] [CrossRef]
- Khalil, H.; McRoberts, C.A.; Del Piero, N.; Stanley, G.D.J.; Martini, R.; Rigaud, S. New Biostratigraphic Constraints for the Martin Bridge Formation (Upper Triassic, Wallowa Terrane, Oregon, U.S.A.). Rev. Paléobiol. 2018, 37, 109–119. [Google Scholar] [CrossRef]
- Heerwagen, E.; Davies, J.H.F.L.; Schmidt, S.T.; Ulianov, A.; Martini, R. The Paleoposition of the Antimonio Depositional System (Sonora, Mexico): New Insights from Nonparametric and Multivariate Analysis of Detrital Zircon Data. J. S. Am. Earth Sci. 2020, 105, 102913. [Google Scholar] [CrossRef]
- Heerwagen, E.; Martini, R. The Vizcaíno Terrane: Another Occurrence of Upper Triassic Shallow-Marine Carbonates in Mexico. Facies 2020, 66, 8. [Google Scholar] [CrossRef]
- Heerwagen, E.; Martini, R. The Antimonio Ramp in Sonora, Mexico: A Shallow-Marine Upper Triassic Mixed Siliciclastic Carbonate Ramp System. Facies 2018, 64, 6. [Google Scholar] [CrossRef]
- Del Piero, N.; Rigaud, S.; Takahashi, S.; Poulton, S.W.; Martini, R. Unravelling the Paleoecology of Flat Clams: New Insights from an Upper Triassic Halobiid Bivalve. Glob. Planet. Chang. 2020, 190, 103195. [Google Scholar] [CrossRef]
- Del Piero, N.; Rigaud, S.; Martini, R. Taanella Yukonensis n. Gen. n. Sp., an Atypically Porous Ataxophragmiidae Foraminifera from the Upper Triassic of Panthalassa. J. Foraminifer. Res. 2022, 52, 99–107. [Google Scholar]
- Forel, M.B.; Del Piero, N.; Rigaud, S.; Martini, R. Ostracods from the Late Triassic (Norian) of Yukon, Canada: New Taxonomic and Palaeogeographic Insights. Riv. Ital. Paleontol. Stratigr. 2022, 128, 325–328. [Google Scholar] [CrossRef]
- Bucur, I.; Rigaud, S.; Del Piero, N.; Fucelli, A.; Heerwagen, E.; Peybernes, C.; Peyrotty, G.; Verard, C.; Chablais, J.; Martini, R. Upper Triassic Calcareous Algae from the Panthalassa Ocean. Riv. Ital. Paleontol. Stratigr. 2020, 126, 499–540. [Google Scholar] [CrossRef]
- Tempelman-kluit, D.J. Reconnaissance Geology, Laberge Map-Area; Geological Survey of Canada: Ottawa, ON, Canada, 1978; pp. 61–66. [Google Scholar]
- England, T.D.J. A Study of Upper Triassic Conodonts of the Intermontane Belt, Yukon Territory. Bachelor’s Thesis, Department of Geological Sciences, The University of British Columbia, Vancouver, BC, Canada, 1980. Unpublished. [Google Scholar]
- Reid, R.P. The Facies and Evolution of an Upper Triassic Reef Complex in Northern Canada. Ph.D. Thesis, University of Miami, Coral Gables, FL, USA, 1985. [Google Scholar]
- Reid, R.P.; Tempelman-Kluit, D.J. Upper Triassic Tethyan-Type Reefs in the Yukon. Bull. Can. Pet. Geol. 1987, 35, 316–332. [Google Scholar]
- Yarnell, J.M.; Stanley, G.D.J.; Hart, C.J.R. New Paleontological Investigations of Upper Triassic Shallow-Water Reef Carbonates (Lewes River Group) in the Whitehorse Area, Yukon. In Yukon Exploration and Geology 1998; Roots, J., Edmond, D.S., Eds.; Exploration and Geological Services Division: Yukon, Indian; Northern Affairs Canada: Gatineau, QC, Canada, 1999; pp. 179–184. [Google Scholar]
- Yarnell, J.M. Paleontology of Two North American Triassic Reef Faunas: Implications for Terrane Paleontology. Master’s Thesis, University of Montana, Missoula, MT, USA, 2000. [Google Scholar]
- Reid, R.P.; Ginsburg, R.N. The Role of Framework in Upper Triassic Patch Reefs in the Yukon Canada. Palaios 1986, 1, 590–600. [Google Scholar] [CrossRef]
- Reid, R.P. Discovery of Triassic Phylloid Algae: Possible Links with the Paleozoic. Can. J. Earth Sci. 1986, 23, 2068–2071. [Google Scholar] [CrossRef]
- Reid, R.P. Nonskeletal Peloidal Precipitates in Upper Triassic Reefs, Yukon Territory (Canada). J. Sediment. Res. 1987, 57, 893–900. [Google Scholar] [CrossRef]
- Reid, R.P. Lime Peak Reef Complex, Norian Age, Yukon. Canada Adjac. Areas 1989, 13, 758–765. [Google Scholar]
- Senowbari-Daryan, B.; Reid, R.P. Upper Triassic Sponges (Sphinctozoa) from Southern Yukon, Stikinia Terrane. Can. J. Earth Sci. 1987, 24, 882–902. [Google Scholar] [CrossRef]
- Gaździcki, A.; Reid, R.P. Upper Triassic Involutinidae (Foraminifera) of Lime Peak in Yukon, Canada. Acta Geol. Pol. 1983, 33, 99–106. [Google Scholar]
- Hart, C.J.R. A Transect across Northern Stikinia: Geology of the Northern Whitehorse Map Area, Southern Yukon Territory (105D/13-16); Indigenous and Northern Affairs Canada: Yellowknife, NT, Canada, 1997. [Google Scholar]
- Coney, P.J.; Jones, D.L.; Monger, J.W.H. Cordilleran Suspect Terranes. Nature 1980, 288, 329–333. [Google Scholar] [CrossRef]
- Jones, D.L.; Silberling, N.J.; Hillhouse, J. Wrangellia—A Displaced Terrane in Northwestern North America. Can. J. Earth Sci. 1977, 14, 2565–2577. [Google Scholar] [CrossRef]
- Colpron, M.; Nelson, J.L.; Murphy, D.C. Northern Cordilleran Terranes and Their Interactions through Time. GSA Today 2007, 17, 4–10. [Google Scholar] [CrossRef]
- Wheeler, J.O.; McFeeely, P. Tectonic Assemblage Map of the Canadian Cordillera and Adjacent Part of the United States of America; Geological Survey of Canada: Ottawa, ON, Canada, 1991. [Google Scholar]
- Belasky, P.; Stevens, C.H.; Hanger, R.A. Early Permian Location of Western North American Terranes Based on Brachiopod, Fusulinid, and Coral Biogeography. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 179, 245–266. [Google Scholar] [CrossRef]
- Beranek, L.P.; Mortensen, J.K. The Timing and Provenance Record of the Late Permian Klondike Orogeny in Northwestern Canada and Arc-Continent Collision along Western North America. Tectonics 2011, 30, TC5017. [Google Scholar] [CrossRef] [Green Version]
- Aberhan, M. Terrane History of the Canadian Cordillera: Estimating Amounts of Latitudinal Displacement and Rotation of Wrangellia and Stikinia. Geol. Mag. 1999, 136, 481–492. [Google Scholar] [CrossRef]
- Smith, P.L. Paleobiogeography and Early Jurassic Molluscs in the Context of Terrane Displacement in Western Canada. In Paleogeography of the North American Cordillera: Evidence for and against Large-Scale Displacements; Haggart, J.W., Enkin, R.J., Monger, J.W.H., Eds.; Geological Association of Canada: St. John’s, NL, Canada, 2006; pp. 81–91. [Google Scholar]
- Golding, M.L. Evaluating Tectonic Models for the Formation of the North American Cordillera Using Multivariate Statistical Analysis of Late Triassic Conodont Faunas. Paleobiodivers. Paleoenviron. 2019, 100, 135–149. [Google Scholar] [CrossRef]
- Golding, M.L.; Mortensen, J.K.; Zonneveld, J.P.; Orchard, M.J. U-Pb Isotopic Ages of Euhedral Zircons in the Rhaetian of British Columbia: Implications for Cordilleran Tectonics during the Late Triassic. Geosphere 2016, 12, 1606–1616. [Google Scholar] [CrossRef]
- Golding, M.L.; Mortensen, J.K.; Ferri, F.; Zonneveld, J.P.; Orchard, M.J. Determining the Provenance of Triassic Sedimentary Rocks in Northeastern British Columbia and Western Alberta Using Detrital Zircon Geochronology, with Implications for Regional Tectonics. Can. J. Earth Sci. 2016, 53, 140–155. [Google Scholar] [CrossRef]
- Schroeder-Adams, C.; Haggart, J.W. Biogeography of Foraminifera in Tectonic Reconstructions; Imitations and Constraints on the Paleogeographic Position of Wrangellia. Geol. Assoc. Canada Spec. Pap. 2006, 46, 95–108. [Google Scholar]
- Colpron, M.; Nelson, J.L. A Digital Atlas of Terranes for the Northern Cordillera; BCGS GeoFile; British Columbia Ministry of Energy and Mines: Victoria, BC, Canada, 2011. [Google Scholar]
- Struik, L.C.; Schiarizza, P.; Orchard, M.J.; Cordey, F.; Sano, H.; MacIntyre, D.G.; Lapierre, H.; Tardy, M. Imbricate Architecture of the Upper Paleozoic to Jurassic Oceanic Cache Creek Terrane, Central British Columbia. Can. J. Earth Sci. 2001, 38, 495–514. [Google Scholar] [CrossRef]
- Mihalynuk, M.G.; Nelson, J.A.; Diakow, L.J. Cache Creek Terrane Entrapment: Oroclinal Paradox within the Canadian Cordillera. Tectonics 1994, 13, 575–595. [Google Scholar] [CrossRef]
- Bordet, E. Bedrock Geology Map of the Teslin Mountain and East Lake Laberge Areas; Parts of NTS 105E/2, 105E/3 and 105E/6; Yukon Geological Survey: Whitehorse, YT, Canada, 2016. [Google Scholar]
- Bordet, E. Updates on the Middle Triassic-Middle Jurassic Stratigraphy and Structure of the Teslin Mountain and East Lake Laberge Areas, South-Central Yukon. In Yukon Exploration and Geology 2016; MacFarlane, K.E., Weston, L.H., Eds.; Yukon Geological Survey: Whitehorse, YT, Canada, 2017; pp. 1–24. [Google Scholar]
- Tempelman-Kluit, D.J. Geology, Laberge (105E) and Carmacks (115I); Yukon Geological Survey: Whitehorse, YT, Canada, 1984. [Google Scholar]
- Monger, J.W.H.; Wheeler, J.O.; Tipper, H.W.; Gabrielse, H.; Harms, T.; Struik, L.C.; Campbell, R.B.; Dodds, C.J.; Gehrels, G.E.; O’Brien, J. Part B. Cordilleran Terranes, Upper Devonian to Middle Jurassic Assemblages. In Geology of the Cordilleran Orogen in Canada; Gabrielse, H., Yorath, C.J., Eds.; Geological Survey of Canada: Ottawa, ON, Canada, 1991; pp. 281–327. [Google Scholar]
- Simmons, A.T.; Tosdal, R.M.; Awmack, H.J.; Wooden, J.L.; Friedman, R.M. Early Triassic Stuhini Group and Tertiary Sloko Group Magmatism (NTS 104K/10W), Northwestern British Columbia: New U-Pb Geochronological Results; British Columbia Geological Survey: Victoria, BC, Canada, 2007. [Google Scholar]
- Bordet, E.; Crowley, J.L.; Piercey, S.J. Geology of the Eastern Lake Laberge Area (105E), South-Central Yukon; Yukon Geological Survey: Whitehorse, YT, Canada, 2019. [Google Scholar]
- White, D.; Colpron, M.; Buffett, G. Seismic and Geological Constraints on the Structure and Hydrocarbon Potential of the Northern Whitehorse Trough, Yukon, Canada. Bull. Can. Pet. Geol. 2012, 60, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Bordet, E. Preliminary Results on the Middle Triassic-Middle Jurassic Stratigraphy and Structure of the Teslin Mountain Area, Southern Yukon. In Yukon Exploration and Geology 2015; MacFarlane, K.E., Nordling, M.G., Eds.; Yukon Geological Survey: Whitehorse, YT, Canada, 2016; pp. 43–61. [Google Scholar]
- Morrison, G.W. Setting and Origin of Skarn Deposits in the Whitehorse Copper Belt, Yukom. Ph.D. Thesis, The University of Western Ontario, London, ON, Canada, 1981. [Google Scholar]
- Lokier, S.W.; Al Junaibi, M. The Petrographic Description of Carbonate Facies: Are We All Speaking the Same Language? Sedimentology 2016, 63, 1843–1885. [Google Scholar] [CrossRef] [Green Version]
- Burne, R.V.; Moore, L.S. Microbialites: Organosedimentary Deposits of Benthic Microbial Communites. Palaios 1987, 2, 241–254. [Google Scholar] [CrossRef]
- Riding, R. Structure and Composition of Organic Reefs and Carbonate Mud Mounds: Concepts and Categories. Earth-Sci. Rev. 2002, 58, 163–231. [Google Scholar] [CrossRef]
- Turnšek, D.; Ramovš, A. Upper Triassic (Norian-Rhaetian) Reef Buildings in the Northern Julian Alps (NW Yugoslavia); Slovenian Academy of Sciences and Arts: Ljubljana, Slovenia, 1987. [Google Scholar]
- Crasquin-Soleau, S.; Vaslet, D.; Le Nindre, Y.-M. Ostracods As Markers of the Permian ⁄ Triassic Boundary in the Khuff Formation of Saudi Arabia. Paleontology 2005, 48, 853–868. [Google Scholar] [CrossRef]
- Lei, J.Z.X.; Golding, M.L.; Hussin, J.M. Paleoenvironmental Interpretation and Identification of the Norian-Rhaetian Boundary in the Whitehorse Trough (Stikine Terrane, Northern Canadian Cordillera). Geol. Soc. Am. Programs 2019, 51, 12–14. [Google Scholar] [CrossRef]
- Golding, M.L.; Orchard, M.J.; Zagorevski, A. Conodonts from the Stikine Terrane in Northern British Columbia and Southern Yukon; Open File; Geological Survey of Canada: Ottawa, ON, Canada, 2017. [Google Scholar]
- Reid, R.P.; Macintyre, I.G.; James, N.P. Internal Precipitation of Microcrystalline Carbonate: A Fundamental Problem for Sedimentologists. Sediment. Geol. 1990, 68, 163–170. [Google Scholar] [CrossRef]
- Flügel, E. Microfacies of Carbonate Rocks. Analysis, Interpretation and Application, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 9788578110796. [Google Scholar]
- Keim, L.; Schlager, W. Automicrite Facies on Steep Slopes (Triassic, Dolomites, Italy). Facies 1999, 41, 15–25. [Google Scholar] [CrossRef]
- Wolf, K.H. “Grain-Diminution” of Algal Colonies to Micrite. J. Sediment. Petrol. 1965, 35, 420–427. [Google Scholar] [CrossRef]
- Russo, F.; Neri, C.; Mastandrea, A.; Baracca, A. The Mud Mound Nature of the Cassian Platform Margins of the Dolomites. A Case History: The Cipit Boulders from Punta Grohmann (Sasso Piatto Massif, Northern Italy). Facies 1997, 36, 25–36. [Google Scholar] [CrossRef]
- McRoberts, C.A. Late Triassic Bivalvia (Chiefly Halobiidae and Monotidae) from the Pardonet Formation, Williston Lake Area, Northeastern British Columbia, Canada. J. Paleontol. 2011, 85, 613–664. [Google Scholar] [CrossRef]
- Stanley, G.D.J.; Swart, P.K. Evolution of the Coral-Zooxanthellae Symbiosis during the Triassic: A Geochemical Approach. Paleobiology 1995, 21, 179–199. [Google Scholar] [CrossRef]
- Muscatine, L.; Goiran, C.; Land, L.; Jaubert, J.; Cuif, J.P.; Allemand, D. Stable Isotopes (Δ13C and Δ15N) of Organic Matrix from Coral Skeleton. Proc. Natl. Acad. Sci. USA 2005, 102, 1525–1530. [Google Scholar] [CrossRef] [Green Version]
- Stanley, G.D.J.; Helme, K.P. Middle Triassic Coral Growth Bands and Their Implication for Photosymbiosis. Palaios 2010, 25, 754–763. [Google Scholar] [CrossRef]
- Kiessling, W. Reef Expansion during the Triassic: Spread of Photosymbiosis Balancing Climatic Cooling. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 290, 11–19. [Google Scholar] [CrossRef]
- Flügel, E. Diversity and Environments of Permian and Triassic Dasycladacean Algae. In Paleoalgology: Contemporary Research and Applications; Toomey, D.F., Nitecki, M.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 344–351. [Google Scholar]
- Valet, G. Paleoecological Approach to Dasycladales from the Ecology of Recent Forms. Bull. Cent. Rech. Explor. Elf-Aquitaine 1979, 3, 859–866. [Google Scholar]
- Reitner, J. Modern Cryptic Microbialite/Metazoan Facies from Lizard Island (Great Barrier Reef, Australia) Formation and Concepts. Facies 1993, 29, 3–40. [Google Scholar] [CrossRef] [Green Version]
- Reolid, M.; Gaillard, C.; Olóriz, F.; Rodríguez-Tovar, F.J. Microbial Encrustations from the Middle Oxfordian-Earliest Kimmeridgian Lithofacies in the Prebetic Zone (Betic Cordillera, Southern Spain): Characterization, Distribution and Controlling Factors. Facies 2005, 50, 529–543. [Google Scholar] [CrossRef]
- Gaillard, C. Les Biohermes à Spongiaires et Leur Environment Dans l’Oxfordien Du Jura Méridional. Trav. Doc. Lab. Géol. Lyon 1983, 90, 3–515. [Google Scholar]
- Gaillard, C. Bioconstructions Jurassiques. Mém. Bur. Rech. Géol. Min. Fr. 1984, 125, 276–281. [Google Scholar]
- Rodríguez-Martínez, M.; Heim, C.; Simon, K.; Zilla, T.; Reitner, J. Tolypammina Gregaria Wendt 1969-Frutexites Assemblage and Ferromanganese Crusts: A Coupled Nutrient-Metal Interplay in the Carnian Sedimentary Condensed Record of Hallstatt Facies (Austria). In Advances in Stromatolite Geobiology; Reitner, J., Quéric, N.-V., Arp, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 409–434. ISBN 978-3-642-10415-2. [Google Scholar]
- Keim, L.; Schlager, W. Quantitative Compositional Analysis of a Triassic Carbonate Platform (Southern Alps. Italy). Sediment. Geol. 2001, 139, 261–283. [Google Scholar] [CrossRef]
- Tosti, F.; Mastandrea, A.; Guido, A.; Demasi, F.; Russo, F.; Riding, R. Biogeochemical and Redox Record of Mid-Late Triassic Reef Evolution in the Italian Dolomites. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 399, 52–66. [Google Scholar] [CrossRef]
- Tinker, S.W. Shelf-to-Basin Facies Distributions and Sequence Stratigraphy of a Steep-Rimmed Carbonate Margin: Capitan Depositional System, Mckittrick Canyon, New Mexico and Texas. J. Sediment. Res. 1998, 68, 1146–1174. [Google Scholar] [CrossRef]
- Kenter, J.A.M.; Harris, P.M.; Della Porta, G. Steep Microbial Boundstone-Dominated Platform Margins—Examples and Implications. Sediment. Geol. 2005, 178, 5–30. [Google Scholar] [CrossRef]
- Reid, R.P.; Macintyre, I.G. Carbonate Recrystallization in Shallow Marine Environments: A Widespread Diagenetic Process Forming Micritized Grains. J. Sediment. Res. 1998, 68, 928–946. [Google Scholar] [CrossRef]
- Reid, R.P.; Macintyre, I.G. Microboring versus Recrystallization: Further Insight into the Micritization Process. J. Sediment. Res. 2000, 70, 24–28. [Google Scholar] [CrossRef]
- Kendall, C.G.S.C.; Alsharhan, A.S. Coastal Holocene Carbonates of Abu Dhabi, UAE: Depositional Setting, Geomorphology, and Role of Cyanobacteria in Micritization. Int. Assoc. Sedimentol. Spec. Publ. 2011, 43, 205–220. [Google Scholar]
- Bathurst, R.G.C. Boring Algae, Micritic Envelopes and Lithification of Molluscan Biosparites. Geol. J. 1966, 5, 15–32. [Google Scholar] [CrossRef]
- Davis, R.A.J.; Dalrymple, R.W. (Eds.) Principles of Tidal Sedimentology; Spinger: Berlin/Heidelberg, Germany, 2012; ISBN 9788578110796. [Google Scholar]
- Shinn, E.A.; Michael, L.; Ginsburg, R.N. Anatomy of a Modern Carbonate Tidal-Flat, Andros Island, Bahamas. J. Sediment. Petrol. 1969, 39, 1202–1228. [Google Scholar]
- Gischler, E. Sedimentary Facies of Bora Bora, Darwin’s Type Barrier Reef (Society Islands, South Pacific): The Unexpected Occurrence of Non-Skeletal Grains. J. Sediment. Res. 2011, 81, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Hunter, A.W.; Underwood, C.J. Palaeoenvironmental Control on Distribution of Crinoids in the Bathonian (Middle Jurassic) of England and France. Acta Palaeontol. Pol. 2009, 54, 77–98. [Google Scholar] [CrossRef] [Green Version]
- Mulder, T.; Alexander, J. The Physical Character of Subaqueous Sedimentary Density Flow and Their Deposits. Sedimentology 2001, 48, 269–299. [Google Scholar] [CrossRef]
- Middleton, G.V.; Southard, J.B. Mechanics of Sediment Transport. Eastern Section Short Course 3, 2nd ed.; SEPM Society for Sedimentary Geology: Tulsa, OK, USA, 1984. [Google Scholar]
- Mohrig, D.; Elverhoi, A.; Parker, G. Experiments on the Relative Mobility of Muddy Subaqueous and Subaerial Debris flows, and Their Capacity to Remobilize Antecedent Deposits. Mar. Geol. 1999, 154, 117–129. [Google Scholar] [CrossRef]
- Mohrig, D.; Whipple, K.X.; Hondzo, M.; Ellis, C.; PArker, G. Hydroplaning of Subaqueous Debris Flows. Geol. Soc. Am. Bull. 1998, 110, 387–394. [Google Scholar] [CrossRef]
- Iverson, R.M. Physics of Debris Flows. Rev. Geophys. 1997, 35, 245–296. [Google Scholar] [CrossRef] [Green Version]
- Hampton, M.A. Competence of Fine-Grained Debris Flows. J. Sediment. Petrol. 1975, 45, 834–844. [Google Scholar] [CrossRef]
- Hampton, M.A. The role of subaqueous debris flow in generating turbidity currents. J. Sediment. Petrol. 1972, 42, 775–793. [Google Scholar]
- Rasser, M.W.; Riegl, B. Holocene Coral Reef Rubble and Its Binding Agents. Coral Reefs 2002, 21, 57–72. [Google Scholar] [CrossRef]
- Beltrán, Y.; Cerqueda-García, D.; Taş, N.; Thomé, P.E.; Iglesias-Prieto, R.; Falcón, L.I. Microbial Composition of Biofilms Associated with Lithifying Rubble of Acropora Palmata Branches. FEMS Microbiol. Ecol. 2016, 92, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camoin, G.F.; Gautret, P.; Montaggioni, L.F.; Cabioch, G. Nature and Environmental Significance of Microbialites in Quaternary Reefs: The Tahiti Paradox. Sediment. Geol. 1999, 126, 271–304. [Google Scholar] [CrossRef]
- Piller, W. Involutinacea (Foraminifera) Der Trias Und Des Lias. Beiträge Paläontol. Osterr. 1978, 5, 1–164. [Google Scholar]
- Sarg, J.F. Carbonate Sequence Stratigraphy. In Sea-Level Changes: An Integrated Approach; Wilgus, C.K., Hastings, B.S., Posamentier, H., Van Wagoner, J., Ross, C.A., Kendall, C.G.S.C., Eds.; Society for Sedimentary Geology: Tulsa, OK, USA, 1988; pp. 155–182. [Google Scholar]
- Schmid, D.U. Marine Mikrobolithe Und Mikroinkrustierer Aus Dem Oberjura. Profil 1996, 9, 101–251. [Google Scholar]
- Watkins, R. Volcaniclastic and Carbonate Sedimentation in Late Paleozoic Island-Arc Deposits, Eastern Klamath Mountains, California. Geology 1985, 13, 709–713. [Google Scholar] [CrossRef]
- Watkins, R. Carboniferous and Permian Island-Arc Deposits of the Eastern Klamath Terrane, California. In Paleozoic and Early Mesozoic Paleogeographic Relations: Sierra Nevada, Klamath Mountains, And Related Terranes; Harwood, D.S., Meghan Miller, M., Eds.; Geological Society of America: Boulder, CO, USA, 1990; pp. 193–200. [Google Scholar]
- Watkins, R. Carbonate Bank Sedimentation in a Volcaniclastic Arc Setting; Lower Carboniferous Limestones of the Eastern Klamath Terrane, California. Geology 1993, 63, 966–973. [Google Scholar]
- Watkins, R.; Wilson, E.C. Paleoecologic and Biogeographic Significance of the Biostromal Organism Palaeoaplysina in the Lower Permian McCloud Limestone, Eastern Klamath Mountains, California. Palaios 1989, 4, 181–192. [Google Scholar] [CrossRef]
- Hathway, B. Sedimentation and Volcanism in an Oligocene-Miocene Intra-Oceanic Arc and Fore-Arc, Southwestern Viti Levu, Fiji. J. Geol. Soc. Lond. 1994, 151, 499–514. [Google Scholar] [CrossRef]
- Utami, D.A.; Reuning, L.; Cahyarini, S.Y. Satellite- and Field-Based Facies Mapping of Isolated Carbonate Platforms from the Kepulauan Seribu Complex, Indonesia. Depos. Rec. 2018, 4, 255–273. [Google Scholar] [CrossRef]
- Anderson, O. Protozoa, Radiolarians. In Encyclopedia of Ocean Sciences; Steve, A.T., Turekian, K.K., Eds.; Academic Press: Oxford, UK, 2001; pp. 613–617. ISBN 978-0-12-374473-9. [Google Scholar]
- Soja, C.M. Island-Arc Carbonates: Characterization and Recognition in the Ancient Geologic Record. Earth-Sci. Rev. 1996, 41, 31–65. [Google Scholar] [CrossRef]
- Dorobek, S.L. Carbonate-Platform Facies in Volcanic-Arc Settings: Characteristics and Controls on Deposition and Stratigraphic Development. Spec. Pap. Geol. Soc. Am. 2008, 436, 55–90. [Google Scholar] [CrossRef]
- Meltzner, A.J.; Sieh, K.; Chiang, H.W.; Wu, C.C.; Tsang, L.L.H.; Shen, C.C.; Hill, E.M.; Suwargadi, B.W.; Natawidjaja, D.H.; Philibosian, B.; et al. Time-Varying Interseismic Strain Rates and Similar Seismic Ruptures on the Nias-Simeulue Patch of the Sunda Megathrust. Quat. Sci. Rev. 2015, 122, 258–281. [Google Scholar] [CrossRef]
- López, G. Walther’s Law of Facies. In Encyclopedia of Scientific Dating Methods; Rink, W.J., Thompson, J.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 957–958. Available online: http://link.springer.com/referenceworkentry/10.1007/978-94-007-6304-3_30 (accessed on 26 January 2022).
- Frisia-Bruni, S.; Jadoul, F.; Weissert, H. Evinosponges in the Triassic Esino Limestone (Southern Alps): Documentation of Early Lithification and Late Diagenetic Overprint. Sedimentology 1989, 36, 685–699. [Google Scholar] [CrossRef]
- Russo, F.; Gautret, P.; Mastandrea, A.; Perri, E. Syndepositional Cements Associated with Nannofossils in the Marmolada Massif: Evidences of Microbially Mediated Primary Marine Cements? (Middle Triassic, Dolomites, Italy). Sediment. Geol. 2006, 185, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Emmerich, A.; Zamparelli, V.; Bechstädt, T.; Zühlke, R. The Reefal Margin and Slope of a Middle Triassic Carbonate Platform: The Latemar (Dolomites, Italy). Facies 2005, 50, 573–614. [Google Scholar] [CrossRef]
- Stoppani, A. Les Petrifications d’Esino Ou Description Des Fossiles Appartenants Au Depot Superieur Des Environs d’Esino En Lombardie. Paleontol. Lomb. 1858, 1, 360. [Google Scholar]
- German, K. Calcite and Dolomite Fibrous Cements (Grossoolith) in the Reef Rocks of the Wettersteinkalk (Ladinian, Middle Trias), Northern Limestone Alps, Bavaria and Tyrol. In Carbonate Cements; Bricker, O.P., Ed.; Johns Hopkins Press: Baltimore, MA, USA, 1971; pp. 185–188. [Google Scholar]
- McKenzie, J.A.; Lister, G. Origin of Alternating Calcite and Dolomite Void Filling Cements (Grossoolites)in Middle Triassic Reefs of the Northern Limestone Alps near Innsbruck, Austria. In I.A.S. 4th European Regional Meeting Abstracts Book; International Association of Sedimentologists: Split, Yugoslavia, 1983; pp. 156–157. [Google Scholar]
- Cosjin, J. De Geologie van the Valle Di Olmo Al Brembo. Leid. Geol. Med. 1928, 2, 251–324. [Google Scholar]
- Hofsteenge, L. La Geologie de La Valee Du Brembo et de Ses Affluents Entre Lenna et San Pellegrino. Leid. Geol. Med. 1932, 4, 25–82. [Google Scholar]
- Russo, F.; Mastandrea, A.; Stefani, M.; Neri, C. Carbonate Facies Dominated by Syndepositional Cements: A Key Component of Middle Triassic Platforms. The Marmolada Case History (Dolomites, Italy). Facies 2000, 42, 211–226. [Google Scholar] [CrossRef]
- Gale, L.; Rigaud, S.; Gennari, V.; Blau, J.; Rettori, R.; Martini, R.; Gaetani, M. Recognition of Upper Triassic Temperate Foraminiferal Assemblages: Insights from the Khodz Group (NW Caucasus, Russia). Glob. Planet. Chang. 2020, 188, 103152. [Google Scholar] [CrossRef]
- Opdyke, B.N.; Wilkinson, B.H. Paleolatitude Distribution of Phanerozoic Marine Ooids and Cements. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 78, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Manzello, D.P.; Kleypas, J.A.; Budd, D.A.; Eakin, C.M.; Glynn, P.W.; Langdon, C. Poorly Cemented Coral Reefs of the Eastern Tropical Pacific: Possible Insights into Reef Development in a High-CO2 World. Proc. Natl. Acad. Sci. USA 2008, 105, 10450–10455. [Google Scholar] [CrossRef] [Green Version]
- Stoner, A.W.; Waite, J.M. Distribution and Behavior of Queen Conch Strombus Gigas Relative to Seagrass Standing Crop. Fish. Bull. US 1990, 88, 451–460. [Google Scholar]
- Stephenson, S.P.; Sheridan, N.E.; Geiger, S.P.; Arnold, W.S. Abundance and Distribution of Large Marine Gastropods in Nearshore Seagrass Beds along the Gulf Coast of Florida. J. Shellfish Res. 2013, 32, 305–313. [Google Scholar] [CrossRef]
- Yancey, T.E.; Stanley, G.D.J. Giant Alatoform Bivalves in the Upper Triassic of Western North America. Palaeontology 1999, 42, 1–23. [Google Scholar] [CrossRef]
- Stanley, G.D.J.; Yancey, T.E.; Shepherd, H.M.E. Giant Upper Triassic Bivalves of Wrangellia, Vancouver Island, Canada. Can. J. Earth Sci. 2013, 50, 142–147. [Google Scholar] [CrossRef]
- Smith, S.V.; Kimmerer, W.J.; Laws, E.A.; Brock, R.E.; Walsh, T.W. Kaneohe Bay Sewage Diversion Experiment: Perspectives on Ecosystem Responses to Nutritional Perturbation. Pacific Sci. 1981, 35, 279–395. [Google Scholar]
- Hallock, P.; Schlager, W. Nutrient Excess and the Demise of Coral Reefs and Carbonate Platforms. Palaios 1986, 1, 389–398. [Google Scholar] [CrossRef]
- Hallock, P. The Role of Nutrient Availability in Bioerosion: Consequences to Carbonate Buildups. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 63, 275–291. [Google Scholar] [CrossRef]
- Hallock, P. Coral Reefs, Carbonate Sedimentation, Nutrients, and Global Change. In The History and Sedimentology of Ancient Reef Ecosystems; Stanley, G.D.J., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2001; pp. 387–427. [Google Scholar]
- Mutti, M.; Hallock, P. Carbonate Systems along Nutrient and Temperature Gradients: Some Sedimentological and Geochemical Constraints. Int. J. Earth Sci. 2003, 92, 465–475. [Google Scholar] [CrossRef]
- Camoin, G.F.; Cabioch, G.; Eisenhauer, A.; Braga, J.C.; Hamelin, B.; Lericolais, G. Environmental Significance of Microbialites in Reef Environments during the Last Deglaciation. Sediment. Geol. 2006, 185, 277–295. [Google Scholar] [CrossRef] [Green Version]
- Elliott, G.F. Ecologic Significance of Post-Palaeozoic Green Calcareous Algae. Geol. Mag. 1978, 115, 437–442. [Google Scholar] [CrossRef]
- Ott, E. Die Gesteinsbildenden Kalkalgen Im Schlauchkar (Karwendelgebirge). Jb. Ver. Schutze Alpenpfl. 1966, 31, 1–8. [Google Scholar]
- Ott, E. Dasycladaceen (Kalkalgen) Aus Der Nordalpinen Obertrias. Mitt. Bayer. Staatssamml. Palaont. Hist. Geol. 1967, 7, 205–226. [Google Scholar]
- Ott, E. Mitteltriadischen Riffe Der Nördlichen Kalkalpen Und Altersgleiche Bildungen Auf Karaburun Und Chios (Ägäis). Mitt. Ges. Geol. Bergbaustud. 1972, 21, 251–276. [Google Scholar]
- Di Stefano, P.; Senowbari-Daryan, B. Upper Triaissic Dasycladales (Green Algae) from the Palermo Mountains (Sicily, Italy). Geol. Rom. 1985, 24, 189–220. [Google Scholar]
- Bodeur, Y. Le Complexe Récifal Jurassique Supérieur Au Sud Des Cévennes: Architecture Sédimentologique. C. R. Acad. Sc. 1976, 282, 835–837. [Google Scholar]
- Rigaud, S. The Late Triassic Martin Bridge Carbonate Platform (Wallowa Terrane, NW U.S.A.): Sedimentology, Biostratigraphy, and Contribution to the Understanding of Aragonitic and Microgranular Foraminifers. Ph.D. Thesis, University of Geneva, Geneva, Switzerland, 2012. [Google Scholar]
- Zorn, H. Über Den Lebensraum Fossiler Wirtelalgen in Der Trias Der Alpen. Naturwissenschaften 1976, 63, 426–429. [Google Scholar] [CrossRef]
- Zorn, H. Del Einfluss Der Umwelt Auf Die Variation Fossiler Arten. Nat. Mus. 1977, 107, 1–5. [Google Scholar]
- Payne, J.L. Evolutionary Dynamics of Gastropod Size across the End-Permian Extinction and through the Triassic Recovery Interval. Paleobiology 2005, 31, 269–290. [Google Scholar] [CrossRef]
- Ketwetsuriya, C.; Nose, M.; Charoentitirat, T.; Nützel, A. Microbial-, Fusulinid Limestones with Large Gastropods and Calcareous Algae: An Unusual Facies from the Early Permian Khao Khad Formation of Central Thailand. Facies 2020, 66, 21. [Google Scholar] [CrossRef]
- Stanton, R.J.; Flügel, E. Problems with Reef Models: The Late Triassic Steinplatte “ Reef “ (Northern Alps, Salzburg/Tyrol, Austria). Facies 1989, 33, 1. [Google Scholar] [CrossRef]
- Connolly, M.W.; Lambert, L.L.; Stanton, R.J. Paleoecology of Lower and Middle Pennsylvanian (Middle Carboniferous) Chaetetes in North America. Facies 1989, 20, 139–168. [Google Scholar] [CrossRef]
- Heindel, K.; Birgel, D.; Peckmann, J.; Kuhnert, H.; Westphal, H. Formation of Deglacial Microbialites in Coral Reefs off Tahiti (IODP 310) Involving Sulfate-Reducing Bacteria. Palaios 2010, 25, 618–635. [Google Scholar] [CrossRef]
- Sánchez-Beristain, F.; Reitner, J. Paleoecology of Microencrusters and Encrusting “Coralline” Sponges in Cipit Boulders from the Cassian Formation (Upper Ladinian-Lower Carnian, Dolomites, Northern Italy). Palaontol. Z. 2012, 86, 113–133. [Google Scholar] [CrossRef]
- Monty, C.L. Cavity or Fissure Dwelling Stromatolites (Endostromatolites) from Belgian Devonian Mud Mounds (Exteded Abstracts). Ann. Soc Géol. Belg. 1982, 105, 343–344. [Google Scholar]
- Granier, B. The Biosignature of Sparite Permits the Distinction between Gravitational Cement and Endostromatolites. Carnets Geol. 2020, 20, 407–419. [Google Scholar] [CrossRef]
- Schopf, W.J. The Fossil Record of Cyanobacteria. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 15–36. [Google Scholar]
- Hohenegger, J.; Piller, W. Ökologie Und Systematische Stellung Der Foraminiferen Im Gebankten Dachsteinkalk (Obertrias) Des Nördlichen Toten Gebirges (Oberösterreich). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1975, 18, 241–276. [Google Scholar] [CrossRef]
- Zaninetti, L.; Ciarapica, G.; Cirilli, S.; Cadet, J.-P. Miliolechina Stellata, n. Gen., n. Sp., et Hirsutospirella Pilosa, n. Gen., n. Sp. (Foraminifères), Dans Le Trias Supérieur (Norien) à Faciès Récifal Des Dinarides. Rev. Paléobiol. 1985, 4, 331–341. [Google Scholar]
- Martini, R.; Zaninetti, L.; Ciarapica, G. Hirsutospirella Pilosa Zaninetti, Ciarapica, Cirilli et Cadet, 1985. (Foraminifère, Trias Supérieur), Morphologie et Paléobiologie. Rev. Paléobiol. 1986, 5, 193–196. [Google Scholar]
- Márquez, L. Foraminiferal Fauna Recovered after the Late Permian Extinctions in Iberia and the Westernmost Tethys Area. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 229, 137–157. [Google Scholar] [CrossRef]
- Martini, R.; Zaninetti, L.; Lathuilière, B.; Cirilli, S.; Cornée, J.J.; Villeneuve, M. Upper Triassic Carbonate Deposits of Seram (Indonesia): Palaeogeographic and Geodynamic Implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 206, 75–102. [Google Scholar] [CrossRef]
- Jones, R.W. Foraminifera and Their Applications; Cambridge University Press: New York, NY, USA, 2014; ISBN 9781139567619. [Google Scholar]
- Martindale, R.C.; Corsetti, F.A.; James, N.P.; Bottjer, D.J. Paleogeographic Trends in Late Triassic Reef Ecology from Northeastern Panthalassa. Earth-Sci. Rev. 2015, 142, 18–37. [Google Scholar] [CrossRef]
- Sun, Y.; Orchard, M.J.; Kocsis, T.; Joachimski, M.M. Carnian–Norian (Late Triassic) Climate Change: Evidence from Conodont Oxygen Isotope Thermometry with Implications for Reef Development and Wrangellian Tectonics. Earth Planet. Sci. Lett. 2020, 534, 116082. [Google Scholar] [CrossRef]
- Martindale, R.C.; Zonneveld, J.P.; Bottjer, D.J. Microbial Framework in Upper Triassic (Carnian) Patch Reefs from Williston Lake, British Columbia, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 609–620. [Google Scholar] [CrossRef]
- Martindale, R.C.; Bottjer, D.J.; Corsetti, F.A. Platy Coral Patch Reefs from Eastern Panthalassa (Nevada, USA): Unique Reef Construction in the Late Triassic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 313–314, 41–58. [Google Scholar] [CrossRef]
- Oldow, J.S. Structure and Stratigraphy of the Luning Allochthon and the Kinematics of Allochthon Emplacement, Pilot Mountains, West-Central Nevada. Geol. Soc. Am. Bull. 1981, 92, 888–911. [Google Scholar] [CrossRef]
- Zonneveld, J.P.; Henderson, C.M.; Stanley, G.D.J.; Orchard, M.J.; Gingras, M.K. Oldest Scleractinian Coral Reefs on the North American Craton: Upper Triassic (Carnian), Northeastern British Columbia, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 243, 421–450. [Google Scholar] [CrossRef]
- Vérard, C. Panalesis: Towards Global Synthetic Palaeogeographies Using Integration and Coupling of Manifold Models. Geol. Mag. 2019, 156, 320–330. [Google Scholar] [CrossRef]
- Preto, N.; Kustatscher, E.; Wignall, P.B. Triassic Climates—State of the Art and Perspectives. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 290, 1–10. [Google Scholar] [CrossRef]
- Buser, S.; Ramovš, A.; Turnšek, D. Triassic Reefs in Slovenia. Facies 1982, 6, 15–23. [Google Scholar] [CrossRef]
- Gale, L. Rhaetian Foraminiferal Assemblage from the Dachstein Limestone of Mt. Begunjscica (Kosuta Unit, Eastern Southern Alps). Geologija 2012, 55, 17–44. [Google Scholar] [CrossRef]
- Gale, L.; Kastelic, A.; Rozic, B. Taphonomic Features of Late Triassic Foraminifera from Mount Begunjscica, Karavanke Mountains, Slovenia. Palaios 2013, 28, 771–792. [Google Scholar] [CrossRef]
- Senowbari-Daryan, B. Coralline Schwämme Aus Dem Norisch-Rhätischen Dachstein- Riff Des Gosaukammes (Nördliche Kalkalpen, Österreich). Jahrb. Geol. Bundesanst. 2009, 149, 111–166. [Google Scholar]
- Senowbari-Daryan, B. Solenoporaceen Aus Den Obertriassischen (Nor) Riffkalken Des Taurusgebirges (Antalya-Gebiet, Südtürkei). Palaontol. Z. 2005, 79, 409–427. [Google Scholar] [CrossRef]
- Senowbari-Daryan, B.; Reitner, J.; Neuweiler, F.; Gunkel, F. Upper Triassic Reefs and Reef Communities of Iran. Gottinger Arb. Geol. Paläontol. 1996, 2, 299–304. [Google Scholar]
- Senowbari-Daryan, B.; Seyed-Emami, K.; Aghanabati, A. Some Inozoid Sponges from Upper Triassic (Norian-Rhaetian) Nayband Formation of Central Iran. Riv. Ital. Paleontol. Stratigr. 1997, 103, 293–322. [Google Scholar]
- Senowbari-Daryan, B.; Hamadani, A. Thalamid Sponges from the Upper Triassic (Norian-Rhaetian) Nayband Formation near Wali-Abad, SE Abadeh, Central Iran. Riv. Ital. Paleontol. Stratigr. 1999, 105, 79–100. [Google Scholar]
- Senowbari-Daryan, B. Peronidellen (Schwämme) Aus Der Trias Und Beschreibung von Peronidella Iranica n. Sp. Aus Der Obertrias (Nor-Rhät) Des Iran Und von Österreich. Jahrb. Geol. Bundesanst. 2003, 143, 63–72. [Google Scholar]
- Senowbari-Daryan, B.; Hamadani, A. Obertriadische (Nor) Dasycladaceen Aus Der Nayband-Formation Vom Zentraliran. Rev. Paléobiol. 2000, 19, 97–121. [Google Scholar]
- Senowbari-Daryan, B.; Amirhassankhani, F. Lovcenipora Iranica Nov. Sp., an Unusually Large Chaetetid Sponge from the Upper Triassic (Howz-e Khan Member, Nayband Formation) of Northeast Iran. Zitteliana 2013, 53, 15–22. [Google Scholar]
- Senowbari-Daryan, B.; Rashidi, K.; Amirzadeh, M.; Saberzadeh, B.; Talebi, A. Sponges from the Upper Triassic (Norian–Rhaetian) Nayband Formation, Northeast Iran. Jb. Geol. B.A. Wien 2011, 113A, 347–355. [Google Scholar]
- Senowbari-Daryan, B.; Schäfer, P.; Abate, B. Obertdadische Rifle Und Rifforganismen in Sizilien (Beiträge Zur Paläontologie Und Mikrofazies Obertriadischer Riffe Im Alpin-Mediterranen Raum, 27). Facies 1982, 6, 165–183. [Google Scholar] [CrossRef]
- Senowbari-Daryan, B. Mikroproblematika Aus Den Obertriadischen Riffkalken von Sizilien. Münster. Forsch. Geol. Paläont. 1984, 61, 1–181. [Google Scholar]
- Stanley, G.D.J.; Senowbari-Daryan, B. Upper Triassic, Dachstein-Type, Reef Limestone from the Wallowa Mountains, Oregon: First Reported Occurrence in the United States. Palaios 1986, 1, 172–177. [Google Scholar] [CrossRef]
- Senowbari-Daryan, B. Die Systematische Stellung Der Thalamiden Schwämme Und Ihre Bedeutung in Der Erdgeschichte. Münchner Geowiss. Abh. 1990, A-21, 5–326. [Google Scholar]
- Zaninetti, L. Orthotrinacria, n. Gen.,(Protista: Foraminiferida) from Upper Triassic (Norian) Reefs of Sicily. Rev. Paléobiologie 1985, 4, 297–300. [Google Scholar]
- Di Stefano, P.; Gullo, M.; Senowbari-Daryan, B. The Upper Triassic Reef of Monte Genuardo (Southwestern Sicily). Boll. della Soc. Geol. Ital. 1990, 109, 103–114. [Google Scholar]
- Senowbari-Daryan, B.; Di Stefano, P.; Abate, B. Hypercalcified Sponges from the Upper Triassic (Norian-Rhaetian) Reefs of Sicily; Quaderni del Museo Geologico: Palermo, Italy, 2015; pp. 1121–1415. [Google Scholar]
- Senowbari-Daryan, B.; Link, M. Hypercalcified Segmented Sponges (“sphinctozoans”) from the Upper Triassic (Norian) Reef Boulders of Taurus Mountains (Southern Turkey). Facies 2011, 57, 663–693. [Google Scholar] [CrossRef]
- Senowbari-Daryan, B.; Link, M. A New Thalamid Sponge from the Upper Triassic (Norian) Reef Limestones of the Antalya Region (Turkey). Acta Geol. Hung. 1998, 42, 343–354. [Google Scholar]
- Senowbari-Daryan, B.; Link, M.; Garcia-Bellido, D.C. Fanthalamia Kadiri n. Sp., a New“Sphinctozoan” Sponge from the Triassic (Carnian) of Turkey. Stud. UBB Geol. 2003, 48, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Okay, A.I.; Altiner, D. Uppermost Triassic Limestone in the Karakaya Complex- Stratigraphic and Tectonic Significance. Turk. J. Earth Sci. 2004, 13, 187–199. [Google Scholar]
- Senowbari-Daryan, B.; Link, M. Bicoelia Corticifera, a New Inozoid Sponge from the Upper Triassic (Norian) Reef Boulders of the Central Taurids (Southern Turkey). Turk. J. Earth Sci. 2014, 23, 575–579. [Google Scholar] [CrossRef] [Green Version]
- Martindale, R.C.; Corsetti, F.A.; Bottjer, D.J.; Senowbari-Daryan, B. Microbialite Fabrics and Diminutive Skeletal Bioconstructors in Lower Norian Summit Point Reefs, Oregon, United States. Palaios 2012, 27, 489–508. [Google Scholar] [CrossRef]
- Wood, R. Reef Evolution; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Nakazawa, T.; Ueno, K.; Wang, X. Sedimentary Facies of Carboniferous-Permian Mid-Oceanic Carbonates in the Changning-Menglian Belt, West Yunnan, Southwest China: Origin and Depositional Process. Isl. Arc 2009, 18, 94–107. [Google Scholar] [CrossRef]
- Milliman, J.D. Marine Carbonates; Springer: New York, NY, USA, 1974. [Google Scholar]
- Milliman, J.D. Four Southwestern Caribbean Atolls: Courtown Cays, Albuquerque Cays, Roncador Bank and Serrana Bank. Atoll Res. Bull. 1969, 129, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Rankey, E.C.; Reeder, S.L. Holocene Ooids of Aitutaki Atool, Cook Islands, South Pacific. Geology 2009, 37, 971–974. [Google Scholar] [CrossRef]
- Caggiati, M.; Gianolla, P.; Breda, A.; Celarc, B.; Preto, N. The Start-up of the Dolomia Principale/Hauptdolomit Carbonate Platform (Upper Triassic) in the Eastern Southern Alps. Sedimentology 2018, 65, 1097–1131. [Google Scholar] [CrossRef]
- Haas, J.; Piros, O.; Budai, T.; Gorog, A.; Mandl, W.G.; Loobitzer, H. Transition Between the Massive Reef-Backreef and Cyclic Lagoon Facies of the Dachstein Limestone in the Southern Part of the Dachstein Plateau. Abh. der Geol. Bundesanst. 2010, 65, 35–56. [Google Scholar]


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
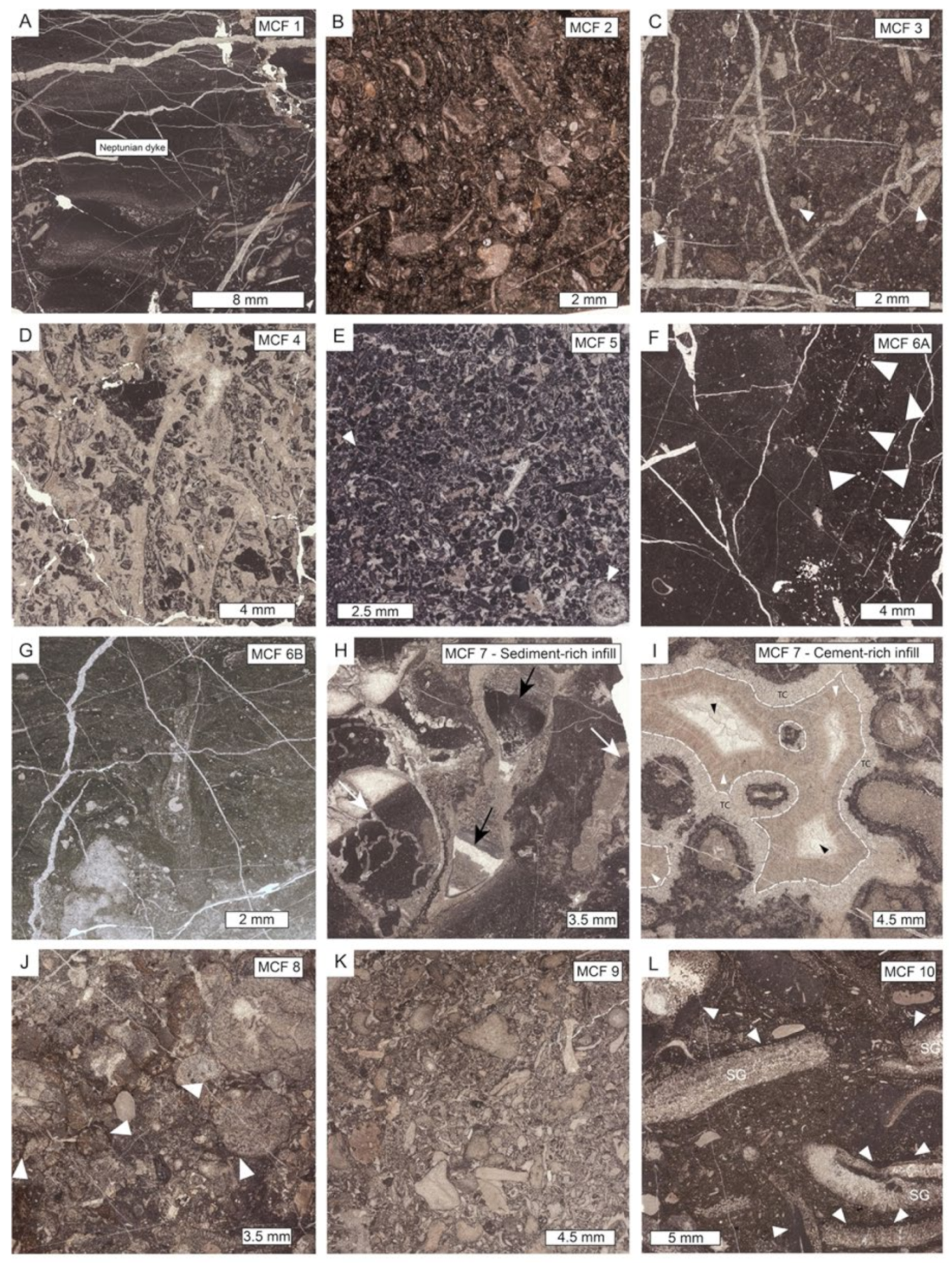
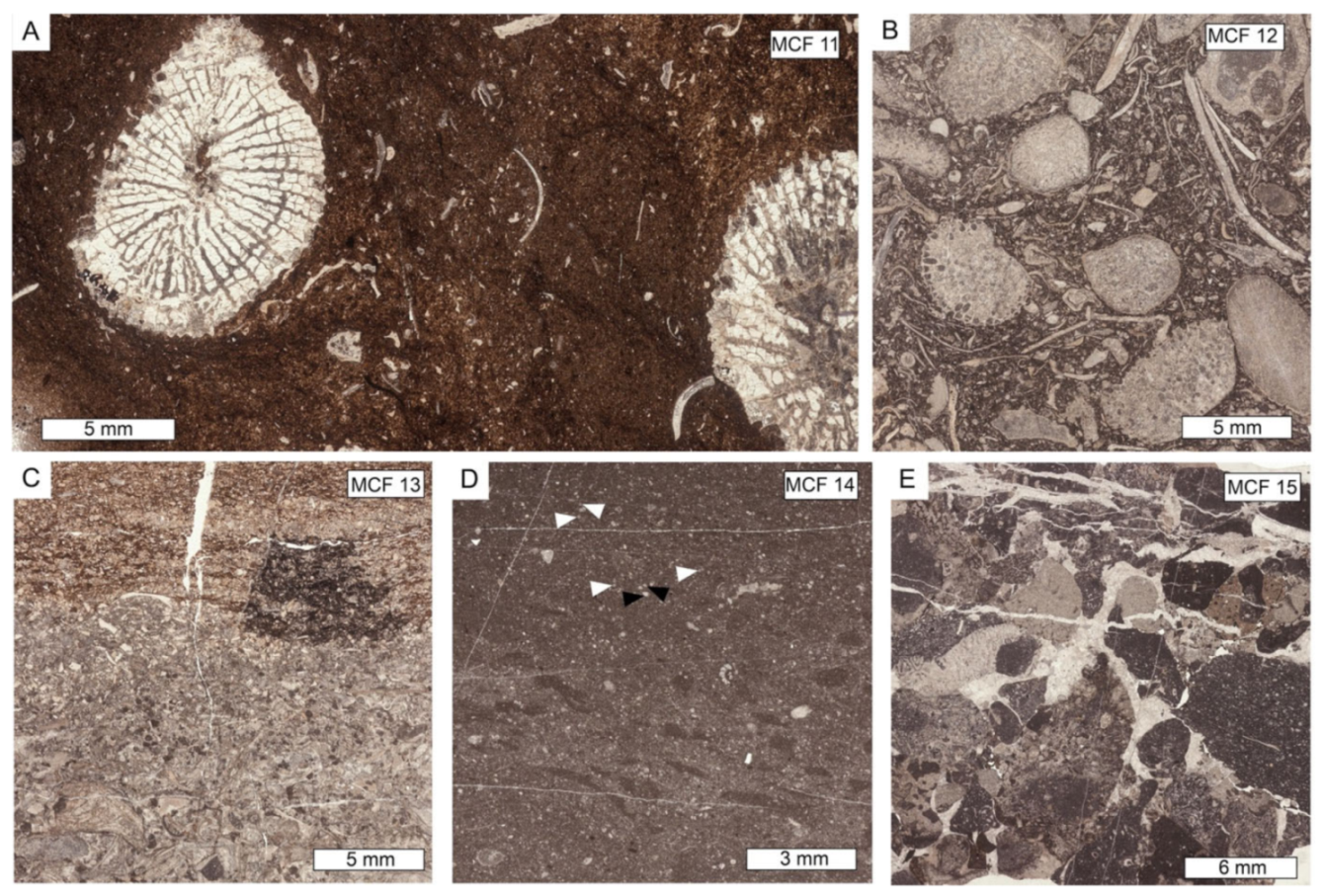
| Microfacies | Lithofacies | Biotic Content and Foraminifers Assemblage | Other Components, Sedimentary Features and Other Remarks | Interpreted Depositional Environment |
|---|---|---|---|---|
| MCF 1—Bioclastic wackestone to floatstone | MM | Molluscs, crinoids+E12, dasycladaleans algae (frequent ? Clypeina sp.), calcareous sponges, solenoporacean algae, ostracods. Foraminifera assemblages include very abundant Taanella yukonensis, textulariidae, common Duostominina, Miliolidae, Wernlina reidae, rare nodosariidae and Agathammina sp. | Frequent neptunian dykes filled by clotted and peloidal micrite and/or very dark micrite | Inner platform (open) |
| MCF 2—Dasycladales-rich wackestone to floatstone | MTB | Dasycladalean green algae Holosporella? rossanae, Patruliuspora oregonica and P. pacifica, gastropods, bivalves, ostracods, spongiomorphs, echinoderms (echinoids and ophiurids), coral fragments, sponge fragments, megalodontid fragments, calcified cyanobacteria (Girvanella sp.), belemnites, Baccanella floriformis. Rich and diversified foraminifera assemblage dominated by very abundant Aulosina oberhauseri, Parvalamella spp., abundant Wernlina reidae, Taanella yukonensis, less common Aulotortus spp., Frentzenella spp., Licispirella sp., Wallowaconus oregonensis, Falsoreinholdella spp., Praerheinoldella spp., Robertonella spp., Glutameandrata vallieri, unidentified Glutameandratidae, and rare Lamelliconus sp., Trocholina spp., Eoguttulina sp., Ichtyolariidae, Trochosiphonia sp. Nodosariidae, Polymorphinidae, Tubulastella comans, Duostominina, Diplotremina sp., ?Praegubkinella sp., ?Cycloforina sp., Agathammina sp., Hoyenella sp., Orthella sp., Opthalmipora sp., Ammodiscidae, Ammodiscus spp., unidentified Miliolidae, Ammovertellinae, Duotaxis sp. and textulariidae | Beds containig this microfacies have a very dark color and have a fetid smell probably due to high organic matter and H2S content. Moreover, they bear large gastropods which are as long as 50 cm. Rest of oxidized ligneous fragments are common. Oncoids, microborings and micritic envelopes are very common. The matrix, often biorturbated and organic-rich, varies from being mud-dominated to very-fine bioclastic | Inner platform (restricted) |
| MCF 3—Peloidal and bioclastic packstone to rare grainstone | MTB and VTB west of K2 | Molluscs (bivalves and gastropods), calcified cyanobacteria (mostly Cayeuxia sp.), dasycladalean green algae, serpulids, ostracods, rare brachiopods, echinoid spines, foraminifers, rare crinoids and fragments of reef-derived biota such as sponges, serpulids and red algae. At times, debris of organisms resembling very small phaceloid corals are abundant. Among the non-skeletal grains peloids and extraclasts of different shapes and sizes are abundant. Foraminifera assemblages are restricted to very abundant Taanella yukonensis, common Miliolidae, Duostiminina, Glomospira sp., unidentified glutameandratidae, rare Decapoalina schaeferae and Involutinina. | Beds containig this microfacies have a very dark in color. Micritic envelope and micritized grains are very abundant. The intergranular porosity is filled either by cement or by a matrix which is fine bioclastic to micritic in nature. | Inner platform (restricted) |
| MCF 4—Bioclastic rudstone | MM | Gastropods, bivalves, foraminifera, serpulids, corals, brachiopods, echinoderms, sponges. Foraminifera assemblages restricted to Duostominina | Angular extraclats (resembling MCF 4) and peloids. Aragonitic allochems (e.g., bivalves) have been dissolved. Rim of isopachous drusy cement and syntaxial overgrowths on some clasts; intergranular porosity is filled by a whitish to yellowish granular mosaic cement | Inner platform (open), tidal channel |
| MCF 5—Peloidal and bioclastic grainstone | TM and MM | Molluscs (bivalves and gastropods), dasycladalean green algae (?Clypeina sp. and possibly ?Kantia sp.), ostracods, crinoids and fragments of reef-derived biota. Foraminifera assemblages are constituted by very abundant Glomospira sp., common Duostominina, Miliolidae, Glutameandratidae and rare Decapoalina schaeferae and Taanella yukonensis | Peloids, extraclast. Micritization processes are widely diffused and led either the formation of a micritic envelope or to the complete micritization of the allochems | Margin area, sand banks |
| MCF 6A—Thrombolitic bindtsone | MM | Microbialite encrusted by Planiinvoluta carinata and other small sphinctozoan sponges. Areas of clotted and peloidal micrite show the presence of ostracods and foraminifera (textulariids, nodosariids and miliolids). | Microbialite acts as framebuilder. Microbial fabric has either a leiolitic to thrombolytic appearance | Outermost margin area |
| MCF 6B—Stromatolitic bindstone | MM | Microbialite encrusted by small ponges, Tolypammina sp. Microtubus communis, Terebella sp., serpulids and Radiomura cautica. The areas between the microbial fabric bear crinoids, molluscs and ostracods. Foraminifera assemblages are restricted to common textulariids and rare unidentified nodosariidae and miliolids. | Microbialite acts as framebuilder. Microbial fabric has either a stromatolitic appearance | Inner platform |
| MCF 7—Metazoans framestone | TM and MM | Inozoan sponges, sphinctozoan sponges, Spongiomorpha ramosa and gibbosa, disjectoporoids sponges, chaetetids sponges, corals, phylloid and solenoporaceans algae, serpulids, brachiopods, ostracods, calcified cyanobacteria, microproblematica. Foraminifers assemblages are dominated by sessile forms like Planiinvoluta carinata, Tolypammina sp., Alphinophragmium sp. and Hirsutospirella pilosa which are found encrusting the main framework. Within the sediment which infills the cavities we find Textularia sp., unidentified miliolids, Agathammina sp., Gsolbergella sp., Hoyenella sp., Sigmoilina sp., Hydrania dulloi, Ophthalmidium sp., Paraophthalmidium sp., unidentified textulariids, unidentified glutameadratidae, Ammodiscus sp., unidentified Ammodiscidae? Endotebanella sp., ?Glomospira sp., Taanella yukonensis, nodosariidae, duostominina | Microbialite is abundant and acts both as binder of pre-existing framework and as framebuilder. Numerous primary (framework) and secondary (dissolution and/or decaying) cavities which are filled either by cements, muddy extraclasts, peloidal micrite or very dark micrite | Margin area, reef |
| MCF 8—Sponges bindstone | TM | Sponges, solenoporacenas algae, calcified cyanobacteria, echinoderms and rare corals. Foraminifera assemblages are restricted to very abundant specimens of Duostominina, Taanella yukonensis, rare Aulosina oberhauseri, Coronipora sp? Lamelliconus sp., Robertonella sp., Gsolbergella sp., Hoyenella sp., Wernlina reidae and unidentified Ammodiscidae | Cavities are very rare. Microbialite acts as binder of a pre-existing substrate | Margin area, crest |
| MCF 9—Crinoidal grainstone to rudstone | TM and VTB | Crinoids, calcified cyanobacteria (Cayeuxia sp.), calcareous sponges (chaetetids the most significant), molluscs. Foraminifera assemblages restricted to abundant Duostominina and rare unidentified Miliolidae | Micritic enveloppes around grains are very frequent. Intergranular porosity filled by a whitish granular mosaic calcite and rarely by micrite | Upper slope |
| MCF 10—Spongiomorpha gibbosa packestone to floatstone | VTB | Spongiomorpha gibbosa, echinoderms, brachiopods, bivalves, ostracods, dasycladalean algae (Clypeina sp.). Foraminifera assemblage is mostly dominated by very abundant Duostominina, Wernlina reidae and Taanella yukonensis | Matrix is formed by dark micrite. Very frequent microbial encrustations | Slope |
| MCF 11—Retiophyllia sp. framestone to floastone | VTB | Presence of Retiophyllia sp. either in living position or as debris. The matrix between the coral branches is given by a wackestone rich in sponge spicules, molluscs (small gastropods and bivalves), echinoderms and ostracods. Foraminifera assemblages are restricted to very abundant Nodosaridae, and rare Wallowaconus oregonensis, Ammodiscus spp. and other unidentified Ammodiscidae | When found in living position, Retiophyllia sp. forms small 30–40 cm wide thickets. Its branches are often silicified and without biogenic encrustations or synsedimentary cement | Slope |
| MCF 12—Sponges-molluscs packstone to rudstone | TMB | Sponges (common chaetetids, Spongiomorpha sp. inozoans and sphinctozoans), calcified cyanobacteria (Cayeuxia sp.), coral fragments (Retiophyllia sp. and Montlivaltia sp.), echinoderms, brachiopods, gastropods, bivalves, brachiopods, ostracods, rare green algae (?Kantia sp.). The foraminifera assemblages are formed by abundant Wernlina reidae, Taanella yukonensis, Duostominina, common Wallowaconus oregonensis, Coronipora spp., Nodosaridae, Agathammina spp. and Miliolidae together with rare Diplotremina sp., ?Oberhausella sp., Falsoreinholdella spp., Praereinholdella spp., Robertonella spp., Trochosiphonia josephi, Parvalamella spp., Aulosina oberhauseri, Aulotortus impressus,?Lamelliconus sp., ?Licispirella iranica, ?Turrispirillina spp., Frentzenella frentzeni, Papillaconus kristani, Trocholina spp., Semiinvoluta spp. | Microbial encrustations are quite common | Slope |
| MCF 13—Laminated and graded wackestone to rudstone | TMB | Sponges (common chaetetids, Spongiomorpha sp. inozoans and sphinctozoans), calcified cyanobacteria, molluscs, brachiopods, ostracods, sponge spicules. Foraminifera assemblages restricted to Duostominina and nodosariids | Erosive base, strongly developed lamination and normal grading | Slope/Toe of the slope |
| MCF 14—Radiolarian-rich bioturbated mudstone to wackestone | TMB | Ostracods, recrystallized radiolarians, sponge spicules, gastropods, thin-shelled bivalves, terebratulid brachiopods. Foraminifera assemblages mainly contain nodosariidae and Duostominina | Matrix is muddy to very fine bioclastic | Toe of the slope |
| MCF 15—Grain-supported limestone breccia or conglomerate | LB | Fragments of corals and sponges | Rounded to sub-angular carbonate and volcaniclastic lithoclasts. Intergranular porosity filled by a whitish granular mosaic calcite cement | ? |
| Reef Framework | |||||||||
| Sponges | Corals | Microbialite | Other Framework Organisms (Encrusters) | Tubular Crust | Total Framework | ||||
| Massive mound N = 42 | 25.22 | 0.66 | 20.14 | 2.67 | 10.61 | 59.29 | |||
| (0–139) | (0–34) | (0–126) | (0–12) | (0–82) | |||||
| Tabular mound N = 10 | 33.05 | 1.15 | 15.20 | 2.55 | 15.00 | 66.95 | |||
| (46–83) | (0–22) | (12–54) | (1–14) | (0–67) | |||||
| Interstitial Sediments | |||||||||
| Dwellers | Peloids or Ooids | Dense Micrite | Clotted and Peloidal Micrite | Intergranular Cement | Primary Cavities Cement | Secondary Cavities Cement | Interstitial Micrite | Total Interstitial Filling | |
| Massive mound N = 42 | 3.52 | 4.51 | 3.13 | 17.13 | 3.32 | 0.84 | 3.19 | 5.07 | 40.71 |
| (0–39) | (0–47) | (0–50) | (0–120) | (0–39) | (0–10) | (0–10) | (0–70) | ||
| Tabular mound N =10 | 3.15 | 6.05 | 1.70 | 10.10 | 2.55 | 2.70 | 1.80 | 5.00 | 33.05 |
| (1–19) | (3–31) | (0–12) | (6–33) | (1–21) | (0–25) | (0–6) | (0–23) | ||
| Areas | Time Interval | References |
|---|---|---|
| Slovenia | Norian–Rhaetian | [89,200,201,202] |
| Northern Calcareous Alps | Norian–Rhaetian | [9,12,203] |
| Iran | Norian | [14,204,205,206,207,208,209,210,211] |
| Sicily | Norian | [212,213,214,215,216,217,218] |
| Turkey | Norian | [204,219,220,221,222,223,224,225] |
| Oman | Norian | [12] |
| Japan (Reef type 2) | Norian | [18,25] |
| Oregon (Summit Point, USA) | Norian | [214,224] |
| Yukon (Canada) | Norian | [51,52,59,60] and this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Piero, N.; Rigaud, S.; Peybernes, C.; Forel, M.-B.; Farley, N.; Martini, R. Upper Triassic Carbonate Records: Insights from the Most Complete Panthalassan Platform (Lime Peak, Yukon, Canada). Geosciences 2022, 12, 292. https://doi.org/10.3390/geosciences12080292
Del Piero N, Rigaud S, Peybernes C, Forel M-B, Farley N, Martini R. Upper Triassic Carbonate Records: Insights from the Most Complete Panthalassan Platform (Lime Peak, Yukon, Canada). Geosciences. 2022; 12(8):292. https://doi.org/10.3390/geosciences12080292
Chicago/Turabian StyleDel Piero, Nicolò, Sylvain Rigaud, Camille Peybernes, Marie-Beatrice Forel, Nicholas Farley, and Rossana Martini. 2022. "Upper Triassic Carbonate Records: Insights from the Most Complete Panthalassan Platform (Lime Peak, Yukon, Canada)" Geosciences 12, no. 8: 292. https://doi.org/10.3390/geosciences12080292