Effect of Sweet Potato Vine on the Onset of Puberty and Follicle Development in Chinese Meishan Gilts

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Data Record and Sample Collection
2.3. Serum and Follicular Fluid Analyses
2.4. Oxidative Stress Biomarkers
2.5. Total RNA Extraction and Real-Time RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Effect of Sweet Potato Vine on the Estrous and Follicular Development in Meishan Gilt
3.2. Effect of Sweet Potato Vine on the Concentration of Hormone in the Serum and Follicular Fluid
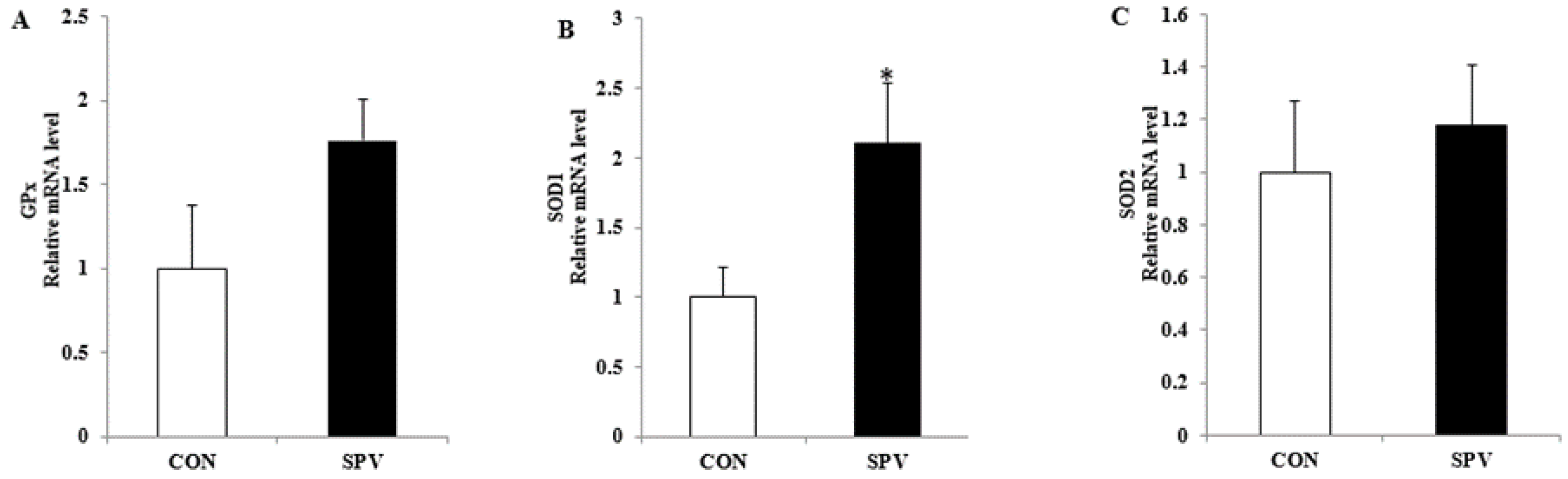
3.3. Effect of Sweet Potato Vine on the Oxidative Stress Status in Meishan Gilts
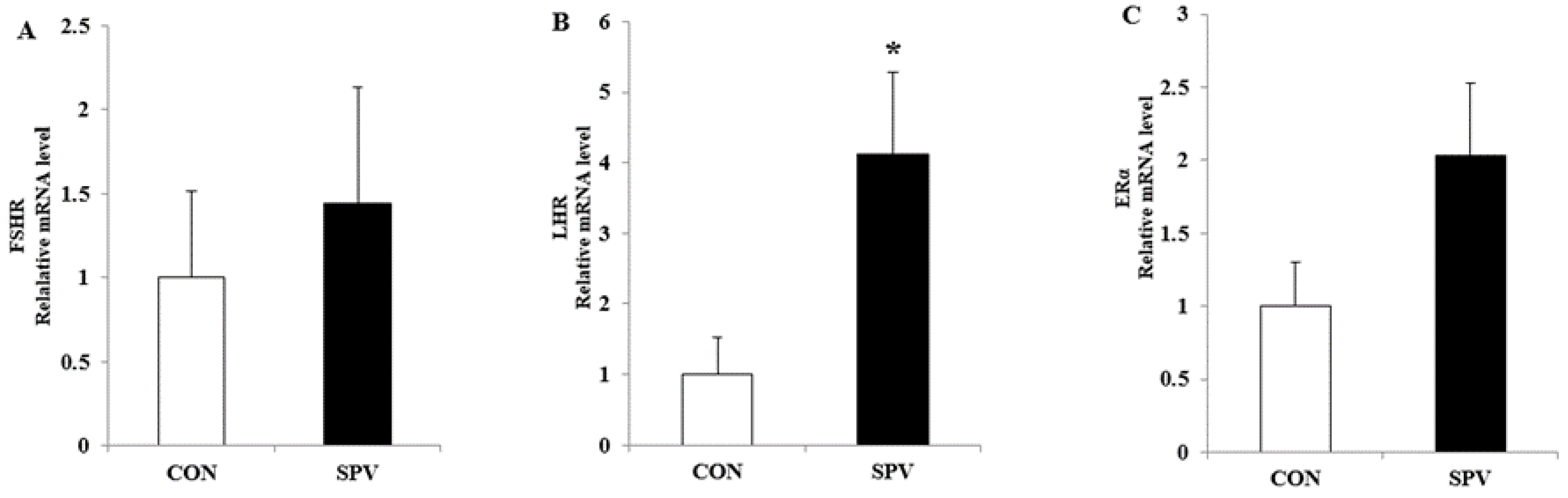
3.4. Effect of Sweet Potato Vine on Antioxidant Stress and Hormone Receptor Related Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nguyen, L.; Everts, H.; Hue, H.; Beynen, A. Feeding of Spinach or Sweet-potato Leaves and Growth Performance of Growing Pigs Kept on Smallholder Farms in Central Vietnam. Trop. Anim. Health Prod. 2004, 36, 815–822. [Google Scholar] [CrossRef]
- Han, I.K.; Yoon, D.J.; Kim, C.S. Studies on the Nutritive Values of Dried Sweetpotato Meal. 1. Partial Substitution of Dried Sweetpotato Meal for Corn in the Ration of Growing Swine. (Korean). Seoul. Natt. Univ. Coll. Agric. Bull. 1976, 1, 67–74. [Google Scholar]
- Dominguez, P.L.; Ly, J. An approach to the nutritional value for pigs of sweet potato vines (Ipomoea batatas (L.) lam). Livest. Res. Rural Dev. 1997, 9, 2. [Google Scholar]
- Phuc, B.H.N.; Lindberg, J.E. Ileal and total tract digestibility in growing pigs fed cassava root meal diets with inclusion of cassava leaves, leucaena leaves and groundnut foliage. Anim. Sci. 2000, 71, 301–308. [Google Scholar] [CrossRef]
- Bazer, F.W.; Thatcher, W.W.; Martinat-Botte, F.; Terqui, M. Sexual maturation and morphological development of the reproductive tract in Large White and prolific Chinese Meishan pigs. Reproduction 1988, 83, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; Natsuaki, M.N.; Neiderhiser, J.M.; Reiss, D. Genetic and Environmental Influences on Pubertal Timing: Results from Two National Sibling Studies. J. Res. Adolesc. 2007, 17, 767–788. [Google Scholar] [CrossRef]
- Hoge, M.D.; Bates, R.O. Developmental factors that influence sow longevity. J. Anim. Sci. 2011, 89, 1238–1245. [Google Scholar] [CrossRef]
- Knauer, M.; Stalder, K.; Serenius, T.; Baas, T.J.; Berger, P.J.; Karriker, L.; Goodwin, R.N.; Johnson, R.K.; Mabry, J.W.; Miller, R.K.; et al. Factors associated with sow stayability in 6 genotypes. J. Anim. Sci. 2010, 88, 3486–3492. [Google Scholar] [CrossRef]
- Hunter, M. Oocyte maturation and ovum quality in pigs. Rev. Reprod. 2000, 5, 122–130. [Google Scholar] [CrossRef]
- Soede, N.M.; Langendijk, P.; Kemp, B. Reproductive cycles in pigs. Anim. Reprod. Sci. 2011, 124, 251–258. [Google Scholar] [CrossRef]
- Tiwari, M.; Prasad, S.; Tripathi, A.; Pandey, A.N.; Ali, I.; Singh, A.K.; Shrivastav, T.G.; Chaube, S.K. Apoptosis in mammalian oocytes: A review. Apoptosis 2015, 20, 1019–1025. [Google Scholar] [CrossRef]
- Hsueh, A.J.W.; Kawamura, K.; Cheng, Y.; Fauser, B.C.J.M. Intraovarian Control of Early Folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.; Zhang, Z.; Chen, X.; Tu, Y. Pig Breeds in China; Institute of Animal Science; Chinese Academy of Agricultural Sciences; Shanghai Science and Technology Press: Shanghai, China, 1986. [Google Scholar]
- Peng, B.; Gao, Y.; Wang, C.; Wang, Y.; Zhang, X.; Han, M. Effects of fresh alfalfa levels and alfalfa meal in the diet on the reproduction performance of sows. Acta Pratacult. Sin. 2011, 20, 219–228. [Google Scholar]
- Zhao, B.Q. Effect of fresh ryegrass on the performance of pregnant sows. Swine Prod. 2014, 3, 22–24. (In Chinese) [Google Scholar]
- Zou, G.Y. Sweet potato and its vine leaves are worth to develop as food sources. Food Sci. (China) 1991, 12, 45–50. [Google Scholar]
- Zhang, L.M.; Wang, Q.M.; Wang, Y.C.H. The Main Nutrient Components and Health Care Function of Sweet Potato. Rain Fed Crops 2003, 23, 162–166. [Google Scholar]
- Xu, M.; Che, L.; Yang, Z.; Zhang, P.; Shi, J.; Li, J.; Lin, Y.; Fang, Z.; Che, L.; Feng, B.; et al. Effect of High Fat Dietary Intake during Maternal Gestation on Offspring Ovarian Health in a Pig Model. Nutrients 2016, 8, 498. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Roongsitthichai, A.; Cheuchuchart, P.; Chatwijitkul, S.; Chantarothai, O.; Tummaruk, P. Influence of age at first estrus, body weight, and average daily gain of replacement gilts on their subsequent reproductive performance as sows. Livest. Sci. 2013, 151, 238–245. [Google Scholar] [CrossRef]
- Tummaruk, P.; Lundeheim, N.; Einarsson, S.; Dalin, A.-M. Effect of birth litter size, birth parity number, growth rate, backfat thickness and age at first mating of gilts on their reproductive performance as sows. Anim. Reprod. Sci. 2001, 66, 225–237. [Google Scholar] [CrossRef]
- Teague, H.S. The Influence of Alfalfa on Ovulation Rate and Other Reproductive Phenomena in Gilts. J. Anim. Sci. 1955, 14, 621–627. [Google Scholar] [CrossRef]
- Pelletier, G.; Labrie, C.; Labrie, F. Localization of oestrogen receptor alpha, oestrogen receptor beta and androgen receptors in the rat reproductive organs. J. Endocrinol. 2000, 165, 359–370. [Google Scholar] [CrossRef]
- Scott, R.T.; Hofmann, G.E.; Muasher, S.J.; Acosta, A.A.; Kreiner, D.K.; Rosenwaks, Z. Correlation of follicular diameter with oocyte recovery and maturity at the time of transvaginal follicular aspiration. J. Assist. Reprod. Genet. 1989, 6, 73–75. [Google Scholar] [CrossRef]
- Foxcroft, G.R. Relationships between luteinizing hormone, follicle-stimulating hormone and prolactin secretion and ovarian follicular development in the weaned sow. Biol. Reprod. 1987, 36, 175–191. [Google Scholar] [CrossRef]
- Ferguson, E.; Ashworth, C.; Edwards, S.; Hawkins, N.; Hepburn, N.; Hunter, M. Effect of different nutritional regimens before ovulation on plasma concentrations of metabolic and reproductive hormones and oocyte maturation in gilts. Reproduction 2003, 126, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Monget, P.; Monniaux, D. Growth factors and the control of folliculogenesis. J. Reprod. Fertil. Suppl. 1995, 49, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Mazerbourg, S.; Bondy, C.; Zhou, J.; Monget, P. The Insulin-like Growth Factor System: A Key Determinant Role in the Growth and Selection of Ovarian Follicles? A Comparative Species Study. Reprod. Domest. Anim. 2003, 38, 247–258. [Google Scholar] [CrossRef]
- Zhou, J.; Kumar, T.R.; Matzuk, M.M.; Bondy, C. Insulin-Like Growth Factor I Regulates Gonadotropin Responsiveness in the Murine Ovary. Mol. Endocrinol. 1997, 11, 1924–1933. [Google Scholar] [CrossRef]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular Growth and Atresia in Mammalian Ovaries: Regulation by Survival and Death of Granulosa Cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.P.; Ratto, M.H. Ovulation-inducing factor in seminal plasma: A review. Anim. Reprod. Sci. 2013, 136, 148–156. [Google Scholar] [CrossRef]
- Wu, D.; Cheung, Q.C.-K.; Wen, L.; Li, J. A Growth-Maturation System That Enhances the Meiotic and Developmental Competence of Porcine Oocytes Isolated from Small Follicles. Biol. Reprod. 2006, 75, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Yding Andersen, C. LHR gene expression in human granulosa cells. Fertil. Wkly. 2013, 1, 5. [Google Scholar]
- Godfrey, K.M.; Barker, D.J. Fetal nutrition and adult disease. Am. J. Clin. Nutr. 2000, 71, 1344s–1352s. [Google Scholar] [CrossRef]
- Poston, L.; Igosheva, N.; Mistry, H.D.; Seed, P.T.; Shennan, A.H.; Rana, S.; Karumanchi, S.A.; Chappell, L.C. Role of oxidative stress and antioxidant supplementation in pregnancy disorders. Am. J. Clin. Nutr. 2011, 94, 1980S–1985S. [Google Scholar] [CrossRef]
- Chew, B.P. Antioxidant vitamins affect food animal immunity and health. J. Nutr. 1995, 125, 1804–1808. [Google Scholar]
- Gardner, P.T.; White, T.A.; McPhail, D.B.; Duthie, G.G. The relative contributions of vitamin C, carotenoids and phenolics to the antioxidant potential of fruit juices. Food Chem. 2000, 68, 471–474. [Google Scholar] [CrossRef]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Hułas-Stasiak, M.; Dobrowolski, P.; Tomaszewska, E.; Kostro, K. Maternal acrylamide treatment reduces ovarian follicle number in newborn guinea pig offspring. Reprod. Toxicol. 2013, 42, 125–131. [Google Scholar] [CrossRef]
- Van Blerkom, J.; Antczak, M.; Schrader, R. The developmental potential of the human oocyte is related to the dissolved oxygen content of follicular fluid: Association with vascular endothelial growth factor levels and perifollicular blood flow characteristics. Hum. Reprod. 1997, 12, 1047–1055. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Moore, J.; Wood, J.M.; Beazley, W.D.; Gaze, D.C.; Tobin, D.J.; Marshall, H.S.; Panske, A.; Panzig, E.; Hibberts, N.A. In Vivo and In Vitro Evidence for Hydrogen Peroxide (H2O2) Accumulation in the Epidermis of Patients with Vitiligo and its Successful Removal by a UVB-Activated Pseudocatalase. J. Investig. Dermatol. Symp. Proc. 1999, 4, 91–96. [Google Scholar] [CrossRef]
- Dalto, D.B.; Roy, M.; Audet, I.; Palin, M.-F.; Guay, F.; Lapointe, J.; Matte, J.J. Interaction between vitamin B6 and source of selenium on the response of the selenium-dependent glutathione peroxidase system to oxidative stress induced by oestrus in pubertal pig. J. Trace Elem. Med. Biol. 2015, 32, 21–29. [Google Scholar] [CrossRef]
- Roy, C.; Lavoie, M.; Richard, G.; Archambault, A.; Lapointe, J. Evidence that oxidative stress is higher in replacement gilts than in multiparous sows. J. Anim. Physiol. Anim. Nutr. 2016, 100, 911–919. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Basal Experimental Diet 1 | Sweet Potato Vine 2 | |
|---|---|---|
| Ingredient, % | ||
| Corn, 7.8% CP | 64.79 | - |
| Soybean meal, 44.2% CP | 14.42 | |
| Wheat bran | 18.00 | |
| L-Lysine-HCL (75%) | 0.06 | |
| Calcium carbonate | 1.00 | |
| Dicalcium phosphate | 0.23 | |
| Salt | 0.40 | |
| Choline chloride (50%) | 0.10 | |
| Mineral premix 3 | 0.50 | |
| Vitamin premix 4 | 0.50 | |
| Total | 100.00 | |
| Nutrient level | ||
| DM, % | - | 10.13 |
| DE, kcal/kg | 3080.00 | 2837.00 |
| CP, % | 14.00 | 14.90 |
| T-Lys, % | 0.68 | - |
| CF, % | 3.11 | 15.51 |
| NDF, % | - | 21.63 |
| ADF, % | - | 19.37 |
| ADL, % | - | 3.30 |
| Ca, % | 0.53 | 1.61 |
| TP, % | 0.48 | 0.47 |
| AP, % | 0.20 | - |
| VA, mg/kg | - | 15.00 |
| VC, mg/kg | - | 123.00 |
| VE, mg/kg | - | 16.00 |
| Carotene, mg/kg | - | 42.00 |
| Fe, mg/kg | - | 11.00 |
| CON | SPV | |
|---|---|---|
| Basal diets, kg/d | 1.64 | 1.46 |
| Sweet potato vine (fresh base), kg/d | -- | 2.00 |
| DE intake, kcal/d | 5051.00 | 5072.00 |
| CP intake, g/d | 230.00 | 234.00 |
| CF intake, g/d | 51.00 | 73.94 |
| Gene 1 | Primers | Sequence (5′-3′) | Accession Number | Product Size (bp) |
|---|---|---|---|---|
| GPx | Forward | GCTCGGTGTATGCCTTCTCT | NM_214201.1 | 103 |
| Reverse | AGCGACGCTACGTTCTCAAT | |||
| SOD1 | Forward | GAGCTGAAGGGAGAGAAGACAGT | NM_001190422.1 | 116 |
| Reverse | GCACTGGTACAGCCTTGTGTAT | |||
| SOD2 | Forward | CTGGACAAATCTGAGCCCTAAC | NM_214127.2 | 118 |
| Reverse | GACGGATACAGCGGTCAACT | |||
| FSHR | Forward | TCACAGTCCCTCGGTTCCTT | NM_214386.1 | 152 |
| Reverse | AGCATCACAGCCTGCTCCA | |||
| LHR | Forward | ATGGGGCTCTACCTGCTACTCA | NM_214449.1 | 255 |
| Reverse | GAGCCACCCTCCAAGCATAA | |||
| ERα | Forward | ATGAAGTGCAAGAACGTGGTG | NM_214220.1 | 151 |
| Reverse | AATGCGATGGAGTTGAGCC | |||
| β-actin | Forward | GGCCGCACCACTGGCATTGTCAT | DQ845171.1 | 104 |
| Reverse | AGGTCCAGACGCAGGATGGCG |
| CON | SPV | p-Value | |
|---|---|---|---|
| Initial body weight, kg | 30.30 ± 0.30 | 30.20 ± 0.30 | 0.92 |
| Body weight at first estrus, kg | 38.80 ± 1.30 | 42.80 ± 1.80 | 0.09 |
| Age at puberty, d | 135.30 ± 3.10 | 144.70 ± 3.90 | 0.08 |
| Slaughtered body weight, kg | 62.20 ± 2.20 | 70.20 ± 3.40 | 0.07 |
| Slaughtered age, d | 197.70 ± 2.30 | 207.40 ± 4.90 | 0.09 |
| Duration of the first estrus cycle, d | 21.00 ± 0.60 | 21.10 ± 0.40 | 0.89 |
| Duration of the second estrus cycle, d | 20.80 ± 0.60 | 20.70 ± 0.50 | 0.92 |
| CON | SPV | p-Value | |
|---|---|---|---|
| Ovarian weight, g | 6.00 ± 0.70 b | 8.10 ± 0.30 a | 0.01 |
| Ovarian relative weight, g/kg | 0.095 ± 0.008 b | 0.116 ± 0.004 a | 0.05 |
| Uterus weight, g | 349.60 ± 56.00 b | 473.80 ± 23.30 a | 0.04 |
| Uterus relative weight, g/kg | 5.8 ± 0.90 | 6.80 ± 0.70 | 0.35 |
| No. Large follicles (> 5mm) | 12.60 ± 1.50 b | 16.40 ± 0.90 a | 0.04 |
| No. Medium follicles (≥ 3mm, ≤ 5mm) | 4.10 ± 1.00 | 4.70 ± 1.20 | 0.76 |
| No. Small follicles (< 3mm) | 26.80 ± 5.00 | 30.90 ± 7.60 | 0.67 |
| No. Corpora lutea | 15.00 ± 4.60 | 25.30 ± 1.00 | 0.06 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Cao, M.; Li, J.; Lin, Y.; Fang, Z.; Che, L.; Feng, B.; Zhuo, Y.; Wang, J.; Wu, D.; et al. Effect of Sweet Potato Vine on the Onset of Puberty and Follicle Development in Chinese Meishan Gilts. Animals 2019, 9, 297. https://doi.org/10.3390/ani9060297
Zhang P, Cao M, Li J, Lin Y, Fang Z, Che L, Feng B, Zhuo Y, Wang J, Wu D, et al. Effect of Sweet Potato Vine on the Onset of Puberty and Follicle Development in Chinese Meishan Gilts. Animals. 2019; 9(6):297. https://doi.org/10.3390/ani9060297
Chicago/Turabian StyleZhang, Pan, Meng Cao, Jian Li, Yan Lin, Zhengfeng Fang, Lianqiang Che, Bin Feng, Yong Zhuo, Jianping Wang, De Wu, and et al. 2019. "Effect of Sweet Potato Vine on the Onset of Puberty and Follicle Development in Chinese Meishan Gilts" Animals 9, no. 6: 297. https://doi.org/10.3390/ani9060297