Vegetable Oils Rich in Polyunsaturated Fatty Acids Supplementation of Dairy Cows’ Diets: Effects on Productive and Reproductive Performance


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Experimental Diets
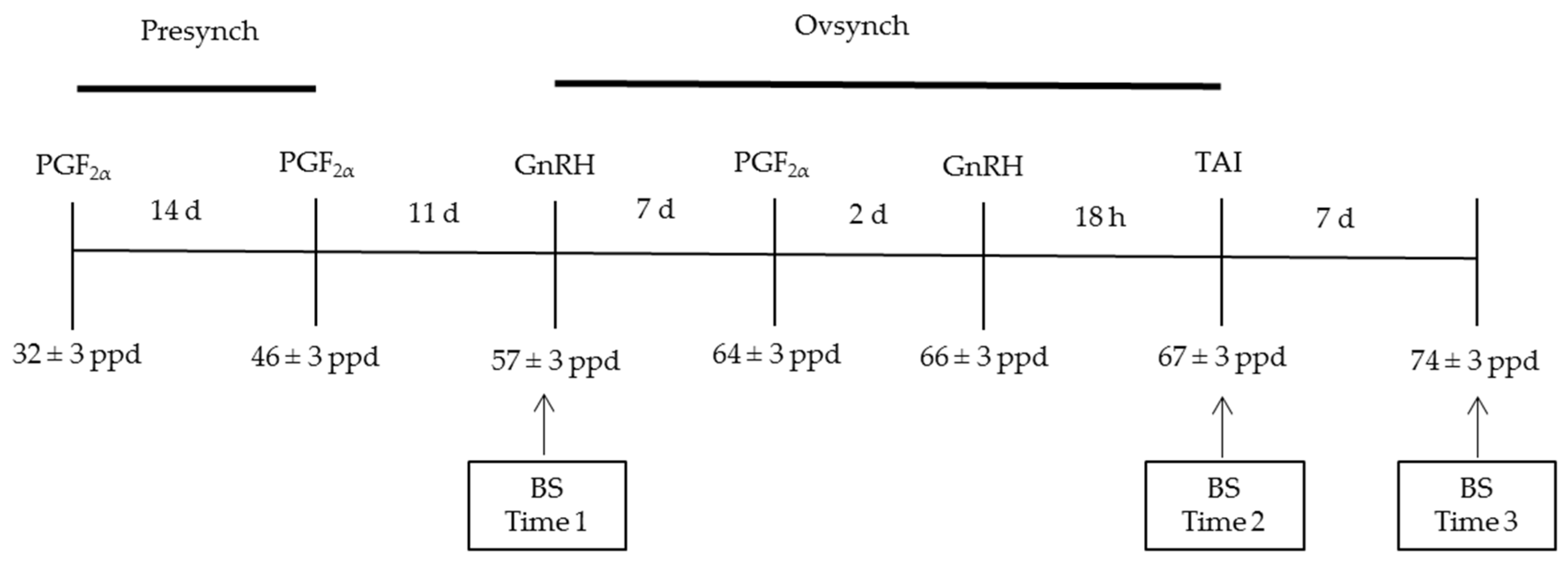
2.2. Reproductive Management
2.3. Experimental Procedure
2.4. Analytical Procedure
2.5. Statistical Analysis
3. Results
3.1. Effects of PUFA Supplementation on Productive Performance
3.2. Effects of PUFA Supplementation on Reproductive Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Staples, C.R.; Burke, J.M.; Thatcher, W.W. Influence of supplemental fats on reproductive tissues and performance of lactating cows. J. Dairy Sci. 1998, 81, 856–871. [Google Scholar] [CrossRef]
- Staples, C.R.; Thatcher, W.W. Effects of fatty acids on reproduction in dairy cows. In Recent Advances in Animal Nutrition; Garnsworthy, P.C., Wiseman, J., Eds.; Nottingham University Press: Nottingham, UK, 2005; pp. 229–256. [Google Scholar] [CrossRef]
- Gulliver, C.E.; Friend, M.A.; King, B.J.; Clayton, E.H. The role of omega-3 polyunsaturated fatty acids in reproduction of sheep and cattle. Anim. Reprod. Sci. 2012, 131, 9–22. [Google Scholar] [CrossRef]
- Kemp, B.; Soede, N.M.; Kankofer, M.; Bevers, M.; Taverne, M.A.; Wensing, T.; Noordhuizen, J.P.T.M. Influence of linoleic/linolenic acid ratio in the diet of periparturient cattle on plasma concentrations of PGF2α, metabolite and placental expulsion rate. Theriogenology 1998, 49, 571–580. [Google Scholar] [CrossRef]
- Gulliver, C.E.; Piltz, J.W.; Friend, M.A.; Clayton, E.H. Improving the omega-3 status of sheep by feeding silage. In Proceedings of the Nutrition Society of Australia, Perth, Australia, 30 November–3 December 2010; Volume 34, p. 73. [Google Scholar] [CrossRef]
- Scollan, N.; Hocquette, J.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A.P. Innovations in beef production systems that enhance the nutritional health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Fisher, W.J.; Tweed, J.K.; Wilkins, R.J. Comparison of grass and legume silages for milk production 1. Production responses with different levels of concentrate. J. Dairy Sci. 2003, 86, 2598–2611. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kinoshita, M.; Ohnishi, M.; Fukui, Y. Lipid and fatty acid analysis of fresh and frozen-thawed immature and in vitro matured bovine oocytes. Reproduction 2001, 122, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Shingfield, K.J.; Chilliard, Y.; Toivonen, V.; Kairenius, P.; Givens, D.I. Trans fatty acids and bioactive lipids in ruminant milk. Adv. Exp. Med. Biol. 2008, 606, 3–65. [Google Scholar] [CrossRef]
- Kliem, K.E.; Shingfield, K.J. Manipulation of milk fatty acid composition in lactating cows: Opportunities and challenges. Eur. J. Lipid Sci. Technol. 2016, 118, 1661–1683. [Google Scholar] [CrossRef]
- Fuentes, M.C.; Calsamiglia, S.; Sánchez, C.; González, A.; Newbold, J.R.; Santos, J.E.P.; Rodríguez-Alcalá, L.M.; Fontecha, J. Effect of extruded linseed on productive and reproductive performance of lactating dairy cows. Livest. Sci. 2008, 113, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Brunschwig, P.; Hurtaud, C.; Chilliard, Y.; Glasser, F. L’apport de lin dans la ration des vaches laitières: Effets sur la production, la composition du lait et des produits laitiers, les émissions de méthane et les performances de reproduction. INRA Prod. Anim. 2010, 23, 307–318. [Google Scholar]
- Institut National de la Recherche Agronomique (INRA). Alimentation des bovins, ovins et caprins. Besoins des animaux-Valeurs des aliments-Tables INRA 2007; Editions Quae: Versailles, France, 2007. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). Native Starch Determination of Starch Content. Ewers Polarimetric Method ISO10520; ISO: Geneva, Switzerland, 1997. [Google Scholar]
- International Dairy Federation (IDF). Determination of Milk Fat, Protein and Lactose Content. Guidance on the Operation of Mid-Infrared Instruments; IDF Standard 141C: Brussels, Belgium, 2000. [Google Scholar]
- Sukhija, P.S.; Palmquist, D.L. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agric. Food Chem. 1988, 36, 1202–1206. [Google Scholar] [CrossRef]
- Kramer, J.K.; Seat, N.; Dugan, M.E.R.; Mossoba, M.M.; Yurawecz, M.P.; Roach, J.A.; Eulitz, K.; Aalhus, J.L.; Schaefer, A.L.; Ku, Y. Distributions of conjugated linoleic acid (CLA) isomers in tissue lipid classes of pigs fed a commercial CLA mixture determined by gas chromatography and silver ion-high performance liquid chromatography. Lipids 1998, 33, 549–558. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–508. [Google Scholar] [PubMed]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar]
- Ury, H.K. A comparison of four procedures for multiple comparisons among means (pairwise contrasts) for arbitrary samples sizes. Technometrics 1976, 18, 89–97. [Google Scholar] [CrossRef]
- Kelsey, J.A.; Corl, B.A.; Collier, R.J.; Bauman, D.E. The effect of breed parity and stage of lactation on conjugated linoleic acid (cla) in milk fat from dairy cows. J. Dairy Sci. 2003, 86, 2588–2597. [Google Scholar] [CrossRef]
- Corl, B.A.; Baumgard, L.H.; Dwyer, D.A.; Griinari, J.M.; Phillips, B.S.; Bauman, D.E. The role of ∆9-desaturase in the production of cis-9,trans-11 CLA. J. Nutr. Biochem. 2001, 12, 622–630. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Huang, Y.; Schoonmaker, J.P.; Bradford, B.J.; Beitz, D.C. Response of milk fatty acid composition to dietary supplementation of soy oil, conjugated linoleic acid, or both. J. Dairy Sci. 2008, 91, 260–270. [Google Scholar] [CrossRef]
- Flowers, G.; Ibrahim, S.A.; AbuGhazaleh, A.A. Milk fatty acid composition of grazing dairy cows when supplemented with linseed oil. J. Dairy Sci. 2008, 91, 722–730. [Google Scholar] [CrossRef]
- Sutton, J.D.; Knight, R.; McAllan, A.B.; Smith, R.H. Digestion and synthesis in the rumen of sheep given diets supplemented with free and protected oils. Br. J. Nutr. 1983, 49, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Doreau, M.; Chilliard, Y. Digestion and metabolism of dietary fat in farm animals. Br. J. Nutr. 1997, 78, S15–S35. [Google Scholar] [CrossRef]
- Ueda, K.; Ferlay, A.; Chabrot, J.; Loor, J.J.; Chilliard, Y. Effect of linseed oil supplementation on ruminal digestion in dairy cows fed diets with different forage:concentrate. J. Dairy Sci. 2003, 86, 3999–4007. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Jenkins, T.C. A 100-Year Review: Fat feeding of dairy cows. J. Dairy Sci. 2017, 100, 10061–10077. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Bauman, D.E.; Perfield, J.W.; Harvatine, K.J.; Baumgard, L.H. Regulation of fat synthesis by conjugated linoleic acid: Lactation and the ruminant model. J. Nutr. 2008, 138, 403–409. [Google Scholar] [CrossRef]
- Brent, M.; Kazala, E.K.; Murdoch, G.K.; Keating, A.F. Effect of CLA and other C18 unsaturated fatty acids on DGAT in bovine milk fat biosynthesis systems. Lipids 2008, 43, 903–912. [Google Scholar] [CrossRef]
- Loor, J.; Ferlay, A.; Ollier, A.; Doreau, M.; Chilliard, Y. Relationship among trans and conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J. Dairy Sci. 2005, 88, 726–740. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Reynolds, C.K.; Hervas, G.; Griinari, J.; Grandison, A.S.; Beever, D.E. Examination of the persistency of milk fatty acid composition responses to fish oil and sunflower oil in the diet of dairy cows. J. Dairy Sci. 2006, 89, 714–732. [Google Scholar] [CrossRef]
- Alzahal, O.; Odongo, N.E.; Mutsvanqwa, T.; Or-Rashid, M.M.; Duffield, T.F.; Baqq, R.; Dick, P.; Vessie, G.; McBride, B.W. Effects of monensin and dietary soybean oil on milk fat percentage and milk fatty acid profile in lactating dairy cows. J. Dairy Sci. 2008, 91, 1166–1174. [Google Scholar] [CrossRef]
- Onetti, S.G.; Grummer, R.R. Response of lactating cows to three supplemental fat sources as affected by forage in the diet and stage of lactation: A meta-analysis of literature. Anim. Feed Sci. Technol. 2004, 115, 65–82. [Google Scholar] [CrossRef]
- Schroeder, G.F.; Gagliostro, G.A.; Bargo, F.; Delahoy, J.E.; Muller, D.L. Effects of fat supplementation on milk production and composition by dairy cows on pasture: A review. Livest. Prod. Sci. 2004, 86, 1–18. [Google Scholar] [CrossRef]
- De Peters, E.J.; Cant, J.P. Nutritional factors influencing the nitrogen composition of bovine milk: A review. J. Dairy Sci. 1992, 75, 2043–2070. [Google Scholar] [CrossRef]
- Bu, D.P.; Wang, J.Q.; Dhiman, T.R.; Liu, S.J. Effectiveness of oils rich in linoleic and linolenic acids to enhance conjugated linoleic acid in milk from dairy cows. J. Dairy Sci. 2007, 90, 998–1007. [Google Scholar] [CrossRef]
- Grummer, R.R. Effect of feed on the composition of milk fat. J. Dairy Sci. 1991, 74, 3244–3257. [Google Scholar] [CrossRef]
- Caroprese, M.; Marzano, A.; Marino, R.; Gliatta, G.; Muschio, A.; Sevi, A. Flaxseed supplementation improves fatty acid profile of cow milk. J. Dairy Sci. 2010, 93, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Doreau, M.; Ferlay, A. Digestion and utilization of fatty acids by ruminants. Anim. Feed Sci. Technol. 1994, 45, 379–396. [Google Scholar] [CrossRef]
- Bauman, D.E.; Griinari, J.M. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 2003, 23, 203–227. [Google Scholar] [CrossRef] [PubMed]
- Fievez, V.; Vlaeminck, B.; Dhanoa, M.S.; Dewhurst, R.J. Use of principal component analysis to investigate the origin of heptadecenoic and conjugated linoleic acids in milk. J. Dairy Sci. 2003, 86, 4047–4053. [Google Scholar] [CrossRef]
- Turpeinen, A.; Mutanen, M.; Aro, A.; Salminen, I.; Basu, S.; Palmquist, D.; Griinari, J.M. Bioconversion of vaccenic acid to conjugated linoleic acid in humans. Am. J. Clin. Nutr. 2002, 76, 504–510. [Google Scholar] [CrossRef] [Green Version]
- Demetrio, D.G.B.; Santos, R.M.; Demetrio, C.G.B.; Vasconcelos, J.M.L. Factors affecting conception rates following artificial insemination or embryo transfer in lactating Holstein cows. J. Dairy Sci. 2007, 90, 5073–5082. [Google Scholar] [CrossRef]
- Lopess, C.N.; Scarpa, A.B.; Cappellozza, B.I.; Cooke, R.F.; Vasconcelos, J.M.L. Effects of rumen-protected polyunsaturated fatty acid supplementation on reproductive performance of Bos indicus beef cows. J. Anim. Sci. 2009, 87, 3935–3943. [Google Scholar] [CrossRef]
- Stevenson, J.S.; Pursley, J.R.; Garverick, H.A.; Fricke, P.M.; Kesler, D.J.; Ottobre, J.S.; Wiltbank, M.C. Treatment of cycling and noncycling lactating dairy cows with progesterone during Ovsynch. J. Dairy Sci. 2006, 89, 2567–2578. [Google Scholar] [CrossRef]
- Chebel, R.C.; Al-Hassan, M.J.; Fricke, P.M.; Santos, J.E.P.; Lima, J.R.; Martel, C.A.; Stevenson, J.S.; Garcia, R.; Ax, R.L. Supplementation of progesterone via controlled internal drug release inserts during ovulation synchronization protocols in lactating dairy cows. J. Dairy Sci. 2010, 93, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, D.J.; Kastelic, J.P.; Corbett, R.; Pitney, P.A.; Petit, H.V.; Small, J.A.; Zalkovic, P. Lower pregnancy losses in lactating dairy cows fed a diet enriched in alpha-linolenic acid. J. Dairy Sci. 2006, 89, 3066–3074. [Google Scholar] [CrossRef]
- Stronge, A.; Sreemrn, J.; Diskin, M.; Mee, J.; Kenny, D.; Morris, D. Post insemination milk progesterone concentration and embryo survival in dairy cows. Theriogenology 2005, 64, 1212–1224. [Google Scholar] [CrossRef] [PubMed]
- Mann, G.E.; Fray, M.D.; Lamming, G.E. Effects of time of progesterone supplementation on embryo development and interferon-τ production in the cow. Vet. J. 2006, 171, 500–503. [Google Scholar] [CrossRef]
- Petit, H.V.; Twagiramungu, H. Conception rate and reproductive function of dairy cows fed different fat sources. Theriogenology 2006, 66, 1316–1324. [Google Scholar] [CrossRef]
- Grummer, R.R.; Carroll, D.J. Effects of dietary fat on metabolic disorders and reproductive performance of dairy cattle. J. Anim. Sci. 1991, 69, 3838–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.A.; Kincaid, R.L.; Westberg, H.H.; Gaskins, C.T.; Lamb, B.K.; Cronrath, J.D. The effect of oilseeds in diets of lactating cows on milk production and methane emissions. J. Dairy Sci. 2002, 85, 1509–1515. [Google Scholar] [CrossRef]
- Petit, H.V.; Germiquet, C.; Lebel, D. Effect of feeding whole unprocessed sunflower seeds and flaxseed on milk production milk composition and prostaglandin secretion in dairy cows. J. Dairy Sci. 2004, 87, 3889–3898. [Google Scholar] [CrossRef]
- Caldari-Torres, C.; Lock, A.L.; Staples, C.R.; Badinga, L. Performance, metabolic, and endocrine responses of periparturient Holstein cows fed 3 sources of fat. J. Dairy Sci. 2011, 94, 1500–1510. [Google Scholar] [CrossRef]
- Robinson, J.J.; Pushpakumara, P.G.A.; Cheng, Z.; Peters, A.R.; Abayasekara, D.R.E.; Wathes, D.C. Effects of dietary polyunsaturated fatty acids on ovarian and uterine function. Reproduction 2002, 124, 119–131. [Google Scholar] [CrossRef]
- Taylor, V.J.; Cheng, Z.; Pushpakumara, P.G.A.; Beever, D.E.; Wathes, D.C. Relationships between the plasma concentrations of insulin-like growth factor-I in dairy cows and their fertility and milk yield. Vet. Rec. 2004, 155, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Bilby, T.R.; Sozzi, A.; Lopez, M.M.; Silvestre, F.T.; Ealy, A.D.; Staples, C.R.; Thatcher, W.W. Pregnancy bovine somatotropin and dietary n-3 fatty acids in lactating dairy cows: I. Ovarian conceptus and growth hormone–insulin-like growth factor system responses. J. Dairy Sci. 2006, 89, 3360–3374. [Google Scholar] [CrossRef]
- Reis, M.M.; Cooke, R.F.; Ranches, J.; Vasconcelos, J.L.M. Effects of calcium salts of polyunsaturated fatty acids on productive and reproductive parameters of lactating Holstein cows. J. Dairy Sci. 2012, 95, 7039–7050. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.R.; Palmquist, D.L. High fat diets increase plasma cholecystokinin and pancreatic polypeptide, and decrease plasma insulin and feed intake in lactating cows. J. Nutr. 1996, 126, 2913–2919. [Google Scholar] [CrossRef]
- Williams, G.L.; Stanko, R.L. Dietary fats as reproductive nutraceuticals in beef cattle. J. Anim. Sci. 2000, 77, 1–12. [Google Scholar] [CrossRef]
- Van Knegsel, A.T.M.; Van den Brand, H.; Dijkstra, J.; Tamminga, S.; Kemp, B.R. Effect of dietary energy source on energy balance, production, metabolic disorders and reproduction in lactating dairy cattle. Reprod. Nutr. Dev. 2005, 45, 665–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Item | Diet 1 | ||
|---|---|---|---|
| CON | SOY | LIN | |
| Ingredients (g/kg dry matter basis) | |||
| Corn silage | 181.0 | 186.0 | 186.0 |
| Alfalfa hay | 204.3 | 209.0 | 209.0 |
| Barley straw | 19.6 | 20.3 | 20.3 |
| Soy hulls | 57.2 | 58.8 | 58.8 |
| Barley grain, ground | 28.5 | 162.7 | 162.7 |
| Wheat grain, ground | 179.9 | 78.6 | 78.6 |
| DDGS 2 | 100 | - | - |
| Palm kernel cake | 54.9 | 40.2 | 40.2 |
| Soybean meal | 52.4 | 96.4 | 96.4 |
| Barley sprouts | 38.5 | 39.6 | 39.6 |
| Wheat bran | 52.6 | 54.0 | 54.0 |
| Molasses, sugarcane | 13.3 | 13.6 | 13.6 |
| Soybean oil | - | 23.0 | - |
| Linseed oil | - | - | 23.0 |
| Limestone | 4.0 | 4.0 | 4.0 |
| Dicalcium phosphate | 0.8 | 0.8 | 0.8 |
| Sodium bicarbonate | 9.6 | 9.6 | 9.6 |
| Sodium chloride | 1.7 | 1.7 | 1.7 |
| Mineral and vitamin mix 3 | 1.7 | 1.7 | 1.7 |
| Estimated nutritive value | |||
| UFL 4 | 0.95 | 0.95 | 0.95 |
| PDI 5 | 104.0 | 104.0 | 104.4 |
| Item | Diet 1 | ||
|---|---|---|---|
| CON | SOY | LIN | |
| Chemical composition (g/kg DM) | |||
| Dry matter | 670.0 | 637.2 | 626.8 |
| Crude protein | 171.1 | 163.9 | 163.5 |
| Ether extract | 36.2 | 49.8 | 48.8 |
| Neutral detergent fibre | 306.8 | 314.5 | 329.5 |
| Acid detergent fibre | 219.4 | 234.0 | 252.1 |
| Starch | 272.0 | 228.1 | 217.9 |
| Fatty acids (% of total fatty acids) | |||
| C12:0 | 5.23 | 2.37 | 4.18 |
| C14:0 | 1.82 | 1.06 | 1.6 |
| C16:0 | 25.05 | 20.32 | 18.32 |
| C16:1 | 1.3 | 0.82 | 0.53 |
| C17:0 | 0.32 | 0.29 | 0.21 |
| C17:1 | 0.32 | 0.11 | 0.21 |
| C18:0 | 4.72 | 3.46 | 4.36 |
| C18:1 | 17.54 | 18.68 | 17.55 |
| C18:2 | 32.75 | 42.56 | 28.3 |
| C18:3 | 9.6 | 8.26 | 24.10 |
| C20:0 | 1.17 | 0.41 | 0.39 |
| C20:1 | 0.18 | 1.66 | 0.25 |
| Saturated fatty acids | 38.31 | 27.91 | 29.06 |
| Monounsaturated fatty acids | 19.34 | 21.27 | 18.54 |
| Polyunsaturated fatty acids | 42.35 | 50.82 | 52.41 |
| Item | Diet 1 | SEM 2 | p-Value | ||
|---|---|---|---|---|---|
| CON | SOY | LIN | |||
| Milk YUield (kg/d) | 31.1 | 32.0 | 31.6 | 1.02 | 0.806 |
| 4% fat-corrected milk yield (kg/d) | 28.1 | 26.2 | 25.8 | 1.51 | 0.556 |
| Fat (%) | 3.41 a | 2.65 b | 2.99 b | 0.011 | |
| Fat Yield (kg/d) | 1.06 | 0.88 | 0.89 | 0.068 | 0.098 |
| Protein (%) | 3.33 a | 3.09 b | 3.16 b | 0.062 | 0.022 |
| Protein Yield (kg/d) | 0.98 | 1.00 | 0.98 | 0.061 | 0.921 |
| Item | Diet 1 | SEM 2 | p-Value | ||
|---|---|---|---|---|---|
| CON | SOY | LIN | |||
| Saturated fatty acids (SFA) | 67.84 a | 60.75 b | 62.02 b | 0.951 | <0.0001 |
| C4:0 | 1.22 a | 0.942 b | 1.05 b | 0.049 | 0.002 |
| C8:0 | 0.085 | 0.177 | 0.074 | 0.0441 | 0.222 |
| C12:0 | 3.99 a | 2.76c | 3.62 b | 0.331 | 0.039 |
| C14:0 | 13.65 | 13.33 | 12.89 | 0.528 | 0.611 |
| C16:0 | 41.64 a | 34.86 b | 34.53 b | 0.383 | <0.0001 |
| C18:0 | 7.26 b | 8.98 a | 9.80 a | 0.467 | 0.002 |
| Monounsaturated fatty acids (MUFA) | 26.14 b | 32.01 a | 30.17 a | 0.859 | <0.001 |
| C14:1 | 1.69 a | 1.59 b | 1.39 c | 0.068 | 0.019 |
| C16:1 | 2.83 a | 2.343 b | 2.12 b | 0.172 | 0.023 |
| C18:1 trans-11 | 1.55 c | 1.80 b | 3.45 a | 0.230 | <0.0001 |
| C18:1 cis-9 | 20.06 c | 26.24 a | 23.20 b | 0.933 | 0.004 |
| Polyunsaturated fatty acids (PUFA) | 5.72 b | 7.01 a | 7.40 a | 0.201 | <0.0001 |
| C18:2 cis-9.cis-12n-6 | 3.99 c | 5.01 a | 4.51 b | 0.118 | <0.0001 |
| C18:2 cis-9. trans-11 CLA | 0.58 c | 0.82 b | 1.30 a | 0.083 | <0.0001 |
| C18:2 trans-10. cis-12 CLA | 0.064 c | 0.128 a | 0.088 b | 0.0102 | <0.0001 |
| C18:3 cis-9. cis-12. cis-15n-3 | 0.640 c | 0.653 b | 1.11 a | 0.048 | <0.0001 |
| C20:4 n-6 | 0.239 a | 0.164 b | 0.170 b | 0.0271 | 0.090 |
| C20:5 n-3 | 0.138 | 0.160 | 0.167 | 0.0160 | 0.208 |
| PUFA/SFA | 0.085 b | 0.116 a | 0.121 a | 0.0043 | <0.0001 |
| n-3 PUFA | 0.845 c | 0.891 b | 1.34 a | 0.051 | <0.0001 |
| n-6 PUFA | 4.23 c | 5.17 a | 4.67 b | 0.118 | <0.0001 |
| n6/n3 | 5.34 ab | 6.04 a | 3.63 b | 0.266 | <0.0001 |
| Short and medium chain fatty acids (SMCFA) 3 | 65.10 a | 55.76 b | 55.77 b | 1.019 | <0.0001 |
| Long chain fatty acids (LCFA) 4 | 34.60 b | 44.07 a | 43.83 a | 1.021 | <0.0001 |
| ∆9 Desaturase index | |||||
| C14:1/(C14:0 + C14:1) | 0.119 ab | 0.122 a | 0.107 b | 0.004 | 0.090 |
| C16:1/(C16:0 + C16:1) | 0.063 | 0.063 | 0.058 | 0.0040 | 0.567 |
| C18:1/(C18:0 + C18:1) | 0.744 ab | 0.741 a | 0.706 b | 0.0212 | 0.053 |
| C18:2 cis-9. trans-11/(C18:1 trans-11 + C18:2 cis-9. trans-11) | 0.289 ab | 0.393 a | 0.268 b | 0.0331 | 0.031 |
| Atherogenicity index 5 | 3.29 a | 2.38 b | 2.42b | 0.163 | <0.0001 |
| Item | Diet 1 | p-Value | ||
|---|---|---|---|---|
| CON (n = 65) | SOY (n = 64) | LIN (n = 65) | ||
| Number of pregnant to first TAI | 15 b | 23 ab | 27 a | 0.07 |
| Number of pregnant | 46 | 48 | 51 | 0.60 |
| Number of TAI 2 Median (interquartile range) | 3 (2) a | 2(3) ab | 2(3) b | 0.08 |
| Open days 3 Median (interquartile range) | 164 (100) | 155 (102) | 123 (89) | 0.29 |
| Item | Diet 1 | Mean | SEM 2 | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| CON | SOY | LIN | D | T | DXT | |||
| Progesterone (ng/mL) | 0.09 | <0.0001 | 0.12 | |||||
| Time 1 | 4.24 | 4.77 | 5.39 | 4.80 | ||||
| Time 2 | 0.67 | 0.77 | 0.73 | 0.73 | ||||
| Time 3 | 4.39 | 4.64 | 5.97 | 5.00 | ||||
| Mean | 3.10 b | 3.40 ab | 4.03 a | 0.311 | ||||
| BHBA (mmol/L) | 0.31 | <0.0001 | 0.30 | |||||
| Time 1 | 0.51 | 0.50 | 0.53 | 0.51 | ||||
| Time 2 | 0.38 | 0.40 | 0.46 | 0.42 | ||||
| Time 3 | 0.52 | 0.50 | 0.52 | 0.51 | ||||
| Mean | 0.47 | 0.47 | 0.50 | 0.018 | ||||
| NEFA (mmol/L) | 0.04 | <0.0001 | 0.45 | |||||
| Time 1 | 0.49 | 0.55 | 0.49 | 0.51 | ||||
| Time 2 | 0.24 | 0.30 | 0.29 | 0.28 | ||||
| Time 3 | 0.20 | 0.24 | 0.27 | 0.24 | ||||
| Mean | 0.31 b | 0.36 a | 0.35a | 0.015 | ||||
| Insulin ((µg/L) | 0.63 | <0.0001 | 0.28 | |||||
| Time 1 | 0.74 | 0.70 | 0.67 | 0.70 | ||||
| Time 2 | 0.49 | 0.53 | 0.50 | 0.50 | ||||
| Time 3 | 0.89 | 0.73. | 0.76 | 0.79 | ||||
| Mean | 0.71 | 0.65 | 0.64 | 0.049 | ||||
| IGF-1 (ng/mL) | 0.002 | <0.0001 | 0.63 | |||||
| Time 1 | 157.47 | 137.45 | 118.8 | 137.91 | ||||
| Time 2 | 159.82 | 155.10 | 135.73 | 150.22 | ||||
| Time 3 | 167.83 | 154.05 | 137.02 | 153.02 | ||||
| Mean | 161.89 a | 149.02 b | 130.60 c | 6.25 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, T.; Martinez, D.; Isabel, B.; Cabezas, A.; Jimeno, V. Vegetable Oils Rich in Polyunsaturated Fatty Acids Supplementation of Dairy Cows’ Diets: Effects on Productive and Reproductive Performance. Animals 2019, 9, 205. https://doi.org/10.3390/ani9050205
Castro T, Martinez D, Isabel B, Cabezas A, Jimeno V. Vegetable Oils Rich in Polyunsaturated Fatty Acids Supplementation of Dairy Cows’ Diets: Effects on Productive and Reproductive Performance. Animals. 2019; 9(5):205. https://doi.org/10.3390/ani9050205
Chicago/Turabian StyleCastro, Teresa, Diego Martinez, Beatriz Isabel, Almudena Cabezas, and Vicente Jimeno. 2019. "Vegetable Oils Rich in Polyunsaturated Fatty Acids Supplementation of Dairy Cows’ Diets: Effects on Productive and Reproductive Performance" Animals 9, no. 5: 205. https://doi.org/10.3390/ani9050205