Efficient Expression of Xylanase by Codon Optimization and Its Effects on the Growth Performance and Carcass Characteristics of Broiler

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Vectors, Reagents, and Culture Medium
2.2. Design and Synthesis of the OptiXynB Gene
2.3. Transformation into P. pastoris
2.4. Expression of optiXynB
2.5. Purification of Recombinant optiXynB
2.6. Detection of optiXynB Activity
2.7. Analysis of optiXynB Properties
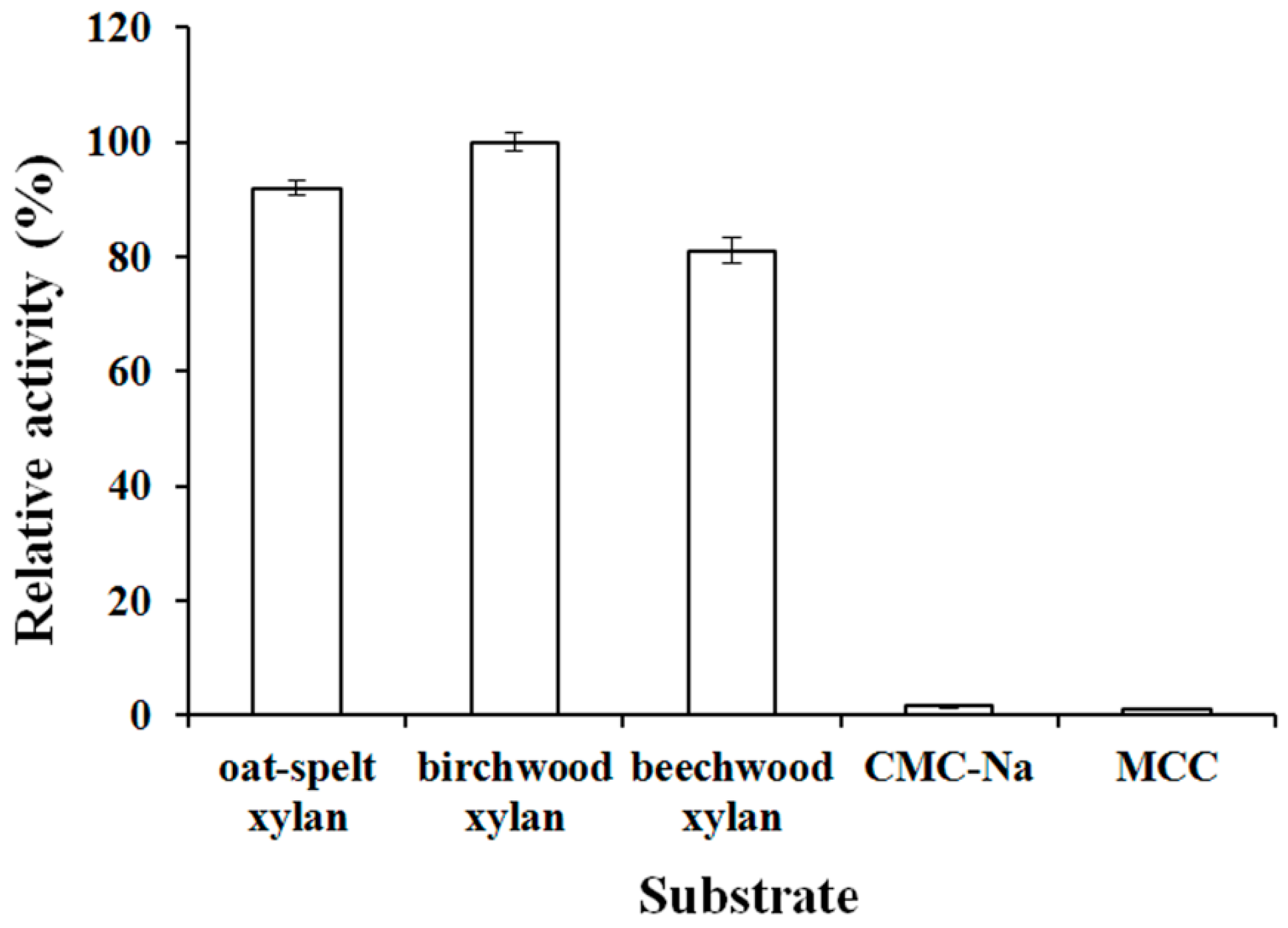
2.8. Analysis of Substrate Specificity
2.9. Broilers, Management, and Diets
2.10. Growth Performance
2.11. Carcass Characteristics
2.12. Statistical Analysis
3. Results
3.1. Design of optiXynB Gene
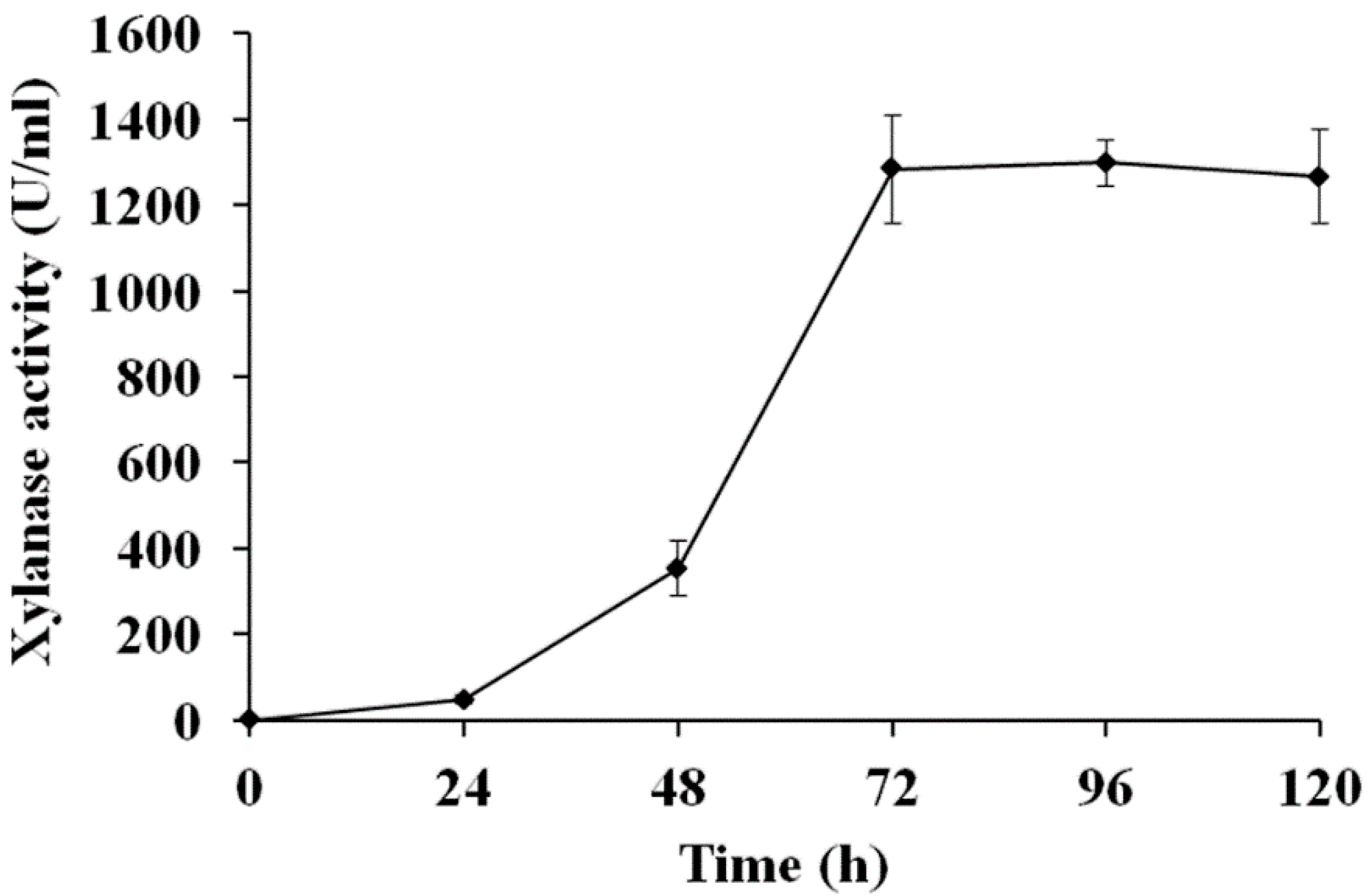
3.2. Expression of optiXynB Gene in P. pastoris
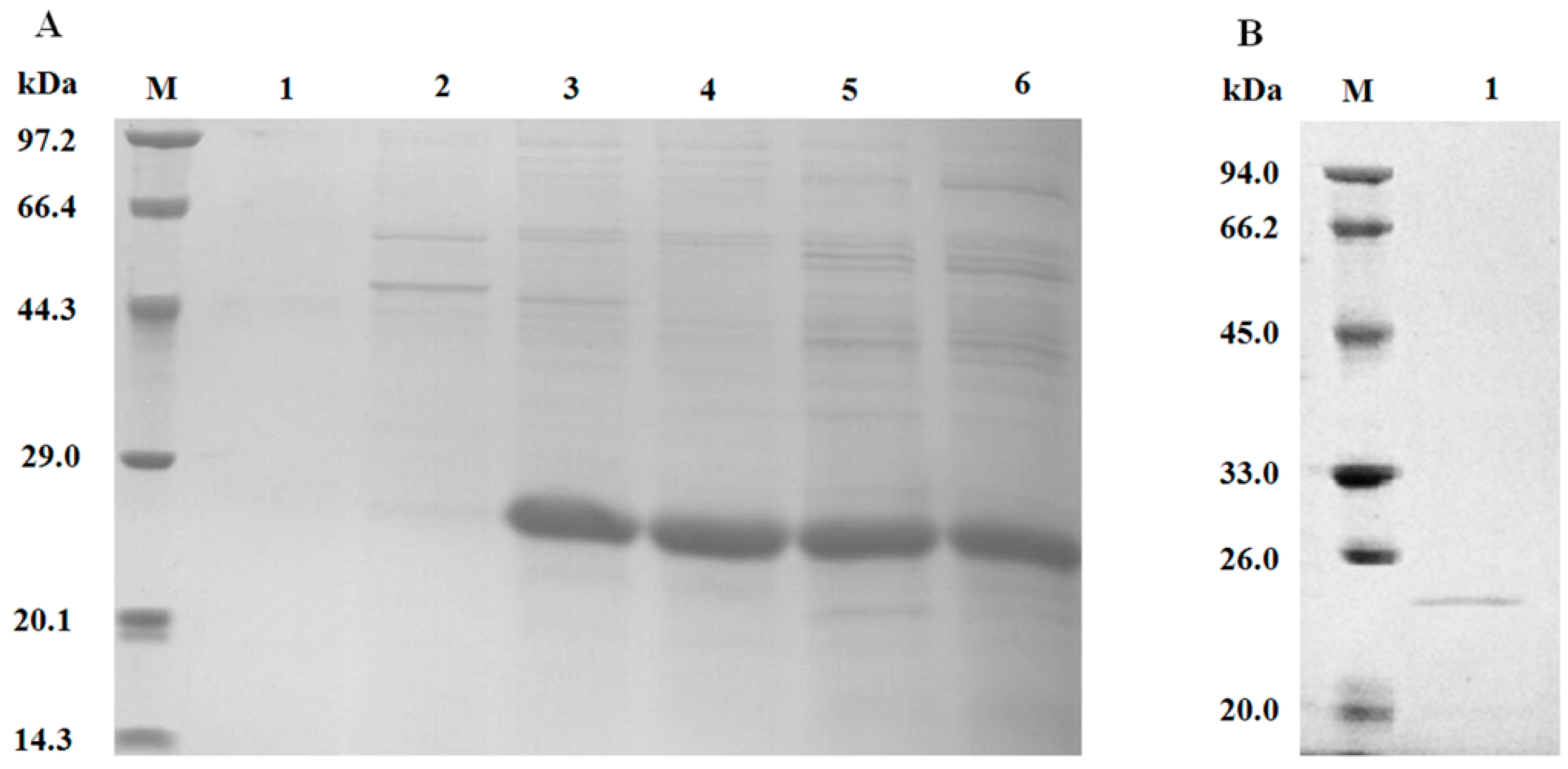
3.3. SDS-PAGE Determination of Crude and Purified optiXynB
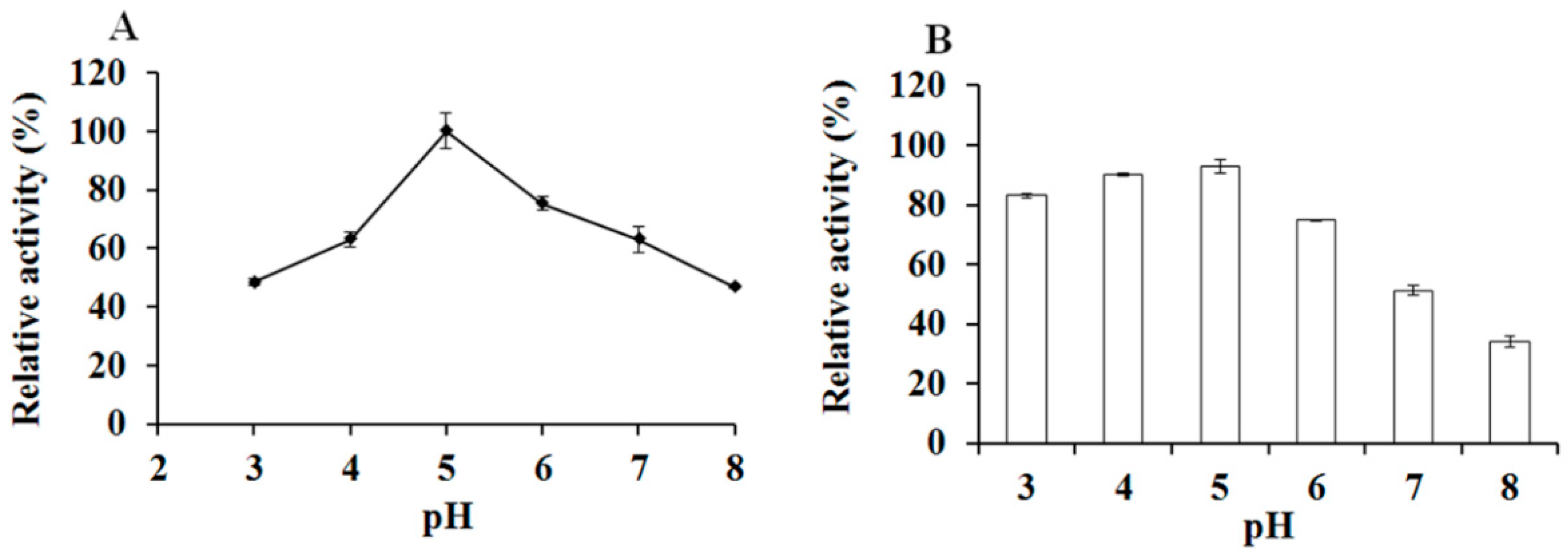
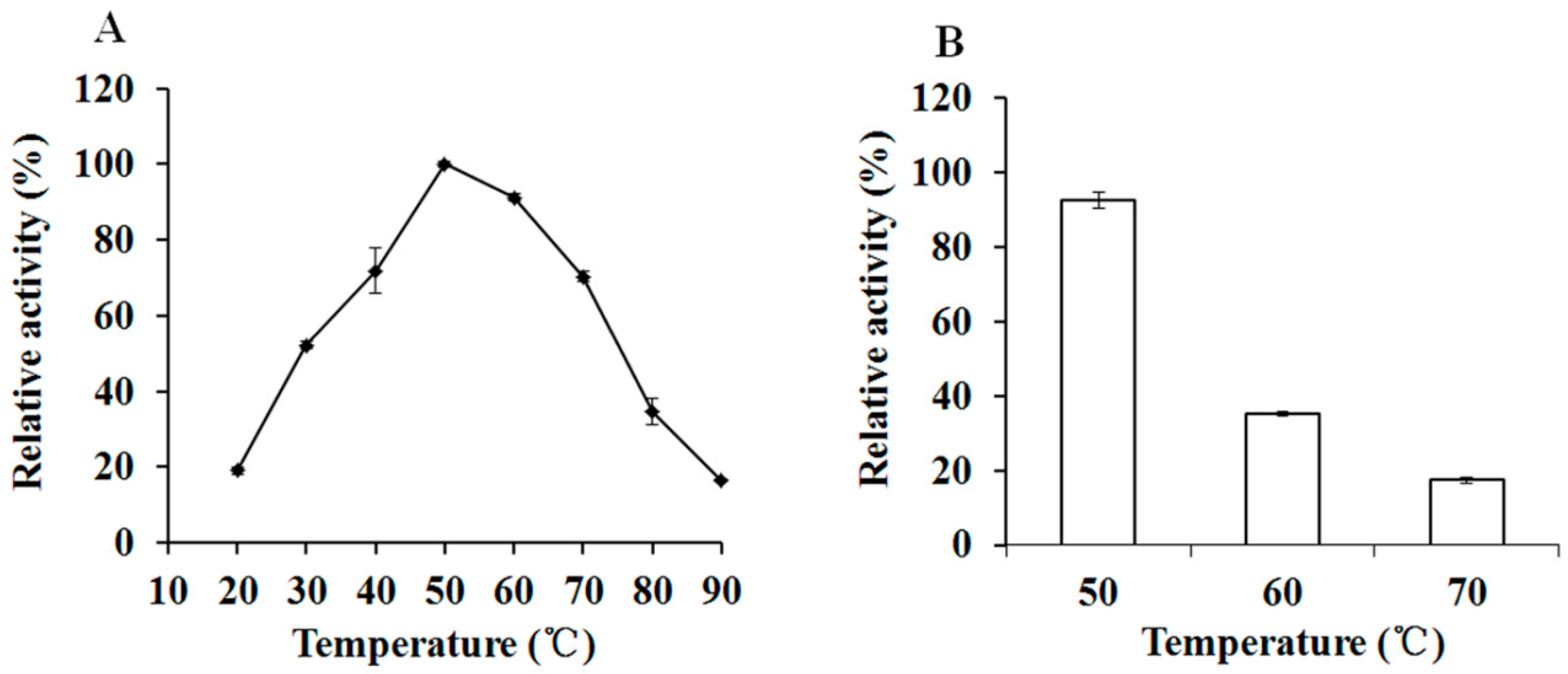
3.4. The optiXynB Properties
3.5. Growth Performance of Broilers
3.6. Carcass Characteristics of Broilers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bueno, R.D.; Borges, L.L.; God, P.I.G.; Piovesan, N.D.; Teixeira, A.I.; Cruz, C.D.; Barros, E.G. Quantification of anti-nutritional factors and their correlations with protein and oil in soybeans. An. Acad. Bras. Cienc. 2018, 90, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ru, Y.J.; Tang, D.F.; Xu, T.S.; Partridge, G.G. Effects of corn distillers dried grains with solubles and xylanase on growth performance and digestibility of diet components in broilers. Anim. Feed Sci. Technol. 2011, 163, 260–266. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial xylanases: Engineering, production and industrial applications. Biotechnol. Adv. 2012, 30, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Kiarie, E.; Romero, L.F.; Ravindran, V. Growth performance, nutrient utilization, and digesta characteristics in broiler chickens fed corn or wheat diets without or with supplemental xylanase. Poult. Sci. 2014, 93, 1186–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowieson, A.J.; Bedford, M.R.; Ravindran, V. Interactions between xylanase and glucanase in maize-soy-based diets for broilers. Brit. Poult. Sci. 2010, 51, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Stricker, A.R.; Mach, R.L.; de Graaff, L.H. Regulation of transcription of cellulases-and hemicellulases-encoding genes in Aspergillus niger and Hypocrea jecorina (Trichoderma reesei). Appl. Microbiol. Biot. 2008, 78, 211–220. [Google Scholar] [CrossRef]
- He, J.; Zhang, K.; Yu, B.; Ding, X.; Chen, D. Expression of a Trichoderma reesei β-xylanase gene in Escherichia coli and the activity of the enzyme on fiber-bond substrates. Protein Expr. Purif. 2009, 67, 1–6. [Google Scholar]
- Spohner, S.C.; Müller, H.; Quitmann, H.; Czermak, P. Expression of enzymes for the usage in food and feed industry with Pichia pastoris. J. Biotechnol. 2015, 202, 118–134. [Google Scholar] [CrossRef]
- Mellitzer, A.; Weis, R.; Glieder, A.; Flicker, K. Expression of lignocellulolytic enzymes in Pichia pastoris. Microb. Cell Fact. 2012, 11, 61. [Google Scholar] [CrossRef]
- Hu, H.; Gao, J.; He, J.; Yu, B.; Zheng, P.; Huang, Z.; Mao, X.; Yu, J.; Han, G.; Chen, D. Codon optimization significantly improves the expression level of a keratinase gene in Pichia pastoris. PLoS ONE 2013, 3, e58393. [Google Scholar] [CrossRef] [PubMed]
- Shumiao, Z.; Huang, J.; Zhang, C.; Deng, L.; Hu, N.; Liang, Y. High-level expression of an Aspergillus niger endo-beta-1, 4-glucanase in Pichia pastoris through gene codon optimization and synthesis. J. Microbiol. Biotechnol. 2010, 20, 467–473. [Google Scholar]
- Gustafsson, C.; Govindarajan, S.; Minshull, J. Codon bias and heterologous protein expression. Trends Biotechnol. 2004, 22, 346–353. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th rev. ed.; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Wen, A.; Bai, X.; Dai, S.; Shah, A.A.; Hu, H. Effect of dietary sodium diacetate on growth performance, carcass characteristics, meat quality, intestinal pH and Escherichia coli of broilers. Anim. Prod. Sci. 2018, 58, 1323–1328. [Google Scholar] [CrossRef]
- Dhiman, S.S.; Sharma, J.; Battan, B. Industrial applications and future prospects of microbial xylanases: A review. BioResources 2008, 3, 1377–1402. [Google Scholar]
- He, J.; Yu, B.; Zhang, K.; Ding, X.; Chen, D. Expression of endo-1, 4-beta-xylanase from Trichoderma reesei in Pichia pastoris and functional characterization of the produced enzyme. BMC Biotechnol. 2009, 9, 56. [Google Scholar] [CrossRef]
- Daly, R.; Hearn, M. Expression of heterologous proteins in Pichia pastoris: A useful experimental tool in protein engineering and production. J. Mol. Recognit. 2005, 18, 119–138. [Google Scholar] [CrossRef]
- Krainer, F.W.; Dietzsch, C.; Hajek, T.; Herwig, C.; Spadiut, O.; Glieder, A. Recombinant protein expression in Pichia pastoris strains with an engineered methanol utilization pathway. Microb. Cell Fact. 2012, 11, 22–35. [Google Scholar] [CrossRef]
- Duan, H.; Wang, H.; Ma, B.; Jiang, P.; Tu, P.; Ni, Z.; Li, X.; Li, M.; Ma, X.; Wang, B.; et al. Codon optimization and expression of irisin in Pichia pastoris GS115. Int. J. Biol. Macromol. 2015, 79, 21–26. [Google Scholar] [CrossRef]
- Jia, H.; Fan, G.; Yan, Q.; Liu, Y.; Yan, Y.; Jiang, Z. High-level expression of a hyperthermostable Thermotoga maritima xylanase in Pichia pastoris by codon optimization. J. Mol. Catal. B-Enzym. 2012, 78, 72–77. [Google Scholar] [CrossRef]
- Yu, P.; Yan, Y.; Gu, Q.; Wang, X. Codon optimisation improves the expression of Trichoderma viride sp. endochitinase in Pichia pastoris. Sci. Rep. 2013, 3, 3043. [Google Scholar] [CrossRef]
- Ata, Ö.; Boy, E.; Güneş, H.; Çalık, P. Codon optimization of xylA gene for recombinant glucose isomerase production in Pichia pastoris and fed-batch feeding strategies to fine-tune bioreactor performance. Bioprocess Biosyst. Eng. 2015, 38, 889–903. [Google Scholar] [CrossRef]
- La Grange, D.C.; Pretorius, I.S.; Van Zyl, W.H. Expression of a Trichoderma reesei beta-xylanase gene (XYN2) in Saccharomyces cerevisiae. Appl. Environ. Microb. 1996, 62, 1036–1044. [Google Scholar]
- Ruanglek, V.; Sriprang, R.; Ratanaphan, N.; Tirawongsaroj, P.; Chantasigh, D.; Tanapongpipat, S.; Pootanakit, K.; Eurwilaichitr, L. Cloning, expression, characterization, and high cell-density production of recombinant endo-1, 4-β-xylanase from Aspergillus niger in Pichia pastoris. Enzym. Microb. Technol. 2007, 41, 19–25. [Google Scholar] [CrossRef]
- Liu, M.Q.; Liu, G.F. Expression of recombinant Bacillus licheniformis xylanse A in Pichia pastoris and xylooligosaccharides released from xylans by it. Protein Expr. Purif. 2008, 57, 101–107. [Google Scholar] [CrossRef]
- Janis, J.; Rouvinen, J.; Leisola, M.; Turunen, O.; Vainiotalo, P. Thermostability of endo-1, 4-β-xylanase II from Trichoderma reesei studied by electrospary ionization Fourier-transform ion cyclotron resonance MS, hydrogen/deuterium-exchange reactions and dynamic light scattering. Biochem. J. 2001, 356, 453–460. [Google Scholar] [CrossRef]
- Irwin, D.; Jung, E.D.; Wilson, D.B. Characterization and sequence of a Thermomonospora fusca xylanase. Appl. Environ. Microbiol. 1994, 60, 763–770. [Google Scholar]
- Zhang, S.Z.; Zhang, K.Y.; Ding, X.M.; Tan, Q. Effects of Xylanase on different protein feeds in vitro. Chin. J. Anim. Nutr. 2007, 19, 719–724. [Google Scholar]
- Bigge, A.E.; Purdum, S.; Hanford, K. Effect of xylanase on the performance of laying hens fed a low energy corn and soybean diet. Poult. Sci. 2018, 9, 3183–3187. [Google Scholar] [CrossRef]
- Cowieson, A.J. Factors that affect the nutritional value of maize for broilers. Anim. Feed Sci. Technol. 2005, 119, 293–305. [Google Scholar] [CrossRef]
- Nian, F.; Guo, Y.M.; Ru, Y.J.; Péron, A.; Li, F.D. Effect of xylanase supplementation on the net energy for production, performance and gut microflora of broilers fed corn/soy-based diet. Asian Austral. J. Anim. 2011, 24, 1282–1287. [Google Scholar] [CrossRef]
- Rajkumar, U.; Muthukumar, M.; Haunshi, S.; Niranjan, M.; Raj, M.V.L.N.; Rama Rao, S.V.; Chatterjee, R.N. Comparative evaluation of carcass traits and meat quality in native Aseel chickens and commercial broilers. Br. Poult. Sci. 2016, 57, 339–347. [Google Scholar] [CrossRef]
- Chen, C.; Han, K.; Xiao, F.; Zhang, W.J. Effects of compound xylanase and celluloses on the growth and slaughter performance of 43~65 days Guangxi partridge chicken. Feed Ind. 2018, 4, 38–41. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Treatment | SEM 1 | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Control | 500 IU/kg | 1000 IU/kg | 1500 IU/kg | Linear 2 | Quadratic 2 | ||
| Final weight (g) | 2022.8a | 2036.9ab | 2047.1b | 2055.9b | 4.26 | 0.003 | 0.711 |
| Body weight gain (g) | 1975.9a | 1990.2ab | 1999.8b | 2009.3b | 4.31 | 0.003 | 0.735 |
| Feed intake (g) | 4340.8 | 4314.5 | 4315.3 | 4305.2 | 15.82 | 0.494 | 0.813 |
| Feed conversion rate (g/g) | 2.20 | 2.17 | 2.16 | 2.14 | 0.01 | 0.033 | 0.667 |
| Item | Treatment | SEM 1 | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Control | 500 IU/kg | 1000 IU/kg | 1500 IU/kg | Linear 2 | Quadratic 2 | ||
| Pre-evisceration weight (g) | 1801.8a | 1831.1b | 1849.8c | 1846.2c | 4.88 | <0.001 | 0.004 |
| Dressed percentage (%) | 89.2a | 90.0b | 90.5b | 90.1b | 0.15 | 0.006 | 0.021 |
| Eviscerated weight (g) | 1339.5a | 1351.4b | 1352.8b | 1353.9b | 1.64 | <0.001 | 0.019 |
| Percentage of eviscerated yield (%) | 66.3 | 66.4 | 66.2 | 66.1 | 0.12 | 0.397 | 0.727 |
| Breast muscle weight | 331.8 | 343.6 | 348.9 | 349.9 | 3.24 | 0.043 | 0.385 |
| Percentage of breast muscle yield (%) | 24.8 | 25.4 | 25.8 | 25.8 | 0.23 | 0.101 | 0.533 |
| Leg muscle weight | 286.8 | 289.9 | 295.2 | 299.4 | 2.35 | 0.049 | 0.903 |
| Percentage of leg muscle yield (%) | 21.4 | 21.5 | 21.8 | 22.1 | 0.16 | 0.109 | 0.675 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, H.; Dai, S.; Wen, A.; Bai, X. Efficient Expression of Xylanase by Codon Optimization and Its Effects on the Growth Performance and Carcass Characteristics of Broiler. Animals 2019, 9, 65. https://doi.org/10.3390/ani9020065
Hu H, Dai S, Wen A, Bai X. Efficient Expression of Xylanase by Codon Optimization and Its Effects on the Growth Performance and Carcass Characteristics of Broiler. Animals. 2019; 9(2):65. https://doi.org/10.3390/ani9020065
Chicago/Turabian StyleHu, Hong, Sifa Dai, Aiyou Wen, and Xi Bai. 2019. "Efficient Expression of Xylanase by Codon Optimization and Its Effects on the Growth Performance and Carcass Characteristics of Broiler" Animals 9, no. 2: 65. https://doi.org/10.3390/ani9020065