Effect of Breed on Transcriptional and Protein Expression of Lipogenic Enzymes in Tail and Subcutaneous Adipose Tissue from Two Grazing Breeds of Lambs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. Lipid Extraction and Fatty Acid Analysis
2.3. qRT-PCR Analysis
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
3.1. Relative mRNA Expression
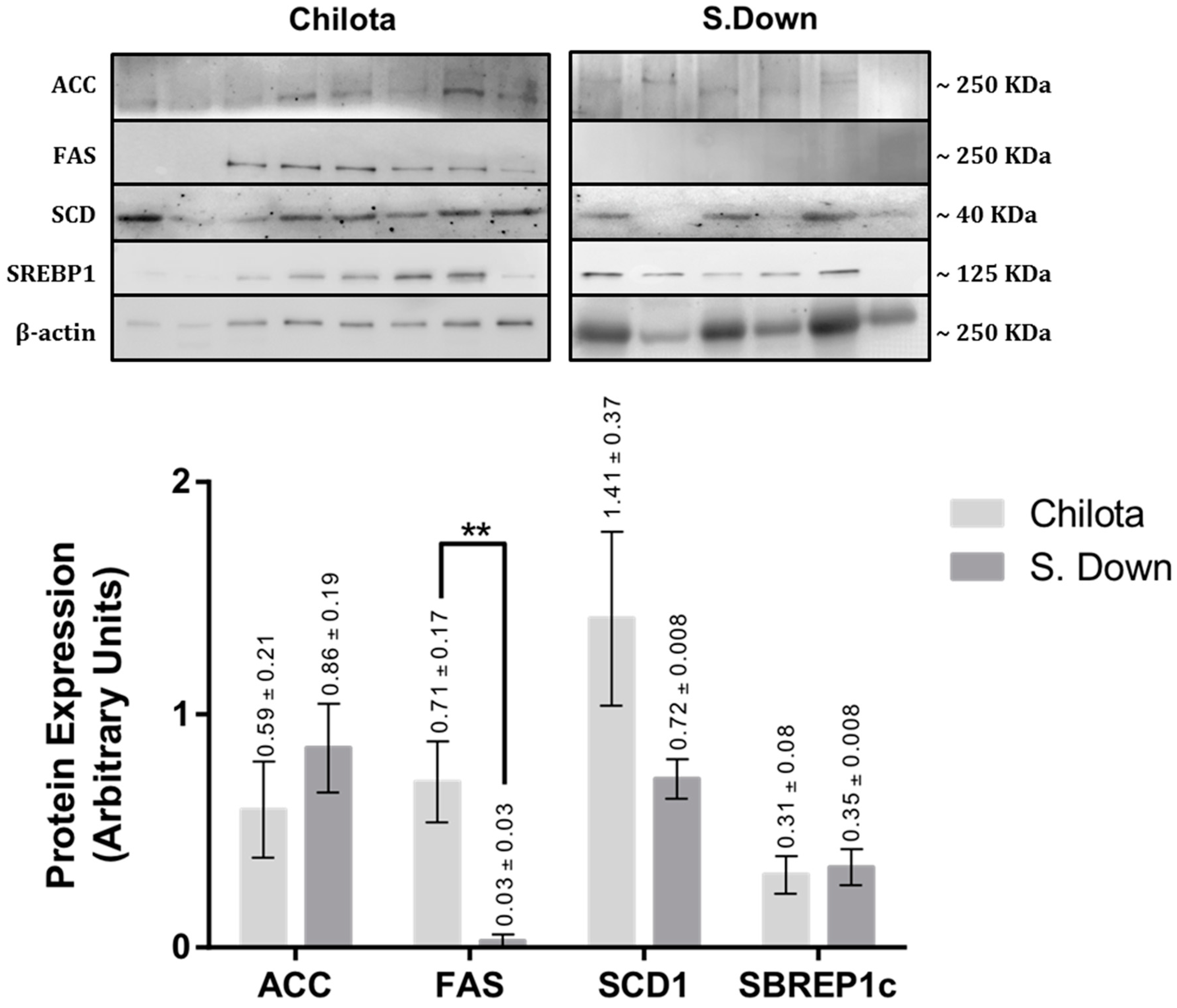
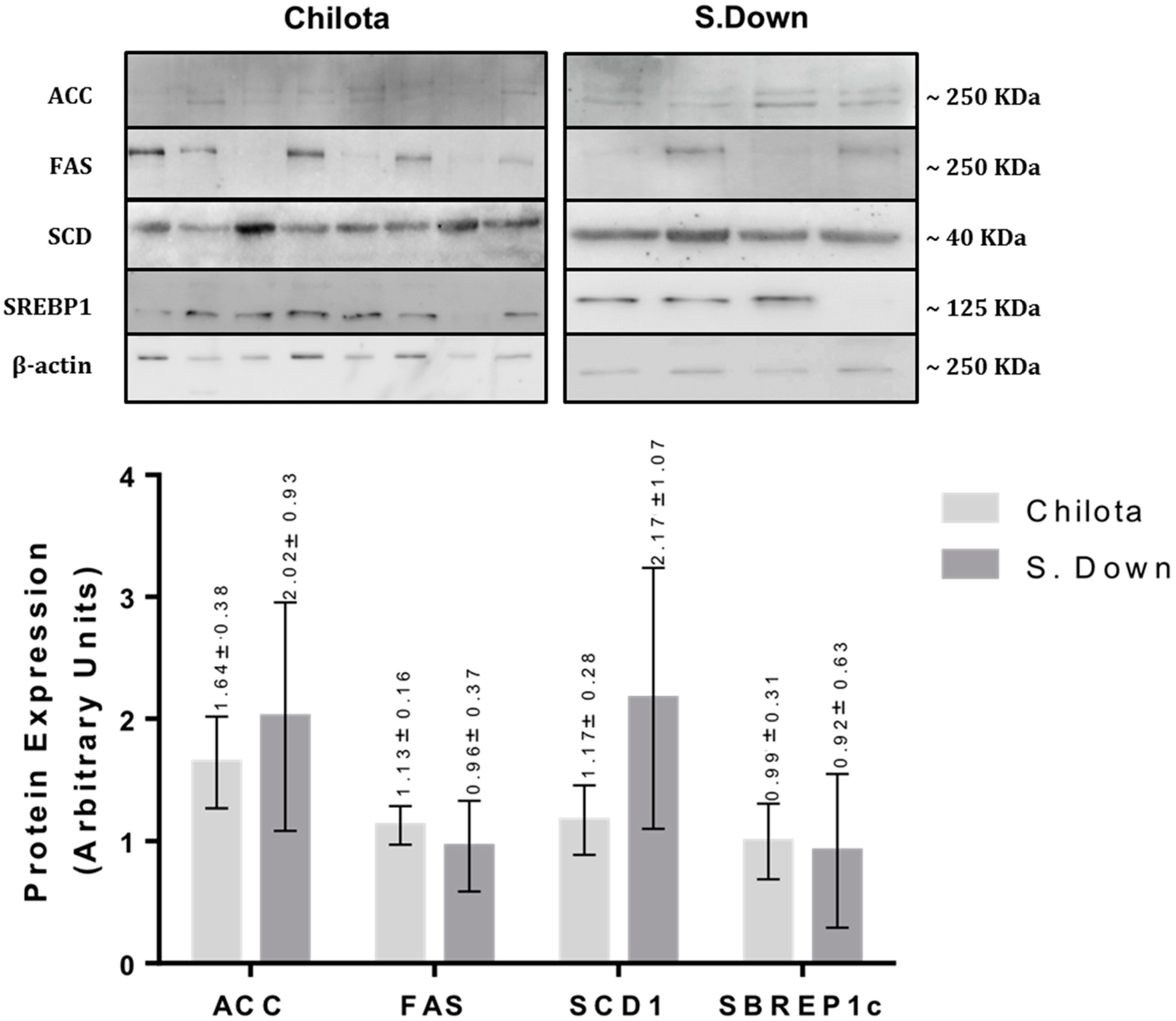
3.2. Protein Expression
3.3. Fatty Acid Composition.
3.3.1. Fatty Acid Composition in Subcutaneous Fat
3.3.2. Fatty Acid Composition in Tail Fat
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De la Barra, R.; Martinez, M.E.; Carvajal, A.M. Genetic relationship between Chilota and Spanish native sheep breeds of Chiloé. J. Livest. Sci. 2012, 3, 79–84. [Google Scholar]
- De la Barra, R.; Uribe, H.; Latorre, E.; San Primitivo, F.; Arranz, J. Genetic structure and diversity of four Chilean sheep breeds. Chilean J. Agric. Res. 2010, 70, 646–651. [Google Scholar] [CrossRef]
- Martínez, M.E.; Calderón, C.; Uribe, H.; De la Barra, R. Effect of management practices in the productive performance of three sheep breeds in the Chiloé Archipelago, Chile. J. Livest. Sci. 2012, 3, 57–66. [Google Scholar]
- De la Barra, R. Efecto de la introducción de la ganadería en el archipiélago de Chiloé, Chile. Doctoral Dissertations, Departamento de Ecología y Departamento de Producción Animal, Universidad de León, León, Spain, 2008. [Google Scholar]
- Scollan, N.; Hocquette, J.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Dance, L.J.E.; Matthews, K.R.; Doran, O. Effect of breed on fatty acid composition and stearoyl-CoA desaturase protein expression in the Semimembranosus muscle and subcutaneous adipose tissue of cattle. Livest. Sci. 2009, 125, 291–297. [Google Scholar] [CrossRef]
- Dannenberger, D.; Nuernberg, G.; Scollan, N.; Schabbel, W.; Steinhart, H.; Ender, K.; Nuernberg, K. Effect of Diet on the Deposition of n-3 Fatty Acids, Conjugated Linoleic and C18:1trans Fatty Acid Isomers in Muscle Lipids of German Holstein Bulls. J. Agric. Food Chem. 2004, 52, 6607–6615. [Google Scholar] [CrossRef] [PubMed]
- Nuernberg, K.; Nuernberg, G.; Ender, K.; Dannenberger, D.; Schabbel, W.; Grumbach, S.; Zupp, W.; Steinhart, H. Effect of grass vs. concentrate feeding on the fatty acid profile of different fat depots in lambs. Eur. J. Lipid. Sci. Technol. 2005, 107, 737–745. [Google Scholar] [CrossRef]
- Nuernberg, K.; Fischer, A.; Nuernberg, G.; Ender, K.; Dannenberger, D. Meat quality and fatty acid composition of lipids in muscle and fatty tissue of Skudde lambs fed grass versus concentrate. Small Rum. Res. 2008, 74, 279–283. [Google Scholar] [CrossRef]
- Popova, T. Effect of the rearing system on the fatty acid composition and oxidative stability of the M. longissimus lumborum and M. semimembranosus in lambs. Small Rum. Res. 2007, 71, 150–157. [Google Scholar] [CrossRef]
- Herdmann, A.; Nuernberg, K.; Martin, J.; Nuernberg, G.; Doran, O. Effect of dietary fatty acids on expression of lipogenic enzymes and fatty acid profile in tissues of bulls. Animal 2010, 4, 755–762. [Google Scholar] [CrossRef]
- Hiller, B.; Herdmann, A.; Nuernberg, K. Dietary n-3 fatty acids significantly suppress lipogenesis in bovine muscle and adipose tissue: A functional genomics approach. Lipids 2011, 46, 557–567. [Google Scholar] [CrossRef]
- Hiller, B.; Hocquette, J.; Cassar-Malek, I.; Nuernberg, G.; Nuernberg, K. Dietary n-3 PUFA affect lipid metabolism and tissue function related genes in bovine muscle. Br. J. Nutr. 2012, 108, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Bakhtiarizadeh, M.R.; Moradi-Shahrbabak, M.; Ebrahimie, E. Underlying functional genomics of fat deposition in adipose tissue. Gene 2013, 521, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Tsiplakou, E.; Flemetakis, E.; Kalloniati, C.; Zervas, G. Differences in mRNA lipogenic gene expression in the subcutaneous adipose tissue of sheep and goats under the same dietary treatments. Small Rumin. Res. 2011, 99, 110–115. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, J.W. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Serrano, C.; Margalida, J.; Serrano, M.; Rodellar, C.; Calvo, J.H. Effect of the feeding system on the fatty acid composition, expression of the Δ9-desaturase, peroxisome proliferator-activated receptor alpha, gamma, and sterol regulatory element binding protein 1 genes in the semitendinous muscle of light lambs of the Rasa Aragonesa breed. BMC Vet. Res. 2010, 6, 40. [Google Scholar] [CrossRef]
- Dervishi, E.; Serrano, C.; Joy, M.; Rodellar, C.; Calvo, J.H. The effect of feeding system in the expression of genes related with fat metabolism in semitendinosus muscle in sheep. Meat Sci. 2011, 89, 91–97. [Google Scholar] [CrossRef]
- Conte, G.; Jeronimo, E.; Serra, A.; Bessa, R.J.B.M.; Mele, M. Effect of dietary polyunsaturated fatty acids on stearoyl CoA-desaturase gene expression in intramuscular lipids of lamb. Ital. J. Anim. Sci. 2012, 11, e79. [Google Scholar] [CrossRef]
- Gallardo, M.A.; Dannenberger, D.; Rivero, M.J.; Pulido, R.; Nürnberg, K. Fatty acid profile of plasma, muscle and adipose tissues in Chilota lambs grazing on two different low quality pasture types in Chiloé Archipelago (Chile). Anim. Sci. J. 2014, 85, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.E.M.; Woodward, B.; Otter, N.; Doran, O. Relationship between the expression of key lipogenic enzymes, fatty acid composition, and intramuscular fat content of Limousin and Aberdeen Angus cattle. Livest. Sci. 2010, 127, 22–29. [Google Scholar] [CrossRef]
- Shirouchi, B.; Albrecht, E.; Nuernberg, G.; Maak, S.; Olavanh, S.; Nakamura, Y.; Sato, M.; Gotoh, T.; Nuernberg, K. Fatty acid profiles and adipogenic gene expression of various fat depots in Japanese Black and Holstein steers. Meat Sci. 2014, 96, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.A.; Cárcamo, J.G.; Hiller, B.; Nuernberg, G.; Nuernberg, K.; Dannenberger, D. Expression of lipid metabolism related genes in subcutaneous adipose tissue from Chilota lambs grazing on two different pasture types. Eur. J. Lipid. Sci. Technol. 2015, 117, 23–30. [Google Scholar] [CrossRef]
- Worgall, T.S.; Sturley, S.L.; Seo, T.; Osborne, T.F.; Deckelbaum, R.J. Polyunsaturated fatty acids decrease expression of promoters with sterol regulatory elements by decreasing levels of mature sterol regulatory element-binding protein. J. Biol. Chem. 1998, 273, 25537–25540. [Google Scholar] [CrossRef]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: transcriptional mediators of lipid homeostasis. Cold Spring Harb. Symp. Quant. Biol. 2002, 67, 491–498. [Google Scholar] [CrossRef]
- Sekiya, M.; Yahagi, N.; Matsuzaka, T.; Najima, Y.; Nakakuki, M.; Nagai, R.; Ishibashi, S.; Osuga, J.; Yamada, N.; Shimano, H. Polyunsaturated fatty acids ameliorate hepatic steatosis in obese mice by SREBP-1 suppression. Hepatology 2003, 38, 1529–1539. [Google Scholar] [CrossRef]
- Kaur, G.; Sinclair, A.J.; Cameron-Smith, D.; Barr, D.P.; Molero-Navajas, J.C.; Konstantopoulos, N. Docosapentaenoic acid (22:5n-3) down-regulates the expression of genes involved in fat synthesis in liver cells. Prostaglandins Leukot. Essent. Fatty Acids 2011, 85, 155–161. [Google Scholar] [CrossRef]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef]
- Castro-Carrera, T.; Frutos, P.; Leroux, C.; Chilliard, Y.; Hervás, G.; Belenguer, A.; Bernard, L.; Toral, P.G. Dietary sunflower oil modulates milk fatty acid composition without major changes in adipose and mammary tissue fatty acid profile or related gene mRNA abundance in sheep. Animal 2014, 9, 582–591. [Google Scholar] [CrossRef]
- Kashani, A.; Behrens Holman, B.W.; Nichols, P.D.; Malau-Aduli, A.E.O. Effect of dietary supplementation with Spirulina on the expressions of AANAT, ADRB3, BTG2 and FASN genes in he subcutaneous adipose and Longissimus dorsi muscle tissues of purebred and crossbreed Australian sheep. J. Anim. Sci. Technol. 2015, 57, 8. [Google Scholar] [CrossRef] [PubMed]
- Angulo, J.; Hiller, B.; Albrecht, E.; Olivera, M.; Mahecha, L.; Nuernberg, G.; Dannenberger, D.; Nuernberg, K. Effect of different dietary fats on protein expression of sterol regulatory element-binding protein 1 (SREBP-1) in mammary gland tissue of lactating cows. Livest. Sci. 2012, 143, 300–304. [Google Scholar] [CrossRef]
- Mauvoisin, D.; Mounier, C. Hormonal and nutritional regulation of SCD1 gene expression. Biochimie 2011, 93, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Flowers, M.T.; Ntambi, J.M. Stearoyl-CoA desaturase and its relation to high carbohydrate diets and obesity. Biochem. Biophys. Acta 2009, 1791, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.T.; Schinckel, A.P.; Weber, T.E.; Mersmann, H.J. Expression of porcine transcription factors and genes related to fatty acid metabolism in different tissues and genetic populations. J. Anim. Sci. 2000, 78, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.Z.; Yang, F.; Zhou, A.G. Study of regulation of exogenous recombinant somatotropin on fat metabolism in different cross-finishing pigs. Acta Vet. Zootech. Sin. 2001, 32, 1–4. [Google Scholar]
- Bernard, L.; Leroux, C.; Chilliard, Y. Expression and nutritional regulation of lipogenic genes in the ruminant lactating mammary gland. Adv. Exp. Med. Biol. 2008, 606, 67–108. [Google Scholar] [CrossRef]
- Larsson, O.; Nadon, R. Gene expression-time to change point of view? Biotechnol. Genet. Eng. Rev. 2008, 25, 77–92. [Google Scholar] [CrossRef]
- Xu, X.; Wei, X.; Yang, Y.; Niu, W.; Kou, Q.; Wang, X.; Chen, Y. mRNA transcription and protein expression of PPARγ, FAS, and HSL in different parts of the carcass between fat-tailed and thin-tailed sheep. Electron. J. Biotechnol. 2015, 18, 215–220. [Google Scholar] [CrossRef]
- Maleki, E.; Kafilzadeh, F.; Meng, G.Y.; Rajion, M.A.; Ebrahim, M. The effect of breed on fatty acid composition of subcutaneous adipose tissues in fat-tailed sheep under identical feeding conditions. S. Afr. J. Anim. Sci. 2015, 45, 12–19. [Google Scholar] [CrossRef]
- Ropka-Molik, K.; Knapik, J.; Pieszka, M.; Szmatola, T. The expression of the SCD1 gene and its correlation with fattening and carcass traits in sheep. Arch. Anim. Breed 2016, 59, 37–43. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Forward Primer Sequence | Accession Number | Amplicon |
|---|---|---|---|
| Reverse Primer Sequence | Length | ||
| ACC | F: ATGTGGCCTGGGTAGATCCT | NM_001009256.1 | 261 bp |
| R: ACGTAACCAGGCTGATGGTG | |||
| FAS | F: GGAAGGCGGGACTATATGGC | XM_004013447.1 | 278 bp |
| R: CATGCTGTAGCCTACGAGGG | |||
| SCD1 | F: GGCGTTCCAGAATGACGTTT | NM_001009254.1 | 251 bp |
| R: TGAAGCACAACAGCAGGACA | |||
| SREBP1 | F: GTCTACCACAAGCTGCACCAG | XM_004013336.1 | 216 bp |
| R: GCTCAGGAAGAAGCGTGTCA | |||
| β-Actin | F: TGAAGTGTGACGTGGACATCCGTA | NM_001009784.1 | 108 bp |
| R: AGGTGATCTCCTTCTGCATCCTGT |
| Breed | Subcutaneous Fat (SCF) | p | Tail Fat (TF) | p | ||
|---|---|---|---|---|---|---|
| Chilota | Suffolk Down | Chilota | Suffolk Down | |||
| Enzymes (FC) | LSM 1 ± SEM | LSM 1 ± SEM | LSM 1 ± SEM | LSM 1 ± SEM | ||
| ACC | 1.75 ± 0.78 | 1.50 ± 0.35 | 0.78 | 3.83 ± 1.71 | 0.64 ± 0.11 | 0.09 |
| FAS | 0.92 ± 0.29 | 1.49 ± 0.24 | 0.16 | 2.24 ± 0.70 | 0.70 ± 0.19 | 0.06 |
| SCD1 | 1.07 ± 0.32 | 1.25 ± 0.17 | 0.63 | 1.66 ± 0.50 | 0.79 ± 0.08 | 0.12 |
| SREBP1c | 1.08 ± 0.34 | 1.50 ± 0.28 | 0.36 | 2.37 ± 0.77 | 0.76 ± 0.19 | 0.06 |
| Breed | Subcutaneous Fat (SCF) | Tail Fat (TF) | ||||
|---|---|---|---|---|---|---|
| Suffolk Down | Diff 1 | p | Suffolk Down | Diff 2 | p | |
| Fatty Acids (%) | LSM ± SEM (n = 6) | LSM ± SEM (n = 6) | ||||
| Fat content (%) | 40.65 ± 3.03 | −2.82 | 0.01 | 67.57 ± 3.12 | −4.49 | 0.01 |
| 14:0 | 6.77 ± 0.29 | −1.38 | <0.01 | 7.04 ± 0.32 | −0.63 | 0.09 |
| 16:0 | 23.40 ± 0.49 | −3.43 | 0.01 | 24.14 ± 0.54 | −0.70 | 0.27 |
| 16:1 | 1.79 ± 0.18 | +2.25 | <0.01 | 2.17 ± 0.12 | +0.03 | 0.86 |
| 18:0 | 20.64 ± 1.50 | −8.15 | 0.01 | 20.90 ± 1.15 | +0.76 | 0.63 |
| 18:1cis-9 | 38.39 ± 2.06 | +9.80 | 0.01 | 36.41 ± 1.62 | −0.06 | 0.98 |
| 18:2n-6 | 1.72 ± 0.11 | −0.06 | 0.85 | 1.64 ± 0.05 | +0.26 | <0.01 |
| 18:3n-3 | 0.97 ± 0.04 | +0.13 | 0.09 | 1.08 ± 0.04 | +0.26 | <0.01 |
| CLAcis-9, trans-11 | 1.72 ± 0.39 | +1.29 | 0.01 | 2.34 ± 0.15 | −0.11 | 0.57 |
| 20:2n-6 | 0.03 ± 0.01 | 0.00 | 0.89 | 0.03 ± 0.01 | 0.00 | 0.35 |
| 20:3n-3 | 0.01 ± 0.00 | 0.00 | 0.92 | 0.01 ± 0.00 | 0.01 | 0.06 |
| 20:4n-6 | 0.25 ± 0.13 | −0.15 | 0.21 | 0.14 ± 0.03 | −0.06 | 0.06 |
| 20:5n-3 | 0.05 ± 0.01 | +0.01 | 0.62 | 0.05 ± 0.01 | −0.01 | 0.74 |
| 22:6n-3 | 0.09 ± 0.05 | −0.04 | 0.39 | 0.05 ± 0.02 | −0.01 | 0.20 |
| Sum SFA † | 53.67 ± 2.09 | −13.57 | <0.01 | 54.94 ± 1.69 | −0.35 | 0.88 |
| Sum MUFA ‡ | 41.23 ± 2.13 | +12.46 | 0.01 | 39.44 ± 1.64 | −0.02 | 0.99 |
| Sum PUFA § | 5.10 ± 0.39 | +1.11 | 0.03 | 5.62 ± 0.23 | +0.37 | 0.20 |
| Sum n-6 PUFA ¶ | 2.17 ± 0.24 | −0.30 | 0.36 | 1.98 ± 0.04 | +0.26 | 0.01 |
| Sum n-3 PUFA ∥ | 1.12 ± 0.09 | +0.10 | 0.40 | 1.20 ± 0.05 | +0.23 | 0.09 |
| n-6/n-3PUFA ratio | 2.07 ± 0.40 | −0.52 | 0.25 | 1.65 ± 0.05 | −0.09 | 0.30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallardo, M.; Arias-Darraz, L.; Cárcamo, J. Effect of Breed on Transcriptional and Protein Expression of Lipogenic Enzymes in Tail and Subcutaneous Adipose Tissue from Two Grazing Breeds of Lambs. Animals 2019, 9, 64. https://doi.org/10.3390/ani9020064
Gallardo M, Arias-Darraz L, Cárcamo J. Effect of Breed on Transcriptional and Protein Expression of Lipogenic Enzymes in Tail and Subcutaneous Adipose Tissue from Two Grazing Breeds of Lambs. Animals. 2019; 9(2):64. https://doi.org/10.3390/ani9020064
Chicago/Turabian StyleGallardo, María, Luis Arias-Darraz, and Juan Cárcamo. 2019. "Effect of Breed on Transcriptional and Protein Expression of Lipogenic Enzymes in Tail and Subcutaneous Adipose Tissue from Two Grazing Breeds of Lambs" Animals 9, no. 2: 64. https://doi.org/10.3390/ani9020064