Population Structure and Genetic Diversity of Italian Beef Breeds as a Tool for Planning Conservation and Selection Strategies

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data
2.2. Pedigree Analysis
2.3. Genetic Diversity
2.4. Principal Component Analysis
3. Results
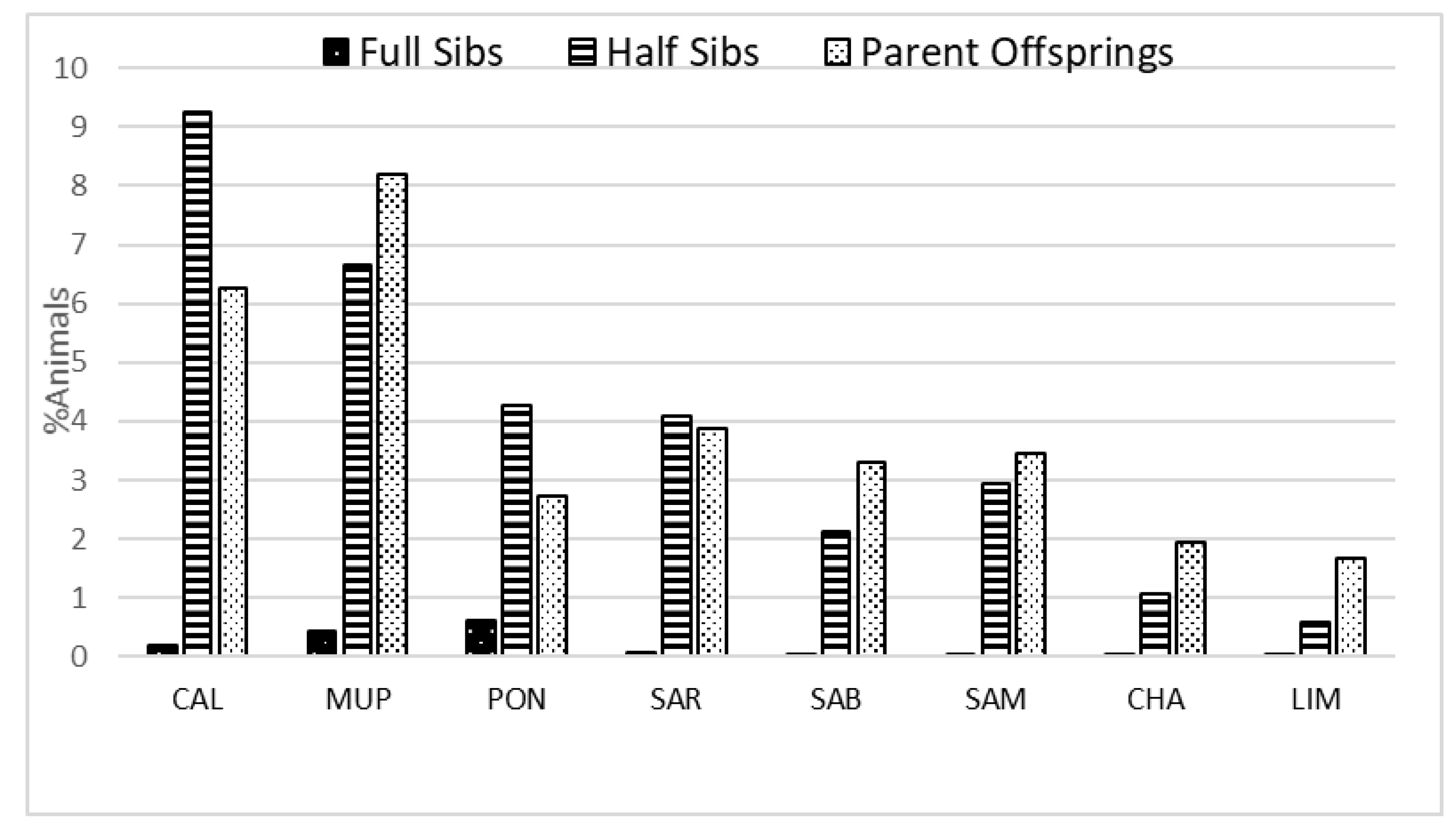
3.1. Pedigree Analysis
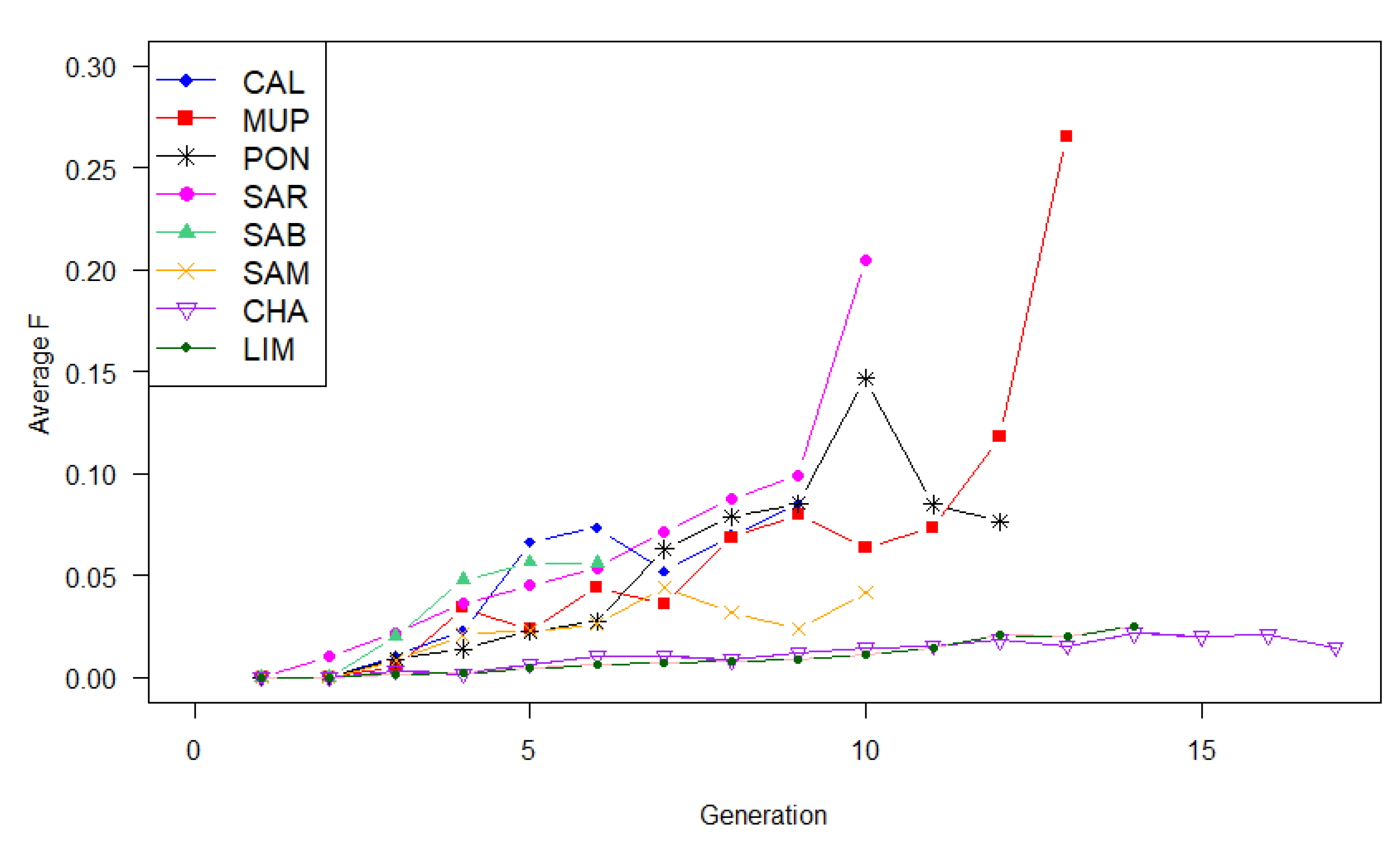
3.2. Genetic Diversity
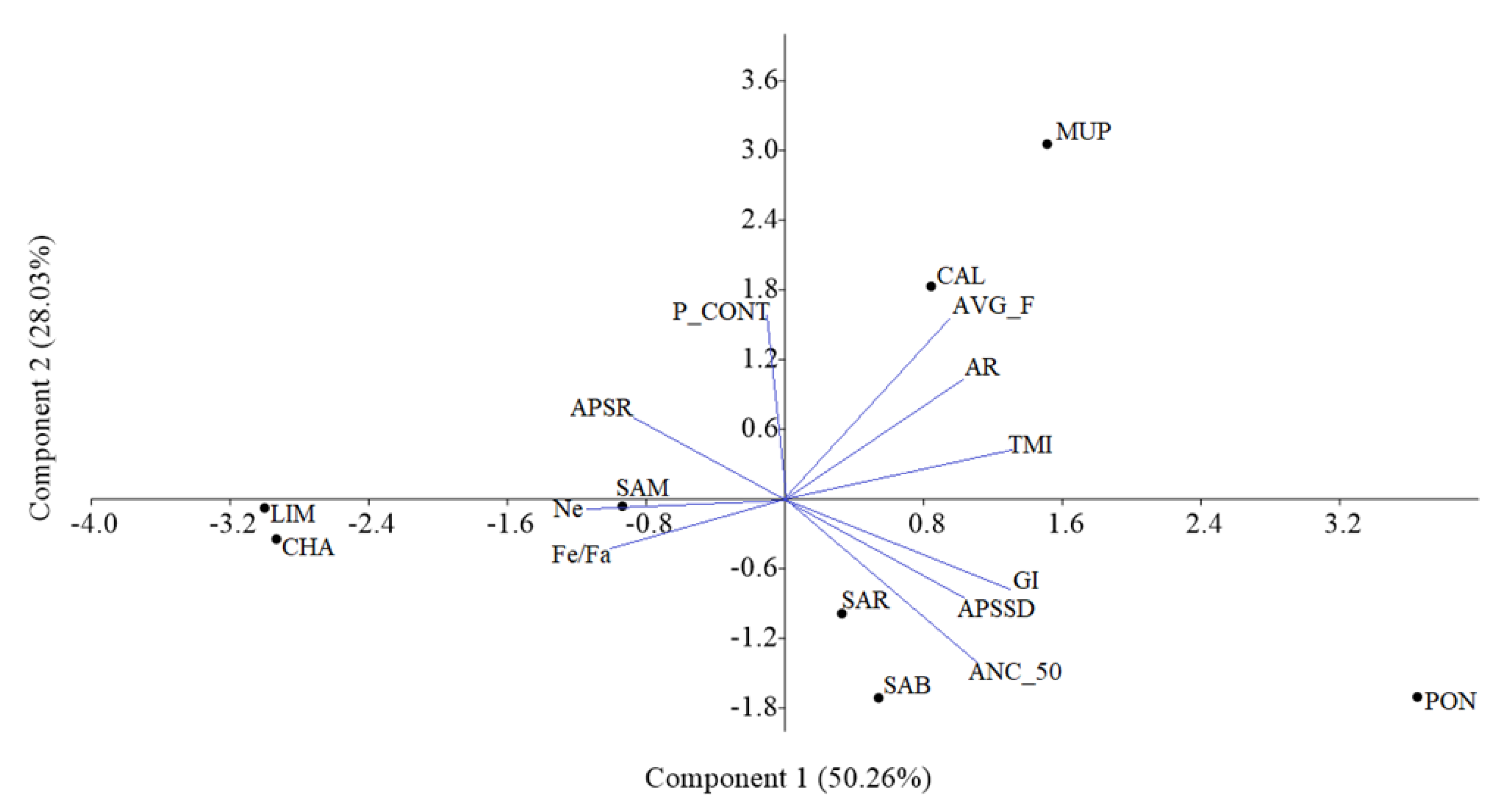
3.3. PCA of the Population Structure Parameters
4. Discussion
4.1. Pedigree Analysis
4.2. Genetic Diversity
4.3. Measures of Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Larson, G.; Bürger, J. A population genetics view of animal domestication. Trends Genet. 2013, 29, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Scheu, A.; Powell, A.; Bollongino, R.; Vigne, J.-D.; Tresset, A.; Çakırlar, C.; Benecke, N.; Bürger, J. The genetic prehistory of domesticated cattle from their origin to the spread across Europe. BMC Genet. 2015, 16, 54. [Google Scholar] [CrossRef] [PubMed]
- Bollongino, R.; Burger, J.; Powell, A.; Mashkour, M.; Vigne, J.-D.; Thomas, M.G. Modern Taurine Cattle Descended from Small Number of Near-Eastern Founders. Mol. Biol. Evol. 2012, 29, 2101–2104. [Google Scholar] [CrossRef] [PubMed]
- Hiemstra, S.J. Local Cattle Breeds in Europe: Development of Policies and Strategies for Self-Sustaining Breeds; Academic Publishing: Wageningen, The Netherlands, 2010; ISBN 978-90-8686-144-6. [Google Scholar]
- Hickey, J.M.; Chiurugwi, T.; Mackay, I.; Powell, W. Implementing Genomic Selection in CGIAR Breeding Programs Workshop Participants. Genomic prediction unifies animal and plant breeding programs to form platforms for biological discovery. Nat. Genet. 2017, 49, 1297–1303. [Google Scholar] [CrossRef]
- Anderson, S. Animal genetic resources and sustainable livelihoods. Ecol. Econ. 2003, 45, 331–339. [Google Scholar] [CrossRef]
- Wollny, C.B. The need to conserve farm animal genetic resources in Africa: Should policy makers be concerned? Ecol. Econ. 2003, 45, 341–351. [Google Scholar] [CrossRef]
- Ovaska, U.; Soini, K. Local Breeds—Rural Heritage or New Market Opportunities? Colliding Views on the Conservation and Sustainable Use of Landraces. Sociol. Rural. 2017, 57, 709–729. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Nelson, G.C.; Rosegrant, M.W.; Palazzo, A.; Gray, I.; Ingersoll, C.; Robertson, R.; Tokgoz, S.; Zhu, T.; Sulser, T.B.; Ringler, C.; et al. Food Security, Farming, and Climate Change to 2050: Scenarios, Results, Policy Options; International Food Policy Research Institute: Washington, DC, USA, 2010; ISBN 978-0-89629-186-7. [Google Scholar]
- The Economist Intelligence Unit. Global Food Security Index 2018; Economist Intelligence Unit: London, UK, 2018. [Google Scholar]
- Risk Status of Animal Genetic Resources|Domestic Animal Diversity Information System (DAD-IS)|Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/dad-is/risk-status-of-animal-genetic-resources/en/ (accessed on 16 September 2019).
- Domestic Animal Diversity Information System (DAD-IS) Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/dad-is/en/ (accessed on 16 September 2019).
- Mendelsohn, R. The challenge of conserving indigenous domesticated animals. Ecol. Econ. 2003, 45, 501–510. [Google Scholar] [CrossRef]
- Curone, G.; Filipe, J.; Cremonesi, P.; Trevisi, E.; Amadori, M.; Pollera, C.; Castiglioni, B.; Turin, L.; Tedde, V.; Vigo, D.; et al. What we have lost: Mastitis resistance in Holstein Friesians and in a local cattle breed. Res. Veter Sci. 2018, 116, 88–98. [Google Scholar] [CrossRef]
- Oldenbroek, K. Utilisation and Conservation of Farm Animal Genetic Resources; Wageningen Academic Publisher: Wageningen, The Netherlands, 2007; ISBN 978-90-8686-032-6. [Google Scholar]
- Hill, W.G.; Mackay, T.F.C. Falconer and Introduction to quantitative genetics. Genetics 2004, 167, 1529–1536. [Google Scholar] [PubMed]
- Sumreddee, P.; Toghiani, S.; Hay, E.H.; Roberts, A.; Agrrey, S.E.; Rekaya, R. Inbreeding depression in line 1 Hereford cattle population using pedigree and genomic information1. J. Anim. Sci. 2019, 97, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Santana, M.; Ayres, D.; Bignardi, A.; Menezes, G.; Silva, L.; Machado, C.; Josahkian, L.; Albuquerque, L. Inbreeding depression in Zebu cattle traits. J. Anim. Breed. Genet. 2016, 133, 523–533. [Google Scholar] [CrossRef]
- Charlesworth, B. Effective population size and patterns of molecular evolution and variation. Nat. Rev. Genet. 2009, 10, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, S.; Ciani, E.; Marsan, P.A.; Bagnato, A.; Battaglini, L.; Bozzi, R.; Carta, A.; Catillo, G.; Cassandro, M.; Casu, S.; et al. Conservation status and historical relatedness of Italian cattle breeds. Genet. Sel. Evol. 2018, 50, 35. [Google Scholar] [CrossRef] [PubMed]
- Bouquet, A.; Venot, E.; Laloë, D.; Forabosco, F.; Fogh, A.; Pabiou, T.; Moore, K.; Eriksson, J.-A.; Renand, G.; Phocas, F. Genetic structure of the European Charolais and Limousin cattle metapopulations using pedigree analyses. J. Anim. Sci. 2011, 89, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, M.L.; Neto, J.B.; Brito, F.V.; Campos, L.T.; Bértoli, C.D.; Campos, G.S.; Cobuci, J.A.; McManus, C.M.; Barcellos, J.O.J.; Gama, L.T. Origins and genetic diversity of British cattle breeds in Brazil assessed by pedigree analyses1. J. Anim. Sci. 2014, 92, 1920–1930. [Google Scholar] [CrossRef]
- Cavani, L.; Silva, R.M.D.O.; Carreño, L.O.D.; Ono, R.K.; Bertipaglia, T.S.; Farah, M.M.; Millen, D.D.; Fonseca, R.D. Genetic diversity of Brazilian Brahman cattle by pedigree analysis. Pesqui. Agropecuária Bras. 2018, 53, 74–79. [Google Scholar] [CrossRef]
- Gutierrez, J.; Goyache, F. A note on ENDOG: A computer program for analysing pedigree information. J. Anim. Breed. Genet. 2005, 122, 172–176. [Google Scholar] [CrossRef]
- Maignel, L.; Boichard, D.; Verrier, E. Genetic variability of French dairy breeds estimated from pedigree information. Interbull Bull. 1996, 14, 49–56. [Google Scholar]
- Kadlečík, O.; Pavlík, I.; Moravčíková, N.; Kasarda, R. Inbreeding and genetic diversity loss of four cattle beef breeds in Slovakia. Acta Fytotech. Zootech. 2016, 19, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Maccluer, J.W.; Boyce, A.J.; Dyke, B.; Pfenning, D.W.; Parsons, C.J.; Weitkamp, L.R. Inbreeding and pedigree structure in Standardbred horses. J. Hered. 1983, 74, 394–399. [Google Scholar] [CrossRef]
- Meuwissen, T.; Luo, Z. Computing inbreeding coefficients in large populations. Genet. Sel. Evol. 1992, 24, 305–313. [Google Scholar] [CrossRef]
- Boichard, D.; Maignel, L.; Verrier, E. The value of using probabilities of gene origin to measure genetic variability in a population. Genet. Sel. Evol. 1997, 29, 5–23. [Google Scholar] [CrossRef]
- Abegaz, F.; Chaichoompu, K.; Génin, E.; Fardo, D.W.; König, I.R.; Mahachie John, J.M.; Van Steen, K. Principals about principal components in statistical genetics. Brief. Bioinform. 2018, bby081. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Cassell, B.; Adamec, V.; Pearson, R. Effect of Incomplete Pedigrees on Estimates of Inbreeding and Inbreeding Depression for Days to First Service and Summit Milk Yield in Holsteins and Jerseys. J. Dairy Sci. 2003, 86, 2967–2976. [Google Scholar] [CrossRef]
- Lutaaya, E.; Misztal, I.; Bertrand, J.; Mabry, J. Inbreeding in populations with incomplete pedigrees. J. Anim. Breed. Genet. 2001, 116, 475–480. [Google Scholar] [CrossRef]
- Cappelloni, M. Sviluppo di Metodi per la Gestione di Razze Autoctone Italiane. Ph.D. Thesis, Università degli Studi di Padova, Padova, Italy, 2013. [Google Scholar]
- Gutiérrez, J.P.; Altarriba, J.; Díaz, C.; Quintanilla, R.; Cañón, J.; Piedrafita, J. Pedigree analysis of eight Spanish beef cattle breeds. Genet. Sel. Evol. 2003, 35, 43–63. [Google Scholar] [CrossRef]
- Cañas-Álvarez, J.J.; Gónzalez-Rodríguez, A.; Martín-Collado, D.; Avilés, C.; Altarriba, J.; Baro, J.A.; De La Fuente, L.F.; Díaz, C.; Molina, A.; Varona, L.; et al. Monitoring changes in the demographic and genealogical structure of the main Spanish local beef breeds1. J. Anim. Sci. 2014, 92, 4364–4374. [Google Scholar] [CrossRef]
- Torrecillas, C.P.; Bozzi, R.; Negrini, R.; Filippini, F.; Giorgetti, A. Genetic variability of three Italian cattle breeds determined by parameters based on probabilities of gene origin. J. Anim. Breed. Genet. 2002, 119, 274–279. [Google Scholar] [CrossRef]
- Falleiro, V.B.; Malhado, C.H.M.; Malhado, A.C.M.; Carneiro, P.L.S.; Carrillo, J.A.; Song, J. Population Structure and Genetic Variability of Angus and Nellore Herds. J. Agric. Sci. 2014, 6, 276. [Google Scholar] [CrossRef]
- Mc Parland, S.; Kearney, J.F.; Rath, M.; Berry, D. Inbreeding trends and pedigree analysis of Irish dairy and beef cattle populations. J. Anim. Sci. 2007, 85, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Cañón, J.; Gutiérrez, J.; Dunner, S.; Goyache, F.; Vallejo, M. Herdbook analyses of the Asturiana beef cattle breeds. Genet. Sel. Evol. 1994, 26, 65–75. [Google Scholar] [CrossRef]
- Herron, N.; Pattie, W. Studies of the Australian Illawarra Shorthorn breed of dairy cattle. II.* Genetic analysis. Aust. J. Agric. Res. 1977, 28, 1119–1132. [Google Scholar] [CrossRef]
- Ozkütük, K.; Bichard, M. Studies of pedigree Hereford cattle breeding. 1. Herdbook analyses. Anim. Sci. 1977, 24, 1–13. [Google Scholar] [CrossRef]
- Gengler, N.; Misztal, I.; Bertrand, J.K.; Culbertson, M.S. Estimation of the dominance variance for postweaning gain in the U.S. Limousin population. J. Anim. Sci. 1998, 76, 2515. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E. Operation of Conservation Schemes. In Genebanks and the Management of Farm Animal Genetic Resources; DLO Institute for Animal Science and Health: Lelystad, The Netherlands, 1999. [Google Scholar]
- Bozzi, R.; Franci, O.; Forabosco, F.; Pugliese, C.; Crovetti, A.; Filippini, F. Genetic variability in three Italian beef cattle breeds derived from pedigree information. Ital. J. Anim. Sci. 2010, 5, 129–137. [Google Scholar] [CrossRef]
- FAO. Secondary Guidelines for Development of National Farm Animal Genetic Resources Management Plans—Management of Small Populations at Risk; FAO: Rome, Italy, 1998. [Google Scholar]
- Melka, M.; Stachowicz, K.; Miglior, F.; Schenkel, F. Analyses of genetic diversity in five Canadian dairy breeds using pedigree data. J. Anim. Breed. Genet. 2013, 130, 476–486. [Google Scholar] [CrossRef]
- Howard, J.T.; Pryce, J.E.; Baes, C.; Maltecca, C. Invited review: Inbreeding in the genomics era: Inbreeding, inbreeding depression, and management of genomic variability. J. Dairy Sci. 2017, 100, 6009–6024. [Google Scholar] [CrossRef]
- Biscarini, F.; Nicolazzi, E.L.; Stella, A.; Boettcher, P.J.; Gandini, G. Challenges and opportunities in genetic improvement of local livestock breeds. Front. Genet. 2015, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Caballero, A.; Santiago, E.; Toro, M.A. Systems of mating to reduce inbreeding in selected populations. Anim. Sci. 1996, 62, 431–442. [Google Scholar] [CrossRef]
- Woolliams, J.; Berg, P.; Dagnachew, B.; Meuwissen, T. Genetic contributions and their optimization. J. Anim. Breed. Genet. 2015, 132, 89–99. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Breed 1 | N | Males | Females | Generations |
|---|---|---|---|---|
| Tuscan | ||||
| CAL | 2798 | 1201 | 1597 | 10 |
| MUP | 3399 | 1447 | 1952 | 14 |
| PON | 328 | 147 | 181 | 13 |
| Sardinian | ||||
| SAR | 97,163 | 28,869 | 68,294 | 11 |
| SAB | 74,981 | 13,697 | 61,284 | 10 |
| SAM | 25,355 | 10,398 | 14,957 | 12 |
| Cosmopolitan | ||||
| CHA | 99,464 | 39,171 | 60,293 | 18 |
| LIM | 322,321 | 133,445 | 188,876 | 15 |
| Breed 1 | EquiGen 2 | FullGen 3 | MaxGen 4 | PCI (%) 5 |
|---|---|---|---|---|
| Tuscan | ||||
| CAL | 2.87 | 2.04 | 4.44 | 66 |
| MUP | 3.91 | 2.44 | 7.55 | 74 |
| PON | 2.10 | 1.06 | 4.24 | 38 |
| Sardinian | ||||
| SAR | 1.10 | 0.64 | 1.89 | 22 |
| SAB | 0.75 | 0.45 | 1.20 | 15 |
| SAM | 1.85 | 1.08 | 3.18 | 39 |
| Cosmopolitan | ||||
| CHA | 2.79 | 1.51 | 6.39 | 50 |
| LIM | 3.07 | 1.79 | 5.83 | 59 |
| Pathway | CAL | MUP | PON | SAR | SAB | SAM | CHA | LIM |
|---|---|---|---|---|---|---|---|---|
| Father to sons | 9.55(13.83) | 9.26(11.33) | 13.20(14.01) | 11.22(14.79) | 6.17(7.69) | 7.26(11.61) | 6.35(7.46) | 7.25(10.11) |
| Father to daughters | 10.37(14.17) | 9.86(11.04) | 17.12(17.59) | 10.66(14.24) | 7.24(8.70) | 7.35(11.45) | 5.46(4.72) | 6.22(6.89) |
| Mother to sons | 10.54(13.30) | 6.82(7.32) | 8.10(10.25) | 8.57(8.47) | 10.86(12.68) | 7.88(6.47) | 7.62(8.80) | 9.13(12.55) |
| Mother to daughters | 10.27(12.77) | 8.20(9.78) | 9.85(9.95) | 10.69(12.68) | 16.49(18.96) | 8.19(8.28) | 7.78(8.11) | 7.52(8.04) |
| Total Interval | 10.29(13.49) | 8.94(10.38) | 12.51(13.86) | 10.60(13.19) | 13.30(16.72) | 7.80(9.54) | 6.69(7.00) | 7.05(8.13) |
| Breed 1 | Ne2 | ΔF (%) 3 | F (%) 4 | TMI (%) 5 | AR (%) 6 |
|---|---|---|---|---|---|
| Tuscan | |||||
| CAL | 19.68 | 2.54 | 5.10 | 6.00 | 6.39 |
| MUP | 18.52 | 2.70 | 7.25 | 8.00 | 10.54 |
| PON | 14.62 | 3.42 | 3.64 | 5.60 | 7.15 |
| Sardinian | |||||
| SAR | 16.64 | 3.00 | 1.90 | 5.10 | 0.04 |
| SAB | 18.91 | 2.64 | 1.23 | 5.10 | 0.05 |
| SAM | 39.79 | 1.26 | 1.66 | 2.80 | 0.37 |
| Cosmopolitan | |||||
| CHA | 90.29 | 0.55 | 0.96 | 1.30 | 0.20 |
| LIM | 132.65 | 0.37 | 0.71 | 0.90 | 0.20 |
| Breed 1 | fe/fa2 | ANC_50 3 | P_CONT (%) 4 | APSR 5 | APSSD 6 |
|---|---|---|---|---|---|
| Tuscan | |||||
| CAL | 1.1 | 8 | 92 | 1.40 | 14.34 |
| MUP | 1.1 | 5 | 94 | −1.42 | 27.57 |
| PON | 1.1 | 5 | 73 | −103.23 | 454.11 |
| Sardinian | |||||
| SAR | 1.2 | 542 | 61 | −4.03 | 21.85 |
| SAB | 1.2 | 294 | 48 | 2.26 | 20.08 |
| SAM | 1.2 | 96 | 78 | −4.38 | 9.50 |
| Cosmopolitan | |||||
| CHA | 3.0 | 219 | 77 | 3.60 | 7.05 |
| LIM | 2.1 | 330 | 83 | 6.04 | 6.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabbri, M.C.; Gonçalves de Rezende, M.P.; Dadousis, C.; Biffani, S.; Negrini, R.; Souza Carneiro, P.L.; Bozzi, R. Population Structure and Genetic Diversity of Italian Beef Breeds as a Tool for Planning Conservation and Selection Strategies. Animals 2019, 9, 880. https://doi.org/10.3390/ani9110880
Fabbri MC, Gonçalves de Rezende MP, Dadousis C, Biffani S, Negrini R, Souza Carneiro PL, Bozzi R. Population Structure and Genetic Diversity of Italian Beef Breeds as a Tool for Planning Conservation and Selection Strategies. Animals. 2019; 9(11):880. https://doi.org/10.3390/ani9110880
Chicago/Turabian StyleFabbri, Maria Chiara, Marcos Paulo Gonçalves de Rezende, Christos Dadousis, Stefano Biffani, Riccardo Negrini, Paulo Luiz Souza Carneiro, and Riccardo Bozzi. 2019. "Population Structure and Genetic Diversity of Italian Beef Breeds as a Tool for Planning Conservation and Selection Strategies" Animals 9, no. 11: 880. https://doi.org/10.3390/ani9110880