
Effects of Dietary Supplementation of Bile Acids on Growth, Glucose Metabolism, and Intestinal Health of Spotted Seabass (Lateolabrax maculatus)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Preparation of Experimental Diets
2.3. Experimental Design and Feeding Trial
2.4. Sample Collection
2.5. Proximate Composition of Diets and Whole Fish
2.6. Glucose Metabolism of Liver
2.7. Intestinal Antioxidant Capacity and Digestive Enzyme Activity
2.8. Histological Analyses and Immunofluorescence Detection in the Intestine
2.9. RNA Extraction and Quantitative Real-Time PCR (qPCR) Assay
2.10. Statistical Analyses
3. Results
3.1. Growth Performance and Proximate Composition Analyses
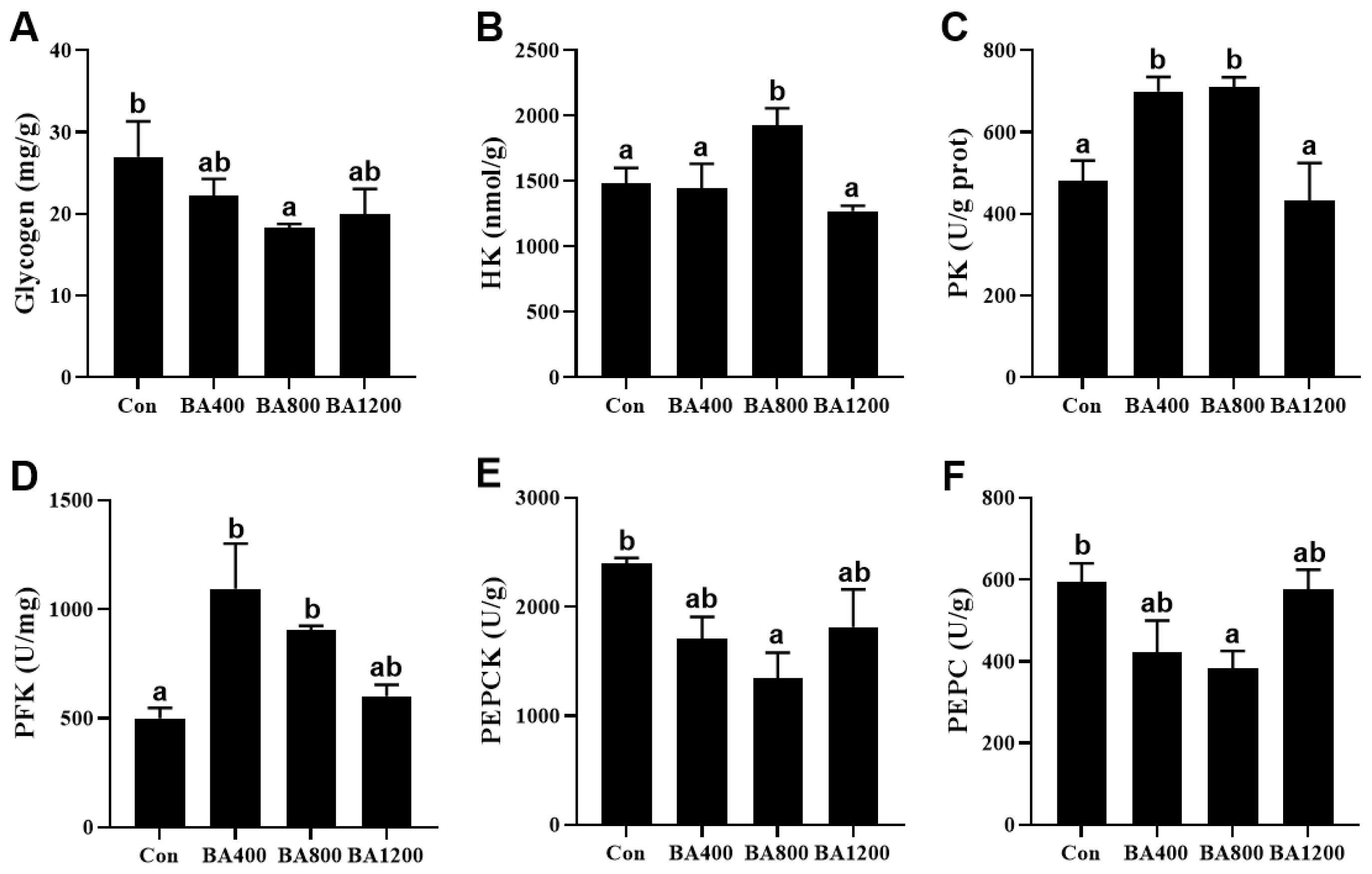
3.2. Glucose Metabolism of Liver
3.3. Histological and Morphological Analyses in the Intestine
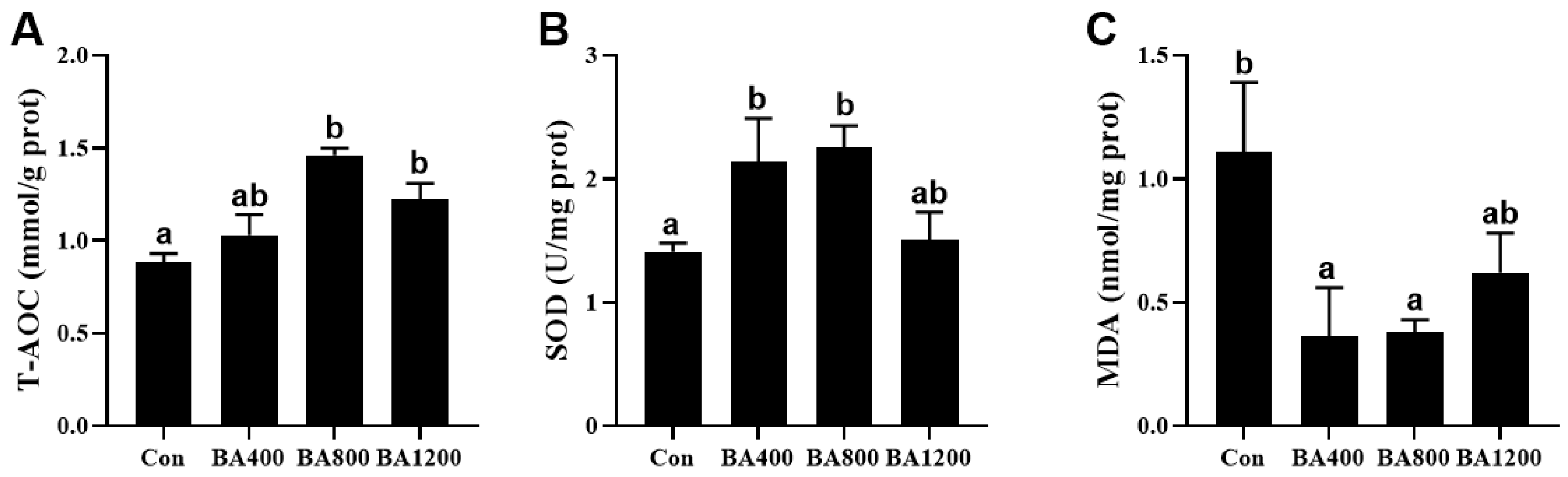
3.4. Antioxidant Capacity in the Intestine
3.5. Digestive Enzyme Activities in the Intestine
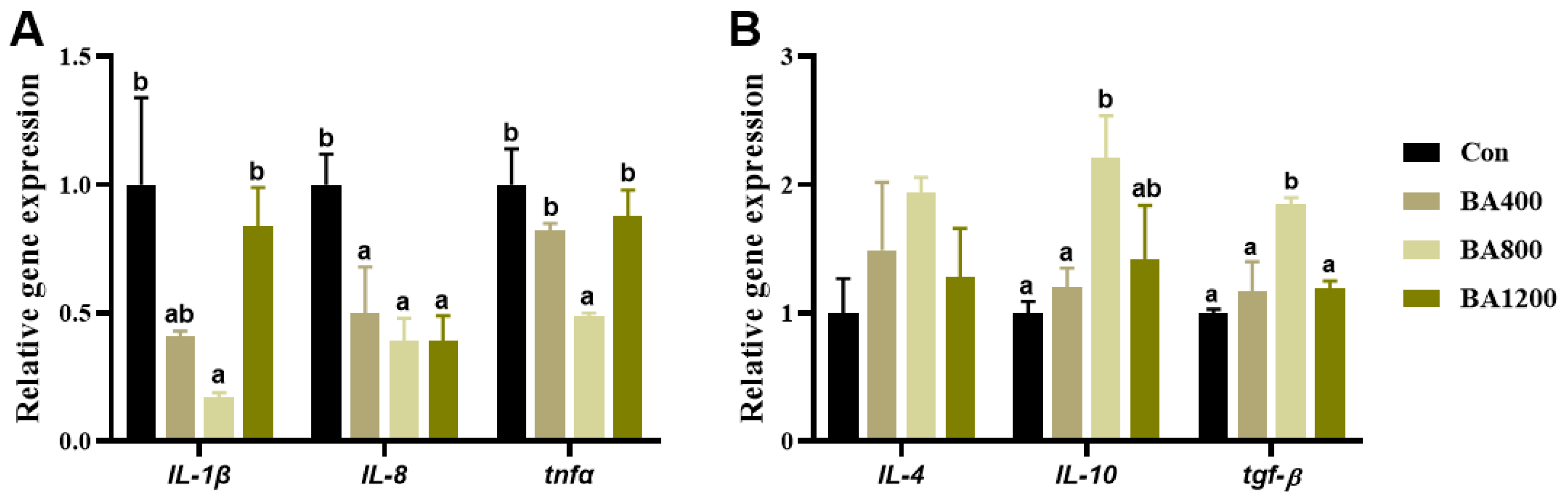
3.6. Immunofluorescence and Transcriptional Expression of Genes Related to Inflammatory Cytokines
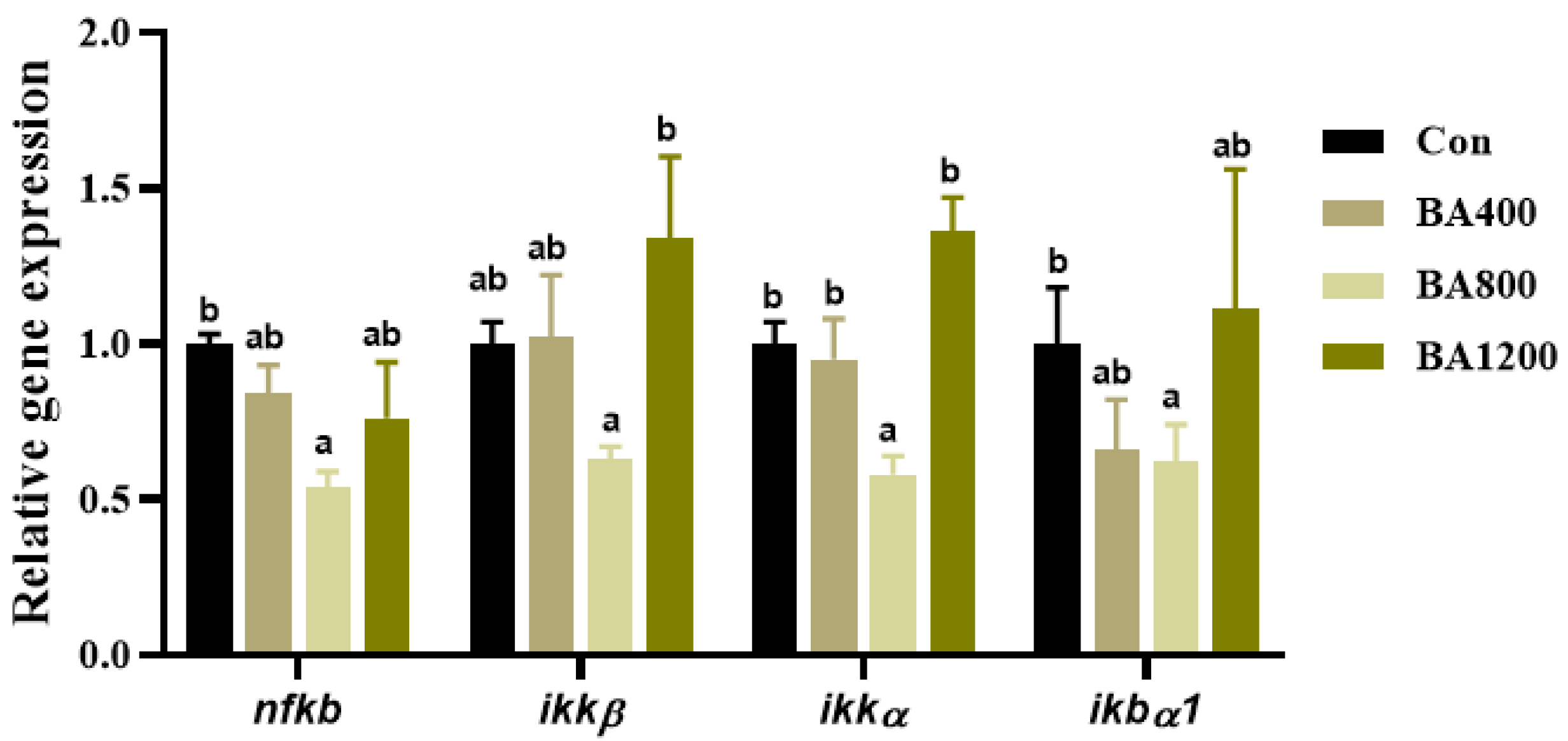
3.7. Transcriptional Expression of Genes Related to NF-κB Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cai, L.; Wang, L.; Song, K.; Lu, K.; Zhang, C.; Rahimnejad, S. Evaluation of protein requirement of spotted seabass (Lateolabrax maculatus) under two temperatures, and the liver transcriptome response to thermal stress. Aquaculture 2020, 516, 734615. [Google Scholar] [CrossRef]
- Ministry of Agriculture of the People’s Republic of China. China fishery statistical yearbook. In Department of Fishery of the Ministry of Agriculture; China Agricultural Press: Beijing, China, 2023; pp. 50–51. [Google Scholar]
- Wen, H.S.; Li, J.F.; Zhang, M.Z.; Li, Y.; Qi, X.; Zhang, K.Q. Physiology and Breeding Technology of Seabass in Mariculture; China Agricultural Press: Beijing, China, 2019; pp. 183–186. [Google Scholar]
- Buentello, J.A.; Gatlin, D.M.; Neill, W.H. Effects of water temperature and dissolved oxygen on daily feed consumption, feed utilization and growth of channel catfish (Ictalurus punctatus). Aquaculture 2000, 182, 339–352. [Google Scholar] [CrossRef]
- Zak, M.A.; Manzon, R.G. Expression and activity of lipid and oxidative metabolism enzymes following elevated temperature exposure and thyroid hormone manipulation in juvenile lake whitefish (Coregonus clupeaformis). Gen. Comp. Endocrinol. 2019, 275, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, Z.; Quan, J.; Lu, J.; Zhao, G.; Sun, J. Dietary nanoselenium supplementation for heat-stressed rainbow trout: Effects on organizational structure, lipid changes, and biochemical parameters as well as heat-shock-protein- and selenoprotein-related gene expression. Fish Physiol. Biochem. 2022, 48, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Kameda, M.; Shoji, Y.; Hayashi, S.; Yamaguchi, T.; Sato, M. Effect of severe environmental thermal stress on redox state in salmon. Redox Biol. 2014, 2, 772–776. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Bacchetta, C.; Cazenave, J. Effect of thermal stress on metabolic and oxidative stress biomarkers of Hoplosternum littorale (Teleostei, Callichthyidae). Ecol. Indic. 2017, 79, 361–370. [Google Scholar] [CrossRef]
- Zhao, H.; Ke, H.; Zhang, L.; Zhao, Z.; Lai, J.; Zhou, J.; Huang, Z.; Li, H.; Du, J.; Li, Q. Integrated analysis about the effects of heat stress on physiological responses and energy metabolism in Gymnocypris chilianensis. Sci. Total Environ. 2022, 806, 151252. [Google Scholar] [CrossRef]
- Kim, B.S.; Jung, S.J.; Choi, Y.J.; Kim, N.N.; Choi, C.Y.; Kim, J.W. Effects of different light wavelengths from LEDs on oxidative stress and apoptosis in olive flounder (Paralichthys olivaceus) at high water temperatures. Fish Shellfish Immun. 2016, 55, 460–468. [Google Scholar] [CrossRef]
- Matthews, K.R.; Berg, N.H. Rainbow trout responses to water temperature and dissolved oxygen stress in two southern California stream pools. J. Fish Biol. 1997, 50, 50–67. [Google Scholar] [CrossRef]
- Vinagre, C.; Madeira, D.; Narciso, L.; Cabral, H.N.; Diniz, M. Effect of temperature on oxidative stress in fish: Lipid peroxidation and catalase activity in the muscle of juvenile seabass, Dicentrarchus labrax. Ecol. Indic. 2012, 23, 274–279. [Google Scholar] [CrossRef]
- Yang, X.; Weng, Q.; Li, X.; Lu, K.; Wang, L.; Song, K.; Zhang, C.; Rahimnejad, S. High water temperature raised the requirements of methionine for spotted seabass (Lateolabrax maculatus). Fish Physiol. Biochem. 2022, 50, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, X.; Song, K.; Li, X.; Zhang, C.; Lu, K.; Wang, L. Effects of dietary zinc levels on growth, intestinal health, antioxidant and non-specific immunity of spotted seabass (Lateolabrax maculatus) reared at two temperatures. Aquaculture 2023, 577, 739959. [Google Scholar] [CrossRef]
- Song, M.; Zhao, J.; Wen, H.; Li, Y.; Li, J.; Li, L.; Tao, Y.; Soengas, J.L. The impact of acute thermal stress on the metabolome of the black rockfish (Sebastes schlegelii). PLoS ONE 2019, 14, e217133. [Google Scholar] [CrossRef] [PubMed]
- Adam, A.H.; Verdegem, M.; Soliman, A.A.; Zaki, M.; Khalil, R.H.; Nour, A.M.; Khaled, A.A.; El Basuini, M.F.; Khalil, H.S. Effect of dietary bile acids: Growth performance, immune response, genes expression of fatty acid metabolism, intestinal, and liver morphology of striped catfish (Pangasianodon hypophthalmus). Aquac. Rep. 2023, 29, 101510. [Google Scholar] [CrossRef]
- Zhou, J.S.; Chen, H.J.; Ji, H.; Shi, X.C.; Li, X.X.; Chen, L.Q.; Du, Z.Y.; Yu, H.B. Effect of dietary bile acids on growth, body composition, lipid metabolism and microbiota in grass carp (Ctenopharyngodon idella). Aquac. Nutr. 2018, 24, 802–813. [Google Scholar] [CrossRef]
- Romano, N.; Kumar, V.; Yang, G.; Kajbaf, K.; Rubio, M.B.; Overturf, K.; Brezas, A.; Hardy, R. Bile acid metabolism in fish: Disturbances caused by fishmeal alternatives and some mitigating effects from dietary bile inclusions. Rev. Aquac. 2020, 12, 1792–1817. [Google Scholar] [CrossRef]
- Li, X.; Shi, M.; Chen, L.; Zhang, S.; Chi, S.; Dong, X.; Deng, J.; Tan, B.; Xie, S. Effects of bile acids supplemented into low fishmeal diet on growth, molting, and intestinal health of Pacific white shrimp, Litopenaeus vannamei. Aquac. Rep. 2023, 29, 101491. [Google Scholar] [CrossRef]
- Wang, L.; Sagada, G.; Wang, C.; Liu, R.; Li, Q.; Zhang, C.; Yan, Y. Exogenous bile acids regulate energy metabolism and improve the health condition of farmed fish. Aquaculture 2023, 562, 738852. [Google Scholar] [CrossRef]
- Yin, P.; Xie, S.; Zhuang, Z.; He, X.; Tang, X.; Tian, L.; Liu, Y.; Niu, J. Dietary supplementation of bile acid attenuate adverse effects of high-fat diet on growth performance, antioxidant ability, lipid accumulation and intestinal health in juvenile largemouth bass (Micropterus salmoides). Aquaculture 2021, 531, 735864. [Google Scholar] [CrossRef]
- Liu, C.; Zhan, S.; Li, N.; Tu, T.; Lin, J.; Li, M.; Chen, M.; Zeng, Z.; Zhuang, X. Bile acid alterations associated with indolent course of inflammatory bowel disease. Scand. J. Gastroenterol. 2023, 58, 988–997. [Google Scholar] [CrossRef]
- Sun, R.; Xu, C.; Feng, B.; Gao, X.; Liu, Z. Critical roles of bile acids in regulating intestinal mucosal immune responses. Ther. Adv. Gastroenter. 2021, 14, 1088201074. [Google Scholar] [CrossRef]
- Lin, S.; Wang, S.; Wang, P.; Tang, C.; Wang, Z.; Chen, L.; Luo, G.; Chen, H.; Liu, Y.; Feng, B.; et al. Bile acids and their receptors in regulation of gut health and diseases. Prog. Lipid Res. 2023, 89, 101210. [Google Scholar] [CrossRef]
- Ding, T.; Xu, N.; Liu, Y.; Du, J.; Xiang, X.; Xu, D.; Liu, Q.; Yin, Z.; Li, J.; Mai, K.; et al. Effect of dietary bile acid (BA) on the growth performance, body composition, antioxidant responses and expression of lipid metabolism-related genes of juvenile large yellow croaker (Larimichthys crocea) fed high-lipid diets. Aquaculture 2020, 518, 734768. [Google Scholar] [CrossRef]
- Jiang, M.; Wen, H.; Gou, G.W.; Liu, T.L.; Lu, X.; Deng, D.F. Preliminary study to evaluate the effects of dietary bile acids on growth performance and lipid metabolism of juvenile genetically improved farmed tilapia (Oreochromis niloticus) fed plant ingredient-based diets. Aquac. Nutr. 2018, 24, 1175–1183. [Google Scholar] [CrossRef]
- Peng, X.R.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.Y.; Tang, L.; Zhou, X.Q. Supplementation exogenous bile acid improved growth and intestinal immune function associated with NF-kappaB and TOR signalling pathways in on-growing grass carp (Ctenopharyngodon idella): Enhancement the effect of protein-sparing by dietary lipid. Fish Shellfish Immunol. 2019, 92, 552–569. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Ma, A.; Yang, S.; Huang, Z. Integrated metabolome and transcriptome analyses revealing the effects of thermal stress on lipid metabolism in juvenile turbot Scophthalmus maximus. J. Therm. Biol. 2021, 99, 102937. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysi; Association of Official Analytical Chemists: Arlington, VA, USA, 1995. [Google Scholar]
- Liu, H.; Li, X.; Lei, H.; Li, D.; Chen, H.; Schlenk, D.; Yan, B.; Yongju, L.; Xie, L. Dietary seleno-L-methionine alters the microbial communities and causes damage in the gastrointestinal tract of Japanese medaka Oryzias latipes. Environ. Sci. Technol. 2021, 55, 16515–16525. [Google Scholar] [CrossRef]
- Im, K.; Mareninov, S.; Diaz, M.F.P.; Wang, W.H. An introduction to performing immunofluorescence staining. In Biobanking: Methods and Protocols; Springer: New York, NY, USA, 2019; pp. 299–311. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Cao, X.; Wang, J.; Gong, Y.; Wang, X.; Lai, W.; Bu, X.; Zheng, J.; Mai, K.; et al. Effects of supplemental mixed bile acids on growth performance, body composition, digestive enzyme activities, skin color, and flesh quality of juvenile large yellow croaker (Larimichthys crocea) in soybean oil based diet. Front. Mar. Sci. 2023, 10, 1149887. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, X.; Yin, Y.; Wang, K.; Sun, Y.; Ye, J. Dietary high β-conglycinin reduces the growth through enhancing hepatic lipid peroxidation and impairing intestinal barrier function of orange-spotted grouper (Clostridium autoethanogenum). Front. Mar. Sci. 2023, 10, 1237387. [Google Scholar] [CrossRef]
- Bhusare, S.; Sardar, P.; Sahu, N.P.; Shamna, N.; Kumar, P.; Paul, M.; Jana, P.; Raghuvaran, N.; Bhavatharaniya, U. Bile acid improves growth, lipid utilization and antioxidative status of genetically improved farmed tilapia (Oreochromis niloticus) fed with varying protein-lipid diets reared in inland saline water. Anim. Feed Sci. Technol. 2023, 303, 115677. [Google Scholar] [CrossRef]
- Xia, R.; Zhang, Q.; Xia, D.; Hao, Q.; Ding, Q.; Ran, C.; Yang, Y.; Cao, A.; Zhang, Z.; Zhou, Z. The direct and gut microbiota-mediated effects of dietary bile acids on the improvement of gut barriers in largemouth bass (Micropterus salmoides). Anim. Nutr. 2023, 14, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhao, P.; Zhai, S. Dietary bile acids supplementation mainly regulates the amino acid metabolic pathways without decreasing bile acids levels in the liver of farmed European eel (Anguilla anguilla) juveniles. Aquac. Rep. 2022, 26, 101283. [Google Scholar] [CrossRef]
- Gu, M.; Bai, N.; Kortner, T.M. Taurocholate supplementation attenuates the changes in growth performance, feed utilization, lipid digestion, liver abnormality and sterol metabolism in turbot (Scophthalmus maximus) fed high level of plant protein. Aquaculture 2017, 468, 597–604. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Abdel-Latif, H.M.R.; Basuini, M.F.E.; El-Nokrashy, A.M.; Khaled, A.A.; Kord, M.; Soliman, A.A.; Zaki, M.; Nour, A.; Labib, E.M.H.; et al. Effects of exogenous bile acids (BAs) on growth, lipid profile, digestive enzymes, and immune responses of thinlip mullet, Liza ramada. Sci. Rep. 2023, 13, 22875. [Google Scholar] [CrossRef] [PubMed]
- Romański, K.W. The role and mechanism of action of bile acids within the digestive system—Bile acids in the liver and bile. Adv. Clin. Exp. Med. 2007, 16, 793–799. [Google Scholar]
- Guo, J.L.; Kuang, W.M.; Zhong, Y.F.; Zhou, Y.L.; Chen, Y.J.; Lin, S.M. Effects of supplemental dietary bile acids on growth, liver function and immunity of juvenile largemouth bass (Micropterus salmoides) fed high-starch diet. Fish Shellfish Immunol. 2020, 97, 602–607. [Google Scholar] [CrossRef]
- Adhami, B.; Amirkolaie, A.K.; Oraji, H.; Kenari, R.E. Growth performance, nutrient digestibility and lipase activity in juvenile rainbow trout (Oncorhynchus mykiss) fed fat powder in diet containing emulsifiers (cholic acid and Tween-80). Aquac. Nutr. 2017, 23, 1153–1159. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, X.; Cao, Y.; An, X.; Chen, J.; Yang, L. Punicalagin protects against diabetic liver injury by upregulating mitophagy and antioxidant enzyme activities. Nutrients 2022, 14, 2782. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, K.; Xu, Y.; Xu, Y.; Li, Y.; Xu, J.; Zhu, Y.; Adorini, L.; Lee, Y.K.; Kasumov, T.; Yin, L.; et al. Reversal of metabolic disorders by pharmacological activation of bile acid receptors TGR5 and FXR. Mol. Metab. 2018, 9, 131–140. [Google Scholar] [CrossRef]
- Makishima, M.; Lu, T.T.; Xie, W.; Whitfield, G.K.; Domoto, H.; Evans, R.M.; Haussler, M.R.; Mangelsdorf, D.J. Vitamin D receptor as an intestinal bile acid sensor. Science 2002, 296, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Fan, W.; Tang, T.; Wan, H.; Zhao, S.; Tan, Y.; Oware, K.A.; Tan, J.; Li, J.; Qu, S.; et al. Farnesoid X receptor deficiency induces hepatic lipid and glucose metabolism disorder via regulation of pyruvate dehydrogenase kinase 4. Oxid. Med. Cell. Longev. 2022, 2022, 3589519–3589525. [Google Scholar] [CrossRef]
- Yamagata, K.; Daitoku, H.; Shimamoto, Y.; Matsuzaki, H.; Hirota, K.; Ishida, J.; Fukamizu, A. Bile acids regulate gluconeogenic gene expression via small heterodimer partner-mediated repression of hepatocyte nuclear factor 4 and foxo1. J. Biol. Chem. 2004, 279, 23158–23165. [Google Scholar] [CrossRef] [PubMed]
- Sheikhzadeh, N.; Ahmadifar, E.; Soltani, M.; Tayefi-Nasrabadi, H.; Mousavi, S.; Naiel, M.A.E. Brown seaweed (Padina australis) extract can promote performance, innate immune responses, digestive enzyme activities, intestinal gene expression and resistance against aeromonas hydrophila in common carp (Cyprinus carpio). Animals 2022, 12, 3389. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Chen, Y.P.; Yang, M.X.; Zhang, L.L.; Lu, Z.X.; Zhou, Y.M.; Wang, T. Bacillus amyloliquefaciens supplementation alleviates immunological stress and intestinal damage in lipopolysaccharide-challenged broilers. Anim. Feed Sci. Tech. 2015, 208, 119–131. [Google Scholar] [CrossRef]
- Ze, Y.; Sheng, L.; Zhao, X.; Hong, J.; Ze, X.; Yu, X.; Pan, X.; Lin, A.; Zhao, Y.; Zhang, C.; et al. TiO2 nanoparticles induced hippocampal neuroinflammation in mice. PLoS ONE 2014, 9, e92230. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y. Intestinal villus structure contributes to even shedding of epithelial cells. Biophys. J. 2021, 120, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wan, M.; Wang, Z.; Zhang, H.; Zhu, S.; Cao, X.; Xu, N.; Zheng, J.; Bu, X.; Xu, W.; et al. Effects of tributyrin supplementation on growth performance, intestinal digestive enzyme activity, antioxidant capacity, and inflammation-related gene expression of large yellow croaker (Larimichthys crocea) Fed with a High Level of Clostridium autoethanogenum Protein. Aquac. Nutr. 2023, 2023, 2687712–2687734. [Google Scholar]
- Torun, A.N.; Kulaksizoglu, S.; Kulaksizoglu, M.; Pamuk, B.O.; Isbilen, E.; Tutuncu, N.B. Serum total antioxidant status and lipid peroxidation marker malondialdehyde levels in overt and subclinical hypothyroidism. Clin. Endocrinol. 2009, 70, 469–474. [Google Scholar] [CrossRef]
- Turk, R.; Juretić, D.; Gereš, D.; Svetina, A.; Turk, N.; Flegar-Meštrić, Z. Influence of oxidative stress and metabolic adaptation on PON1 activity and MDA level in transition dairy cows. Anim. Reprod. Sci. 2008, 108, 98–106. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Hu, Y.; Cheng, J.; Cheng, X.; Cheng, P.; Cui, Z. Dietary bile acid supplementation reveals beneficial effects on intestinal healthy status of tongue sole (Cynoglossus semiliaevis). Fish Shellfish Immunol. 2021, 116, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; An, Z.; Shi, C.; Li, P.; Liu, L. A sensitive and efficient method for simultaneous profiling of bile acids and fatty acids by UPLC-MS/MS. J. Pharm. Biomed. Anal. 2020, 178, 112815. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Shu, T.; Liu, G.; Mei, H.; Zhu, X.; Huang, X.; Zhang, L.; Jiang, Z. Quantitative profiling of 19 bile acids in rat plasma, liver, bile and different intestinal section contents to investigate bile acid homeostasis and the application of temporal variation of endogenous bile acids. J. Steroid Biochem. Mol. Biol. 2017, 172, 69–78. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Bile Acids Supplementation (mg/kg Diet) | |||
|---|---|---|---|---|
| Con | BA400 | BA800 | BA1200 | |
| Fishmeal 1 | 300.00 | 300.00 | 300.00 | 300.00 |
| Poultry by-product meal | 80.00 | 80.00 | 80.00 | 80.00 |
| Wheat gluten | 30.00 | 30.00 | 30.00 | 30.00 |
| Soybean meal | 285.00 | 285.00 | 285.00 | 285.00 |
| Wheat flour | 200.00 | 200.00 | 200.00 | 200.00 |
| Squid paste | 10.00 | 10.00 | 10.00 | 10.00 |
| Fish oil | 20.00 | 20.00 | 20.00 | 20.00 |
| Soybean oil | 20.00 | 20.00 | 20.00 | 20.00 |
| Lecithin | 20.00 | 20.00 | 20.00 | 20.00 |
| Ca(H2PO4)2 | 18.00 | 18.00 | 18.00 | 18.00 |
| L-Ascorbate-2-phosphate | 0.50 | 0.50 | 0.50 | 0.50 |
| Vitamin Premix 2 | 2.00 | 2.00 | 2.00 | 2.00 |
| Mineral Premix 3 | 5.00 | 5.00 | 5.00 | 5.00 |
| Choline chloride | 5.00 | 5.00 | 5.00 | 5.00 |
| Microcrystalline cellulose | 4.50 | 4.10 | 3.70 | 3.30 |
| Bile acids 4 | 0.00 | 0.40 | 0.80 | 1.20 |
| Proximate composition | ||||
| Crude protein | 426.00 | 430.0 | 431.00 | 432.00 |
| Crude lipid | 121.00 | 118.00 | 119.70 | 117.00 |
| Crude ash | 80.40 | 79.80 | 80.40 | 80.30 |
| Target Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | AT/°C | AE |
|---|---|---|---|---|
| pepck | CGGGAGAACATCACACACCT | CAGTGGGTCGATGATGGGAC | 60 | 1.07 |
| g6pase | CAGGTCATGGGGTACTGCTC | TTCCCGCTTTGGTTTCACCT | 60 | 1.06 |
| fbp | AACTGAGAAAGTCCCCCGAC | CCGGCCAAAACCTCGTATCT | 60 | 0.92 |
| hk | CTGGCTTGTGGGGACAGATT | GAGGCTGGCCCTCTTTATCC | 60 | 1.05 |
| pk | GTGGCCCAATCCAAATGTCC | GCAAGAGTGAGAGTTGGGGT | 60 | 1.08 |
| pfk | CGAGGGGCTAAATGTCAGGG | AAGGGGCATTCCGGTGATTT | 60 | 0.91 |
| gys | CGCATCCAGGAGTTCATCAGAGG | GTAGCGTCCAGCAATGAAGAAGAAG | 60 | 1.09 |
| pygl | TGTGATGGTTCTGTCGCTGGAG | AAGGAGTGGACGAAGATGGTGATC | 60 | 0.98 |
| fxr | GGAGGACAGGATACGCAAGAGTG | CAGGATGGTTACGGTGGTGAGG | 60 | 0.96 |
| tgr5 | AGCGGTATGGTGATGGCGTAG | CATGACGGACAGCAACGACTC | 60 | 1.03 |
| tnf-α | GATCGTCATCCCACAAACCG | GCTTTGCTGCCTATGGAGTC | 60 | 1.03 |
| il-1β | GTCAACTTACGTGCACCCTG | AAATCGTACCATGTCGCTGC | 60 | 0.95 |
| il-8 | GGATCAGTTTCTTCACCCAGG | CAGGTGGAGTCGAGGATCAT | 60 | 1.00 |
| tgf-β | ACCGACAATGAGCAGGGTTT | GGTGGCTGCTGATGTTTTGG | 60 | 0.96 |
| il-4 | ACCATGCATTACTACAGCACTG | CACATTCAGGGGCGTTTGTC | 60 | 1.06 |
| il-10 | TTCAAAACTCCGTTCGCCTG | TCACTCTTGAGCTCGTCGAA | 60 | 0.97 |
| nfκb | TGTGGTGTACGTACCGCTTC | TTCTCACACGGCTGGACTAC | 60 | 1.03 |
| ikbα | GCACGAGTGGAAGACGCAGATC | CGTCCGCCTGGTTCGTTATTACA | 60 | 0.93 |
| ikkβ | TCGGCAGCAGCTCCATCACA | AGGTGGTGCGTCTGGTGGTT | 60 | 1.03 |
| ikkα | ACAGCCAGCACCTCTTCATCCA | ACCAGCATCCAGCACGACCTT | 60 | 1.05 |
| β-actin | CAACTGGGATGACATGGAGAAG | TTGGCTTTGGGGTTCAGG | 60 | 1.08 |
| 18S | GGGTCCGAAGCGTTTACT | TCACCTCTAGCGGCACAA | 60 | 0.94 |
| Items | Con | BA400 | BA800 | BA1200 |
|---|---|---|---|---|
| FBW (g) | 130.5 ± 3.46 a | 135.53 ± 5.94 ab | 147.6 ± 2.19 b | 136.43 ± 6.54 ab |
| WG (%) | 539.38 ± 17.14 a | 562.83 ± 29.47 ab | 625.18 ± 10.54 b | 566.54 ± 31.65 ab |
| FCR | 1.22 ± 0.02 | 1.31 ± 0.04 | 1.22 ± 0.05 | 1.28 ± 0.08 |
| FI (g/fish) | 133.34 ± 4.8 a | 148.84 ± 3.76 b | 154.55 ± 4.35 b | 147.24 ± 2.33 b |
| AFR (%) | 4.41 ± 0.1 | 4.70 ± 0.17 | 4.89 ± 0.03 | 4.99 ± 0.40 |
| Survival (%) | 98.33 ± 1.67 | 95.00 ± 0.00 | 100.00 ± 0.00 | 98.33 ± 1.67 |
| Items | Con | BA400 | BA800 | BA1200 |
|---|---|---|---|---|
| Moisture | 69.98 ± 0.35 | 69.46 ± 0.41 | 69.15 ± 0.46 | 70.03 ± 0.24 |
| Crude protein | 16.7 ± 0.19 | 16.55 ± 0.29 | 16.23 ± 0.39 | 16.5 ± 0.08 |
| Crude lipid | 7.08 ± 0.38 a | 7.4 ± 0.2 ab | 8.15 ± 0.52 b | 7.3 ± 0.04 ab |
| Crude ash | 5.01 ± 0.11 b | 4.71 ± 0.15 ab | 4.57 ± 0.04 a | 4.75 ± 0.12 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, X.; Lin, J.; Song, K.; Li, X.; Wang, L.; Zhang, C.; Lu, K. Effects of Dietary Supplementation of Bile Acids on Growth, Glucose Metabolism, and Intestinal Health of Spotted Seabass (Lateolabrax maculatus). Animals 2024, 14, 1299. https://doi.org/10.3390/ani14091299
Liu Y, Li X, Lin J, Song K, Li X, Wang L, Zhang C, Lu K. Effects of Dietary Supplementation of Bile Acids on Growth, Glucose Metabolism, and Intestinal Health of Spotted Seabass (Lateolabrax maculatus). Animals. 2024; 14(9):1299. https://doi.org/10.3390/ani14091299
Chicago/Turabian StyleLiu, Yongping, Xiao Li, Jibin Lin, Kai Song, Xueshan Li, Ling Wang, Chunxiao Zhang, and Kangle Lu. 2024. "Effects of Dietary Supplementation of Bile Acids on Growth, Glucose Metabolism, and Intestinal Health of Spotted Seabass (Lateolabrax maculatus)" Animals 14, no. 9: 1299. https://doi.org/10.3390/ani14091299