
Alginate Oligosaccharide Alleviates Lipopolysaccharide-Induced Apoptosis and Inflammatory Response of Rumen Epithelial Cells through NF-κB Signaling Pathway


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Source of AOS and LPS, Configuration of AOS
2.3. Experimental Design
2.3.1. Effect of AOS on the Activity and Cytokines of ORECs
2.3.2. Construction of In Vitro Injury Model
2.3.3. Effect of AOS on Apoptosis and Inflammatory Response
2.4. Cell Viability
2.5. Immunoglobulin and Cytokine
2.6. ROS Detection
2.7. Apoptosis Rate
2.8. RNA Extraction, Reverse Transcription
2.9. RT-qPCR
2.10. Western Blot
2.11. Statistical Analysis
3. Results
3.1. Effect of AOS on the Viability of ORECs
3.2. Effect of AOS on the Content of ROS, Cytokine and Immunoglobulin in ORECs
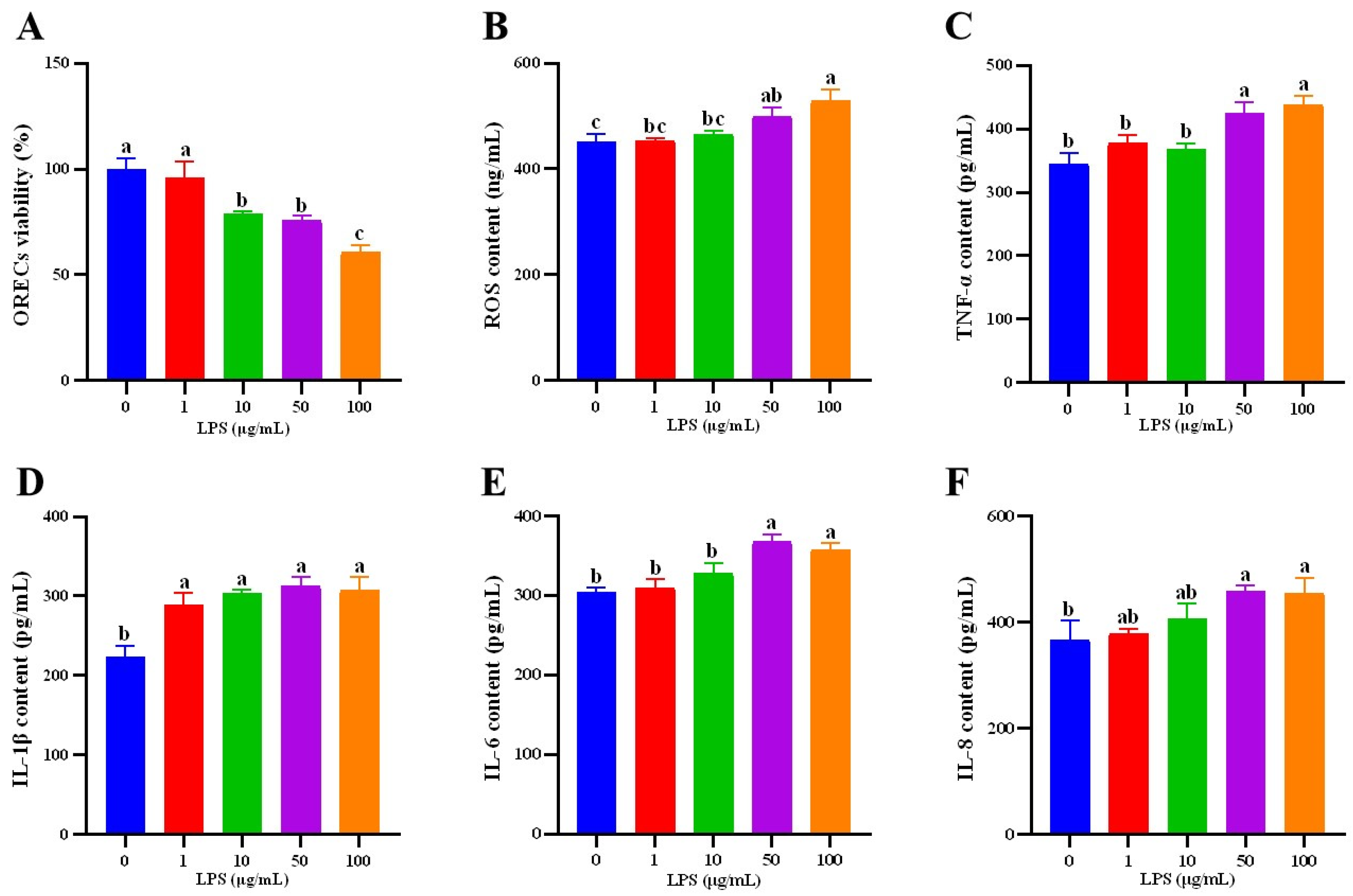
3.3. Establishment of ORECs Inflammatory Injury Model
3.4. Effects of AOS on LPS-Induced ORECs Viability, Immunoglobulins, Cytokines and Tight Junction Proteins
3.5. Effect of AOS on LPS-Induced ORECs ROS Content
3.6. Effects of AOS on LPS-Induced ORECs Apoptosis Rate and Apoptosis-Related Genes
3.7. Effects of AOS on LPS-Induced ORECs NF-κB Signaling Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andersen, P.H.; Bergelin, B.; Christensen, K.A. Effect of Feeding Regimen on Concentration of Free Endotoxin in Ruminal Fluid of Cattle. J. Anim. Sci. 1994, 72, 487–491. [Google Scholar] [CrossRef]
- Zhu, B.; Ni, F.; Sun, Y.; Ning, L.; Yao, Z. Elucidation of degrading pattern and substrate recognition of a novel bifunctional alginate lyase from Flammeovirga sp. NJ-04 and its use for preparation alginate oligosaccharides. Biotechnol. Biofuels 2019, 12, 13. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, Z.; Yang, G.; Wan, J.; Li, G.; Du, L.; Lu, P. Alginate oligosaccharide indirectly affects toll-like receptor signaling via the inhibition of microRNA-29b in aneurysm patients after endovascular aortic repair. Drug Des. Dev. Ther. 2017, 11, 2565–2579. [Google Scholar] [CrossRef]
- Wu, A.; Gao, Y.; Kan, R.; Ren, P.; Xue, C.; Kong, B.; Tang, Q. Alginate Oligosaccharides Prevent Dextran-Sulfate-Sodium-Induced Ulcerative Colitis via Enhancing Intestinal Barrier Function and Modulating Gut Microbiota. Foods 2023, 12, 220. [Google Scholar] [CrossRef]
- Guo, J.-J.; Ma, L.-L.; Shi, H.-T.; Zhu, J.-B.; Wu, J.; Ding, Z.-W.; An, Y.; Zou, Y.-Z.; Ge, J.-B. Alginate Oligosaccharide Prevents Acute Doxorubicin Cardiotoxicity by Suppressing Oxidative Stress and Endoplasmic Reticulum-Mediated Apoptosis. Mar. Drugs 2016, 14, 231. [Google Scholar] [CrossRef]
- Kawada, A.; Hiura, N.; Shiraiwa, M.; Tajima, S.; Hiruma, M.; Hara, K.; Ishibashi, A.; Takahara, H. Stimulation of human keratinocyte growth by alginate oligosaccharides, a possible co-factor for epidermal growth factor in cell culture. FEBS Lett. 1997, 408, 43–46. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, J.; Xu, Q.; Yin, H.; Chen, D.; Yu, B.; He, J. Alginate oligosaccharide protects against enterotoxigenic Escherichia coli-induced porcine intestinal barrier injury. Carbohydr. Polym. 2021, 270, 118316. [Google Scholar] [CrossRef]
- Zhao, J.; Han, Y.; Wang, Z.; Zhang, R.; Wang, G.; Mao, Y. Alginate oligosaccharide protects endothelial cells against oxidative stress injury via integrin-α/FAK/PI3K signaling. Biotechnol. Lett. 2020, 42, 2749–2758. [Google Scholar] [CrossRef]
- Reisinger, N.; Wendner, D.; Schauerhuber, N.; Mayer, E. Effect of Lipopolysaccharides (LPS) and Lipoteichoic Acid (LTA) on the Inflammatory Response in Rumen Epithelial Cells (REC) and the Impact of LPS on Claw Explants. Animals 2021, 11, 2058. [Google Scholar] [CrossRef]
- Yan, G.L.; Guo, Y.M.; Yuan, J.M.; Liu, D.; Zhang, B.K. Sodium alginate oligosaccharides from brown algae inhibit Salmonella Enteritidis colonization in broiler chickens. Poult. Sci. 2011, 90, 1441–1448. [Google Scholar] [CrossRef]
- Bai, N.; Deng, W.; Qi, Z.; Pan, S.; Li, Q.; Gu, M. The effect of alginate oligosaccharides on intestine barrier function and Vibrio parahaemolyticus infections in the white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2023, 141, 109011. [Google Scholar] [CrossRef]
- Zhang, L.F. Effects of Different Microecological Regulators Ongrowth Performance and Rumen Microecology of Beefcattle. Master’s Thesis, Guangxi University, Nanning, China, 2022; pp. 1–77. [Google Scholar]
- Li, R.; Fang, H.; Shen, J.; Jin, Y.; Zhao, Y.; Wang, R.; Fu, Y.; Tian, Y.; Yu, H.; Zhang, J. Curcumin Alleviates LPS-Induced Oxidative Stress, Inflammation and Apoptosis in Bovine Mammary Epithelial Cells via the NFE2L2 Signaling Pathway. Toxins 2021, 13, 208. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Yang, S.; Lian, G. ROS and diseases: Role in metabolism and energy supply. Mol. Cell Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial Reactive Oxygen Species (ROS) and ROS-Induced ROS Release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Huang, W.; Wang, G.; Lu, S.-E.; Kipen, H.; Wang, Y.; Hu, M.; Lin, W.; Rich, D.; Ohman-Strickland, P.; Diehl, S.R.; et al. Inflammatory and Oxidative Stress Responses of Healthy Young Adults to Changes in Air Quality during the Beijing Olympics. Am. J. Respir. Crit. Care Med. 2012, 186, 1150–1159. [Google Scholar] [CrossRef]
- Rastegari-Pouyani, M.; Mostafaie, A.; Mansouri, K.; Mortazavi-Jahromi, S.S.; Mohammadi-Motlagh, H.R.; Mirshafiey, A. Anti-angiogenesis effect of β-D-mannuronic acid (M2000) as a novel NSAID with immunosuppressive properties under experimental model. Clin. Exp. Pharmacol. Physiol. 2018, 45, 370–376. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, Y.; Yuan, X.; Sun, G.; Shen, B.; Xu, F.; Fan, G.; Jin, M.; Li, X.; Liu, G. Berberine inhibits lipopolysaccharide-induced expression of inflammatory cytokines by suppressing TLR4-mediated NF-ĸB and MAPK signaling pathways in rumen epithelial cells of Holstein calves. J. Dairy Res. 2019, 86, 171–176. [Google Scholar] [CrossRef]
- Kent-Dennis, C.; Aschenbach, J.R.; Griebel, P.J.; Penner, G.B. Effects of lipopolysaccharide exposure in primary bovine ruminal epithelial cells. J. Dairy Sci. 2020, 103, 9587–9603. [Google Scholar] [CrossRef]
- Lu, S.; Na, K.; Wei, J.; Tao, T.; Zhang, L.; Fang, Y.; Li, X.; Guo, X. Alginate oligosaccharide structures differentially affect DSS-induced colitis in mice by modulating gut microbiota. Carbohydr. Polym. 2023, 312, 120806. [Google Scholar] [CrossRef]
- Kawada, A.; Hiura, N.; Tajima, S.; Takahara, H. Alginate oligosaccharides stimulate VEGF-mediated growth and migration of human endothelial cells. Arch. Dermatol. Res. 1999, 291, 542–547. [Google Scholar] [CrossRef]
- Steele, M.A.; Penner, G.B.; Chaucheyras-Durand, F.; Guan, L.L. Development and physiology of the rumen and the lower gut: Targets for improving gut health. J. Dairy Sci. 2016, 99, 4955–4966. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Zebeli, Q.; Patra, A.K.; Greco, G.; Amasheh, S.; Penner, G.B. Symposium review: The importance of the ruminal epithelial barrier for a healthy and productive cow. J. Dairy Sci. 2019, 102, 1866–1882. [Google Scholar] [CrossRef]
- Baaske, L.; Gäbel, G.; Dengler, F. Ruminal epithelium: A checkpoint for cattle health. J. Dairy Res. 2020, 87, 322–329. [Google Scholar] [CrossRef]
- Stumpff, F.; Georgi, M.-I.; Mundhenk, L.; Rabbani, I.; Fromm, M.; Martens, H.; Günzel, D. Sheep rumen and omasum primary cultures and source epithelia: Barrier function aligns with expression of tight junction proteins. J. Exp. Biol. 2011, 214, 2871–2882. [Google Scholar] [CrossRef]
- Liu, J.-H.; Xu, T.-T.; Liu, Y.-J.; Zhu, W.-Y.; Mao, S.-Y. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, 232–241. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Khatib, K.; Guo, S.; Ye, D.; Youssef, M.; Ma, T. Occludin regulates macromolecule flux across the intestinal epithelial tight junction barrier. Am. J. Physiol.-Gastrointest. Liver Physiol. 2011, 300, G1054–G1064. [Google Scholar] [CrossRef]
- Chin, A.C.; Flynn, A.N.; Fedwick, J.P.; Buret, A.G. The role of caspase-3 in lipopolysaccharide-mediated disruption of intestinal epithelial tight junctions. Can. J. Physiol. Pharmacol. 2006, 84, 1043–1050. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Biosci. 2009, 14, 2765–2778. [Google Scholar] [CrossRef]
- Kaminsky, L.W.; Al-Sadi, R.; Ma, T.Y. IL-1β and the Intestinal Epithelial Tight Junction Barrier. Front. Immunol. 2021, 12, 767456. [Google Scholar] [CrossRef]
- Lin, J.; Wu, J.; Wang, F.; Tang, F.; Sun, J.; Xu, B.; Jiang, M.; Chu, Y.; Chen, D.; Li, X.; et al. QingBai decoction regulates intestinal permeability of dextran sulphate sodium-induced colitis through the modulation of notch and NF-κBsignalling. Cell Prolif. 2019, 52, e12547. [Google Scholar] [CrossRef] [PubMed]
- Varfolomeev, E.; Vucic, D. Intracellular regulation of TNF activity in health and disease. Cytokine 2018, 101, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Möller, B.; Villiger, P.M. Inhibition of IL-1, IL-6, and TNF-α in immune-mediated inflammatory diseases. Springer Semin. Immunopathol. 2006, 27, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, J.; Goodrich, M.E.; Eichelberger, H.; McGee, D.W. Polarized Secretion of IL-6 by IEC-6 Intestinal Epithelial Cells: Differential Effects of IL-1β and TNF-α. Immunol. Investig. 1996, 25, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, S.; Choi, H.B.; Kim, S.U.; McLarnon, J.G. IL-8 enhancement of amyloid-beta (Aβ1-42)-induced expression and production of pro-inflammatory cytokines and COX-2 in cultured human microglia. J. Neuroimmunol. 2005, 159, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, C.; Li, Y.; Han, X.; Luo, X.; Chen, L.; Zhang, T.; Wang, N.; Wang, W. Alginate Oligosaccharides Ameliorate DSS-Induced Colitis through Modulation of AMPK/NF-κB Pathway and Intestinal Microbiota. Nutrients 2022, 14, 2864. [Google Scholar] [CrossRef]
- Kornfeld, O.S.; Hwang, S.; Disatnik, M.-H.; Chen, C.-H.; Qvit, N.; Mochly-Rosen, D. Mitochondrial Reactive Oxygen Species at the Heart of the Matter. Circ. Res. 2015, 116, 1783–1799. [Google Scholar] [CrossRef]
- Viatour, P.; Merville, M.-P.; Bours, V.; Chariot, A. Phosphorylation of NF-κB and IκB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2020, 78, 1233–1261. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF-B Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.S., Jr. The NF-κB and IκB Proteins: New Discoveries and Insights. Annu. Rev. Immunol. 1996, 14, 649–681. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence, 5′to 3′ | Accession Number | Size (bp) |
|---|---|---|---|
| β-actin | F:ACAGCCGAGCGGGAAAT | XM_004013078.5 | 215 |
| R:ATGCCACAGGACTCCATACC | |||
| TLR4 | F:AAAGAACTTGGAGGAGGGC | XM_042242671.1 | 141 |
| R:TGCTGGGACACCACGAC | |||
| MyD88 | F:CCCATCAAGTACAAGCCAATG | NM_001166183.1 | 109 |
| R:CGAGGCGAGTCCAGAACC | |||
| NF-kB | F:GTTTACGCCTGATGATTTGC | XM_042251202.1 | 180 |
| R:TTGGAGGGAGCGGGACT | |||
| ZO-1 | F:TGAACGCAAGTTTGAAAGTCC | XM_042235171.1 | 288 |
| R:CGGGCAGTAGCCACCAC | |||
| Occludin | F:CGGAGGAAGTGCCTTTGG | XM_012096797.4 | 101 |
| R:CCTTTGCCGCTCTTGGAT | |||
| Claudin-1 | F:GGGGCTGTGGATGTCGT | NM_001185016.1 | 202 |
| R:CTTCTGTGCCTCGTCGTCT | |||
| Claudin-4 | F:TTCATCGGCAGCAACATCG | NM_001185017.2 | 189 |
| R: CAACAGCACGCCAAACACG | |||
| BCL-2 | F:CGCATCGTGGCCTTCTT | XM_027960877.2 | 291 |
| R:TCCCAGCCTCCGTTGTC | |||
| BAX | F:TCCGACGGCAACTTCAA | XM_027978594.2 | 242 |
| R:AGCACTCCAGCCACAAAGA | |||
| BAD | F:TCCCAGAGTTTGAGCAGAGTGA | XM_042238345.2 | 428 |
| R:GCTAGGGCTTTGTCGCATTT | |||
| Caspase3 | F:TGGAACCAATGGACCCG | XM_027962551.2 | 247 |
| R:CTTTGAGTTTCGCCAGGAA | |||
| Caspase9 | F:ACCAGCAGACAAGCAGCAA | XM_042257438.1 | 253 |
| R:CAGTGAATCCTCCAGAACCAA | |||
| Caspase8 | F:AATGCCCTTCCCTTGTCG | XM_042244960.1 | 173 |
| R:CAGCAGAAAGTCAGCCTCAT |
| AOS (µg/mL) | Time (h) | p | |||
|---|---|---|---|---|---|
| 6 | 12 | 24 | 48 | ||
| 0 | 107.66 ± 10.04 | 101.39 ± 5.02 c | 106.31 ± 8.52 ab | 100.00 ± 4.52 a | 0.423 |
| 200 | 121.29 ± 12.99 AB | 131.00 ± 14.48 Ab | 111.60 ± 18.02 ABab | 100.56 ± 8.24 Ba | 0.049 |
| 400 | 101.20 ± 14.20 B | 137.59 ± 9.18 Ab | 126.12 ± 23.27 Aa | 99.23 ± 7.85 Ba | 0.008 |
| 600 | 108.37 ± 10.67 B | 160.80 ± 13.52 Aa | 106.97 ± 6.94 Bab | 87.97 ± 6.76 Cb | <0.001 |
| 800 | 134.21 ± 45.63 A | 157.33 ± 11.16 Aa | 95.01 ± 3.24 Bb | 80.51 ± 5.64 Bb | 0.002 |
| p | 0.313 | <0.001 | 0.083 | 0.002 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, X.; Yin, F.; Du, C.; Ma, J.; Gan, S. Alginate Oligosaccharide Alleviates Lipopolysaccharide-Induced Apoptosis and Inflammatory Response of Rumen Epithelial Cells through NF-κB Signaling Pathway. Animals 2024, 14, 1298. https://doi.org/10.3390/ani14091298
Qiu X, Yin F, Du C, Ma J, Gan S. Alginate Oligosaccharide Alleviates Lipopolysaccharide-Induced Apoptosis and Inflammatory Response of Rumen Epithelial Cells through NF-κB Signaling Pathway. Animals. 2024; 14(9):1298. https://doi.org/10.3390/ani14091298
Chicago/Turabian StyleQiu, Xiaoyuan, Fuquan Yin, Chunmei Du, Jian Ma, and Shangquan Gan. 2024. "Alginate Oligosaccharide Alleviates Lipopolysaccharide-Induced Apoptosis and Inflammatory Response of Rumen Epithelial Cells through NF-κB Signaling Pathway" Animals 14, no. 9: 1298. https://doi.org/10.3390/ani14091298