

Effects of Dietary Terminalia chebula Extract on Growth Performance, Immune Function, Antioxidant Capacity, and Intestinal Health of Broilers
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Management
2.2. Sample Collection
2.3. Growth Performance
2.4. Immune Function
2.5. Antioxidant Capacity
2.6. Intestinal Morphology
2.7. Short-Chain Fatty Acid Analysis
2.8. 16S rRNA Sequencing of Microflora in Cecum Contents
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Immune Function
3.3. Antioxidant Capacity
3.4. Intestinal Morphology
3.5. SCFA Analysis
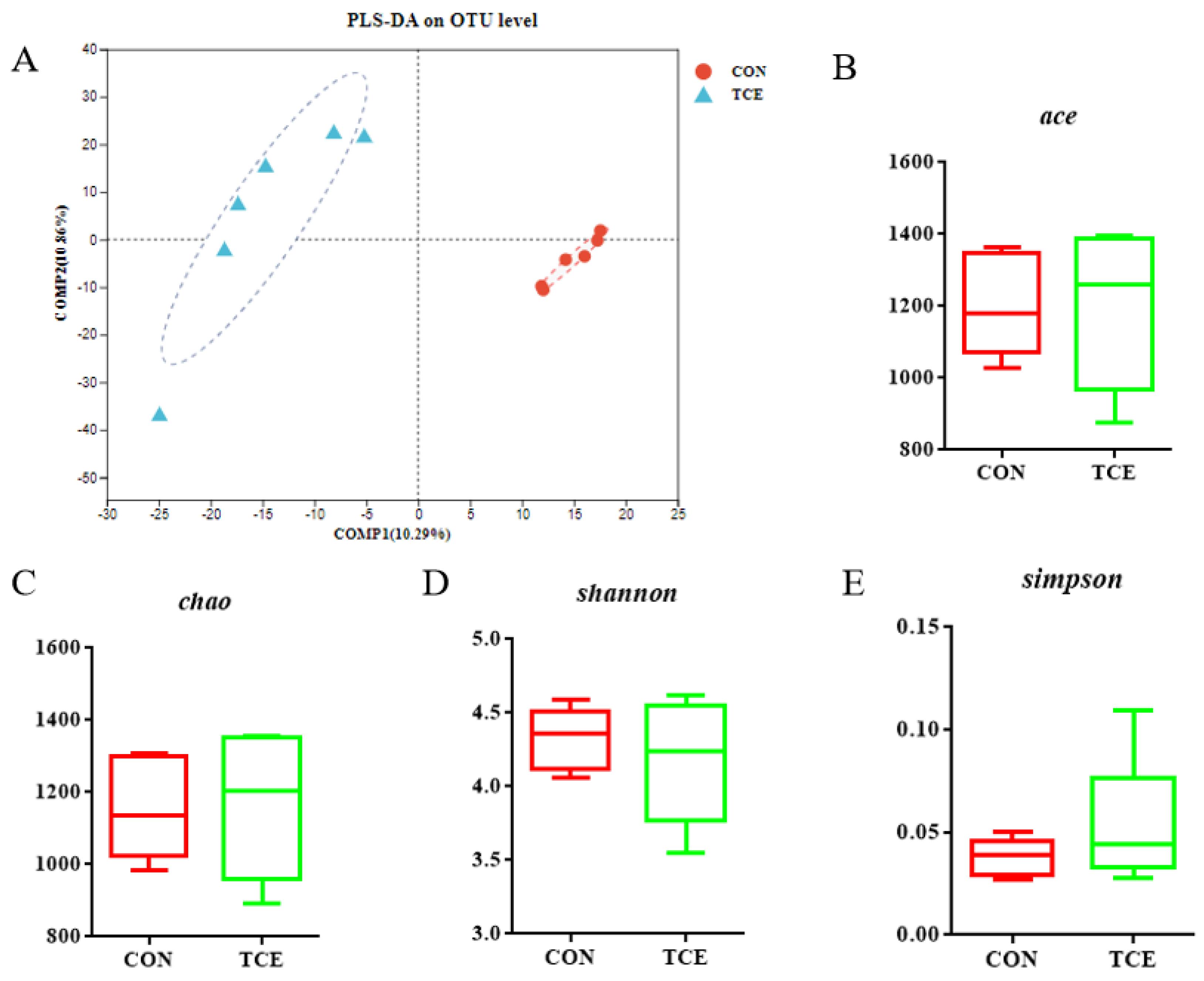
3.6. 16S rRNA Sequencing of Microflora in Cecum Contents
3.7. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, H.; Li, T.; Zhang, Y.; Liu, X.; Gong, R.; Bao, J.; Li, J. Cold stimulation causes oxidative stress, inflammatory response and apoptosis in broiler heart via regulating Nrf2/HO-1 and NF-κB pathway. J. Therm. Biol. 2023, 116, 103658. [Google Scholar] [CrossRef]
- Hu, P.; Li, K.; Peng, X.; Yao, T.; Zhu, C.; Gu, H.; Liu, H.Y.; Sun, M.A.; Hu, Y.; Ennab, W.; et al. Zinc intake ameliorates intestinal morphology and oxidative stress of broiler chickens under heat stress. Front. Immunol. 2024, 14, 1308907. [Google Scholar] [CrossRef]
- Nasr, M.A.F.; Alkhedaide, A.Q.; Ramadan, A.A.I.; Hafez, A.S.E.; Hussein, M.A. Potential impact of stocking density on growth, carcass traits, indicators of biochemical and oxidative stress and meat quality of different broiler breeds. Poult. Sci. 2021, 100, 101442. [Google Scholar] [CrossRef]
- Ballini, A.; Santacroce, L.; Cantore, S.; Bottalico, L.; Dipalma, G.; Topi, S.; Saini, R.; De Vito, D.; Inchingolo, F. Probiotics Efficacy on Oxidative Stress Values in Inflammatory Bowel Disease: A Randomized Double-Blinded Placebo-Controlled Pilot Study. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 373–381. [Google Scholar] [CrossRef]
- Lauridsen, C. From oxidative stress to inflammation: Redox balance and immune system. Poult. Sci. 2019, 98, 4240–4246. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, J.; Cao, Q.; Zhang, C.; Dong, Z.; Feng, D.; Ye, H.; Zuo, J. Dietary Catalase Supplementation Alleviates Deoxynivalenol-Induced Oxidative Stress and Gut Microbiota Dysbiosis in Broiler Chickens. Toxins 2022, 14, 830. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kwon, J.S.; Lee, H.J.; Song, C.S.; Lee, S.W. Eradication of Mycoplasma synoviae from a multi-age broiler breeder farm using antibiotics therapy. Poult. Sci. 2015, 94, 2364–2368. [Google Scholar] [CrossRef]
- Eisenreich, W.; Rudel, T.; Heesemann, J.; Goebel, W. Link Between Antibiotic Persistence and Antibiotic Resistance in Bacterial Pathogens. Front. Cell. Infect. Microbiol. 2022, 12, 900848. [Google Scholar] [CrossRef]
- Pratiwi, R.; Ramadhanti, S.P.; Amatulloh, A.; Megantara, S.; Subra, L. Recent Advances in the Determination of Veterinary Drug Residues in Food. Food 2023, 12, 3422. [Google Scholar] [CrossRef]
- Liu, R.; Wang, Y.; Wang, L.; Wang, Y.; Peng, X.; Cao, L.; Liu, Y. Spatio-temporal distribution and source identification of antibiotics in suspended matter in the Fen River Basin. Chemosphere 2023, 345, 140497. [Google Scholar] [CrossRef]
- Goodarzi Boroojeni, F.; Männer, K.; Zentek, J. The impacts of Macleaya cordata extract and naringin inclusion in post-weaning piglet diets on performance, nutrient digestibility and intestinal histomorphology. Arch. Anim. Nutr. 2018, 72, 178–189. [Google Scholar] [CrossRef]
- Roque-Borda, C.A.; Pereira, L.P.; Guastalli, E.A.L.; Soares, N.M.; Mac-Lean, P.A.B.; Salgado, D.D.; Meneguin, A.B.; Chorilli, M.; Vicente, E.F. HPMCP-Coated Microcapsules Containing the Ctx(Ile21)-Ha Antimicrobial Peptide Reduce the Mortality Rate Caused by Resistant Salmonella Enteritidis in Laying Hens. Antibiotics 2021, 10, 616. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Kim, I.H. Effects of multistrain probiotics on growth performance, apparent ileal nutrient digestibility, blood characteristics, cecal microbial shedding, and excreta odor contents in broilers. Poult. Sci. 2014, 93, 364–370. [Google Scholar] [CrossRef]
- Naiel, M.A.E.; Ismael, N.E.M.; Shehata, S.A. Ameliorative effect of diets supplemented with rosemary (Rosmarinus officinalis) on aflatoxin B1 toxicity in terms of the performance, liver histopathology, immunity and antioxidant activity of Nile Tilapia (Oreochromis niloticus). Aquaculture 2019, 511, 734264. [Google Scholar] [CrossRef]
- Qiao, Y.; Guo, Y.; Zhang, W.; Guo, W.; Oleksandr, K.; Bozhko, N.; Wang, Z.; Liu, C. Effects of Compound Polysaccharides Derived from Astragalus and Glycyrrhiza on Growth Performance, Meat Quality and Antioxidant Function of Broilers Based on Serum Metabolomics and Cecal Microbiota. Antioxidants 2022, 11, 1872. [Google Scholar] [CrossRef]
- Nalamolu, K.R.; Nammi, S. Antidiabetic and renoprotective effects of the chloroform extract of Terminalia chebula Retz. seeds in streptozotocin-induced diabetic rats. BMC Complement. Altern. Med. 2006, 6, 17. [Google Scholar]
- Datta, S.; Pal, N.K.; Nandy, A.K. In vitro Antibacterial Activity of Bioactive Potent Compounds from Terminalia chebula against Some Common Human Pathogens. Pharmacol. Pharm. 2017, 8, 283–391. [Google Scholar] [CrossRef]
- Majed, F.; Nafees, S.; Rashid, S.; Ali, N.; Hasan, S.K.; Ali, R.; Shahid, A.; Sultana, S. Terminalia chebula Attenuates DMBA/Croton Oil-Induced Oxidative Stress and Inflammation in Swiss albino Mouse Skin. Toxicol. Int. 2015, 22, 21–29. [Google Scholar]
- Bajpai, M.; Pande, A.; Tewari, S.K.; Prakash, D. Phenolic contents and antioxidant activity of some food and medicinal plants. Int. J. Food Sci. Nutr. 2005, 56, 287–491. [Google Scholar] [CrossRef]
- Patil, V.S.; Harish, D.R.; Vetrivel, U.; Roy, S.; Deshpande, S.H.; Hegde, H.V. Hepatitis C Virus NS3/4A Inhibition and Host Immunomodulation by Tannins from Terminalia chebula: A Structural Perspective. Molecules 2022, 27, 1076. [Google Scholar] [CrossRef]
- Liu, J.; Li, B.; Liu, J.; Qiu, F.; Diao, Y.; Lei, Y.; Liu, J.; Zhang, W. Phenolic Acids from Fructus Chebulae Immaturus Alleviate Intestinal Ischemia-Reperfusion Injury in Mice through the PPARα/NF-κB Pathway. Molecules 2022, 27, 5227. [Google Scholar] [CrossRef]
- Jafari, M.; Lorigooini, Z.; Kheiri, S.; Naeini, K.M. Anti-Toxoplasma Effect of Hydroalchohlic Extract of Terminalia chebula Retz in Cell Culture and Murine Model. Iran. J. Parasitol. 2021, 16, 631–640. [Google Scholar] [CrossRef]
- Sarabhai, S.; Sharma, P.; Capalash, N. Ellagic acid derivatives from Terminalia chebula Retz. downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS ONE 2013, 8, e53441. [Google Scholar] [CrossRef]
- He, M.; Wu, Q.; Wei, J.; Zhang, Y.; Guo, H.; Guo, X. Research Note: Effect of Rubia cordifolia L. processed Terminalia chebula Retz polysaccharide on the histological structure and apoptosis in the spleen in immunosuppressed Chinese yellow quail. Poult. Sci. 2023, 102, 102416. [Google Scholar] [CrossRef]
- Lee, H.H.; Paudel, K.R.; Kim, D.W. Terminalia chebula Fructus Inhibits Migration and Proliferation of Vascular Smooth Muscle Cells and Production of Inflammatory Mediators in RAW 264. 7. Evid. Based Complement. Alternat. Med. 2015, 2015, 502182. [Google Scholar]
- Jung, H.L.; Yang, S.Y.; Pyo, M.C.; Hong, C.O.; Nam, M.H.; Lee, J.W.; Lee, K.W. Protective effects of chebulic acid from Terminalia chebula Retz. against t-BHP-induced oxidative stress by modulations of Nrf2 and its related enzymes in HepG2 cells. Food Sci. Biotechnol. 2018, 28, 555–562. [Google Scholar] [CrossRef]
- Mandeville, A.; Cock, I.E. Terminalia chebula Retz. Fruit Extracts Inhibit Bacterial Triggers of Some Autoimmune Diseases and Potentiate the Activity of Tetracycline. Indian J. Microbiol. 2018, 58, 496–506. [Google Scholar] [CrossRef]
- Gaire, B.P.; Jamarkattel-Pandit, N.; Lee, D.; Song, J.; Kim, J.Y.; Park, J.; Jung, S.; Choi, H.Y.; Kim, H. Terminalia chebula extract protects OGD-R induced PC12 cell death and inhibits LPS induced microglia activation. Molecules 2013, 18, 3529–3542. [Google Scholar] [CrossRef]
- Dong, W.R.; Li, Y.Y.; Liu, T.T.; Zhou, G.; Chen, Y.X. Ethyl acetate extract of Terminalia chebula alleviates DSS-induced ulcerative colitis in C57BL/6 mice. Front. Pharmacol. 2023, 14, 1229772. [Google Scholar] [CrossRef]
- He, S.; Yu, Q.; He, Y.; Hu, R.; Xia, S.; He, J. Dietary resveratrol supplementation inhibits heat stress-induced high-activated innate immunity and inflammatory response in spleen of yellow-feather broilers. Poult. Sci. 2019, 98, 6378–6387. [Google Scholar] [CrossRef]
- Zhao, M.; Sun, Q.; Khogali, M.K.; Liu, L.; Geng, T.; Yu, L.; Gong, D. Dietary Selenized Glucose Increases Selenium Concentration and Antioxidant Capacity of the Liver, Oviduct, and Spleen in Laying Hens. Biol. Trace Elem. Res. 2021, 199, 4746–4752. [Google Scholar] [CrossRef]
- Tong, Y.; Yu, C.; Xie, Z.; Zhang, X.; Yang, Z.; Wang, T. Trans-anethole ameliorates lipopolysaccharide-induced acute liver inflammation in broilers via inhibiting NF-κB signaling pathway. Poult. Sci. 2022, 101, 101962. [Google Scholar] [CrossRef] [PubMed]
- Wickramasuriya, S.S.; Park, I.; Lee, K.; Lee, Y.; Kim, W.H.; Nam, H.; Lillehoj, H.S. Role of Physiology, Immunity, Microbiota, and Infectious Diseases in the Gut Health of Poultry. Vaccines 2022, 10, 172. [Google Scholar] [CrossRef] [PubMed]
- Reicher, N.; Uni, Z. Intestinal brush border assembly during the peri-hatch period and its contribution to surface area expansion. Poult. Sci. 2021, 100, 101401. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, D.Y.; Lee, J.; Kim, H.W.; Sung, S.H.; Han, J.S.; Jeon, W.K. Terminalia chebula extract prevents scopolamine-induced amnesia via cholinergic modulation and anti-oxidative effects in mice. BMC Complement. Altern. Med. 2018, 18, 136. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Song, H.K.; Park, S.H.; Jang, S.; Park, K.S.; Song, K.H.; Lee, S.K.; Kim, T. Terminalia chebula Retz. extract ameliorates the symptoms of atopic dermatitis by regulating anti-inflammatory factors in vivo and suppressing STAT1/3 and NF-ĸB signaling in vitro. Phytomedicine. 2022, 104, 154318. [Google Scholar] [CrossRef] [PubMed]
- NY/T 33-2004; Feeding Standard of Chicken. Ministry of Agriculture and Rural Affairs: Beijing, China, 2004.
- Xie, T.; Liu, W.; Chen, Y.; Zhou, Y. An evaluation of graded levels of beta-sitosterol supplementation on growth performance, antioxidant status, and intestinal permeability-related parameters and morphology in broiler chickens at an early age. Poult. Sci. 2022, 101, 102108. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Hu, Y.; Yao, X.; He, Y.; Chen, J.; Wu, J.; Wu, S.; Zhang, H.; He, X.; Song, Z. Dietary essential oils improves the growth performance, antioxidant properties and intestinal permeability by inhibiting bacterial proliferation, and altering the gut microbiota of yellow-feather broilers. Poult. Sci. 2022, 101, 102087. [Google Scholar] [CrossRef]
- Zhang, L.; Piao, X. Different dietary protein sources influence growth performance, antioxidant capacity, immunity, fecal microbiota and metabolites in weaned piglets. Anim. Nutr. 2022, 8, 71–81. [Google Scholar] [CrossRef]
- Song, Y.; Luo, Y.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Huang, Z.; Luo, J.; Luo, Y.; Yan, H.; et al. Tannic acid extracted from gallnut prevents post-weaning diarrhea and improves intestinal health of weaned piglets. Anim. Nutr. 2021, 7, 1078–1086. [Google Scholar] [CrossRef]
- Chang, W.Y.; Yu, Y.H. Effect of Bacillus species-fermented products and essential oils on growth performance, gut morphology, cecal short-chain fatty acid levels, and microbiota community in broilers. Poult. Sci. 2022, 101, 101970. [Google Scholar] [CrossRef]
- Wang, F.; Chen, J.; Yin, Y.; Yang, M.; Xiao, Y.; Cheng, Y.; Yin, L.; Fu, C. The effects of dietary ellagic acid supplementation on growth performance, immune response, antioxidant activity, digestive enzyme activities, and intestinal functions in yellow-feathered broilers. J. Anim. Sci. 2022, 100, skac301. [Google Scholar] [CrossRef]
- Hao, Y.; Ji, Z.; Shen, Z.; Wu, Y.; Zhang, B.; Tang, J.; Hou, S.; Xie, M. Effects of Total Dietary Fiber on Cecal Microbial Community and Intestinal Morphology of Growing White Pekin Duck. Front. Microbiol. 2021, 12, 727200. [Google Scholar] [CrossRef] [PubMed]
- Shendge, A.K.; Sarkar, R.; Mandal, N. Potent anti-inflammatory Terminalia chebula fruit showed in vitro anticancer activity on lung and breast carcinoma cells through the regulation of Bax/Bcl-2 and caspase-cascade pathways. J. Food Biochem. 2020, 44, e13521. [Google Scholar] [CrossRef]
- Kher, M.N.; Sheth, N.R.; Bhatt, V.D. In Vitro Antibacterial Evaluation of Terminalia chebula as an Alternative of Antibiotics against Bovine Subclinical Mastitis. Anim. Biotechnol. 2019, 30, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Haghani, M.; Jafari, M.; Meftahi, G.H.; Behzadnia, M.J.; Bahari, Z.; Salimi-Sabour, E.; Jangravi, Z. Analgesic effects of Terminalia chebula extract are mediated by the suppression of the protein expression of nerve growth factor and nuclear factor-κB in the brain and oxidative markers following neuropathic pain in rats. Mol. Biol. Rep. 2022, 49, 10457–10467. [Google Scholar] [CrossRef] [PubMed]
- Sobeh, M.; Mahmoud, M.F.; Hasan, R.A.; Abdelfattah, M.A.O.; Osman, S.; Rashid, H.O.; El-Shazly, A.M.; Wink, M. Chemical composition, antioxidant and hepatoprotective activities of methanol extracts from leaves of Terminalia bellirica and Terminalia sericea (Combretaceae). PeerJ 2019, 7, e6322. [Google Scholar] [CrossRef]
- Li, K.; Han, X.; Li, R.; Xu, Z.; Pan, T.; Liu, J.; Li, B.; Wang, S.; Diao, Y.; Liu, X. Composition, Antivirulence Activity, and Active Property Distribution of the Fruit of Terminalia chebula Retz. J. Food Sci. 2019, 84, 1721–1729. [Google Scholar] [CrossRef]
- Liu, F.; Zhan, S.; Zhan, P.; Jia, C.; Zhu, Q.; Dai, Q.; Yu, M.; Cheng, L.; Xiong, L.; Sun, F.; et al. Simultaneous quantitative analysis and in vitro anti-arthritic effects of five polyphenols from Terminalia chebula. Front. Physiol. 2023, 14, 1138947. [Google Scholar] [CrossRef]
- Nam, Y.J.; Hwang, Y.S. Antibacterial and antioxidant effect of ethanol extracts of Terminalia chebula on Streptococcus mutans. Clin. Exp. Dent. Res. 2021, 7, 987–994. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Xi, Y.; Wang, S.T.; Zheng, L.Y.; Qi, Y.; Guo, S.S.; Ding, B.Y. Effects of Chinese gallnut tannic acid on growth performance, blood parameters, antioxidative status, intestinal histomorphology, and cecal microbial shedding in broilers challenged with aflatoxin B1. J. Anim. Sci. 2022, 100, skac099. [Google Scholar] [CrossRef]
- Prihambodo, T.R.; Sholikin, M.M.; Qomariyah, N.; Jayanegara, A.; Batubara, I.; Utomo, D.B.; Nahrowi, N. Effects of dietary flavonoids on performance, blood constituents, carcass composition and small intestinal morphology of broilers: A meta-analysis. Anim. Biosci. 2021, 34, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.C.; Zhu, Y.R.; Zhao, Z.H.; Jiang, P.; Yin, F.Q. Effects of Dietary Supplementation of Algae-Derived Polysaccharides on Morphology, Tight Junctions, Antioxidant Capacity and Immune Response of Duodenum in Broilers under Heat Stress. Animals 2021, 11, 2279. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Liu, C.; Guo, Y.; Zhang, W.; Guo, W.; Oleksandr, K.; Wang, Z. Polysaccharides derived from Astragalus membranaceus and Glycyrrhiza uralensis improve growth performance of broilers by enhancing intestinal health and modulating gut microbiota. Poult. Sci. 2022, 101, 101905. [Google Scholar] [CrossRef] [PubMed]
- Erinle, T.J.; MacIsaac, J.; Yang, C.; Adewole, D.I. Effect of red osier dogwood extract on growth performance, blood biochemical parameters, and gut functionality of broiler chickens challenged or unchallenged intraperitoneally with Salmonella Enteritidis lipopolysaccharide. Poult. Sci. 2022, 101, 101861. [Google Scholar] [CrossRef] [PubMed]
- Tonda, R.M.; Rubach, J.K.; Lumpkins, B.S.; Mathis, G.F.; Poss, M.J. Effects of tannic acid extract on performance and intestinal health of broiler chickens following coccidiosis vaccination and/or a mixed-species Eimeria challenge. Poult. Sci. 2018, 97, 3031–3042. [Google Scholar] [CrossRef]
- Engström, M.T.; Sun, X.; Suber, M.P.; Li, M.; Salminen, J.P.; Hagerman, A.E. The Oxidative Activity of Ellagitannins Dictates Their Tendency To Form Highly Stabilized Complexes with Bovine Serum Albumin at Increased pH. J. Agric. Food Chem. 2016, 64, 8994–9003. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Huang, R.; Wang, N.; Deng, Y.; Tan, B.; Yin, Y.; Qi, M.; Wang, J. Ellagic Acid Alleviates Oxidative Stress by Mediating Nrf2 Signaling Pathways and Protects against Paraquat-Induced Intestinal Injury in Piglets. Antioxidants 2022, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Abbasalipour, H.; Hajizadeh Moghaddam, A.; Ranjbar, M. Sumac and gallic acid-loaded nanophytosomes ameliorate hippocampal oxidative stress via regulation of Nrf2/Keap1 pathway in autistic rats. J. Biochem. Mol. Toxicol. 2022, 36, e23035. [Google Scholar] [CrossRef]
- Ekambaram, S.P.; Aruldhas, J.; Srinivasan, A.; Erusappan, T. Modulation of NF-κB and MAPK signalling pathways by hydrolysable tannin fraction from Terminalia chebula fruits contributes to its anti-inflammatory action in RAW 264.7 cells. J. Pharm. Pharmacol. 2022, 74, 718–729. [Google Scholar] [CrossRef]
- Anderson, R.C.; Vodovnik, M.; Min, B.R.; Pinchak, W.E.; Krueger, N.A.; Harvey, R.B.; Nisbet, D.J. Bactericidal effect of hydrolysable and condensed tannin extracts on Campylobacter jejuni in vitro. Folia Microbiol. 2012, 57, 253–258. [Google Scholar] [CrossRef]
- Nakashima, H.; Murakami, T.; Yamamoto, N.; Sakagami, H.; Tanuma, S.; Hatano, T.; Yoshida, T.; Okuda, T. Inhibition of human immunodeficiency viral replication by tannins and related compounds. Antiviral. Res. 1992, 18, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yao, L.; Su, J.; Fan, R.; Zheng, J.; Han, Y. Effects of Lactobacillus plantarum and its fermentation products on growth performance, immune function, intestinal pH, and cecal microorganisms of Lingnan yellow chicken. Poult. Sci. 2023, 102, 102610. [Google Scholar] [CrossRef]
- Xing, Y.; Wu, Y.; Mao, C.; Sun, D.; Guo, S.; Xu, Y.; Jin, X.; Yan, S.; Shi, B. Water extract of Artemisia ordosica enhances antioxidant capability and immune response without affecting growth performance in weanling piglets. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Q.; Zhong, D.; Zhang, X.; Li, G.Z. IL-10 targets Th1/Th2 balance in vascular dementia. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5614–5619. [Google Scholar]
- Cai, G.; Wusiman, A.; Gu, P.; Mao, N.; Xu, S.; Zhu, T.; He, J.; Liu, Z.; Wang, D. Supplementation of Alhagi honey polysaccharides contributes to the improvement of the intestinal immunity regulating the structure of intestinal flora in mice. Food Funct. 2021, 12, 9693–9707. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Tang, S.; Zhao, H.; Zhong, R.; Liu, L.; Meng, Q.; Zhang, H.; Chen, L. Combined effects of sodium butyrate and xylo-oligosaccharide on growth performance, anti-inflammatory and antioxidant capacity, intestinal morphology and microbiota of broilers at early stage. Poult. Sci. 2023, 102, 102585. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.X.; Zhang, M.; Yang, R.; Min, Y.; Guo, P.T.; Zhang, J.; Wang, C.K.; Jin, L.; Gao, Y.Y. The anti-inflammatory effect of lutein in broilers is mediated by regulating Toll-like receptor 4/myeloid-differentiation-factor 88 signaling pathway. Poult. Sci. 2023, 102, 102622. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.B.; Jeong, J.Y.; Park, J.Y.; Jun, E.M.; Lee, S.I.; Choe, S.S.; Park, D.Y.; Choi, E.W.; Seen, D.S.; Lim, J.S.; et al. Anti-Arthritic and Analgesic Effect of NDI10218, a Standardized Extract of Terminalia chebula, on Arthritis and Pain Model. Biomol. Ther. 2012, 20, 104–112. [Google Scholar] [CrossRef]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Z.; Liu, M.; Li, E.; Shen, J.; Wang, J.; Liu, W.; Jin, X. The Terminalia chebula Retz extract treats hyperuricemic nephropathy by inhibiting TLR4/MyD88/NF-κB axis. J. Ethnopharmacol. 2024, 322, 117678. [Google Scholar] [CrossRef]
- Wei, C.; Chen, X.; Chen, D.; He, J.; Zheng, P.; Chen, H.; Yan, H.; Yu, B.; Luo, Y.; Huang, Z. Effects of dietary dihydromyricetin supplementation on intestinal barrier and humoral immunity in growing-finishing pigs. Anim. Biotechnol. 2022, 33, 1398–1406. [Google Scholar] [CrossRef]
- Wu, F.; Wang, H.; Li, S.; Wei, Z.; Han, S.; Chen, B. Effects of dietary supplementation with quercetagetin on nutrient digestibility, intestinal morphology, immunity, and antioxidant capacity of broilers. Front. Vet. Sci. 2022, 9, 1060140. [Google Scholar] [CrossRef]
- Long, L.N.; Zhang, H.H.; Wang, F.; Yin, Y.X.; Yang, L.Y.; Chen, J.S. Research Note: Effects of polysaccharide-enriched Acanthopanax senticosus extract on growth performance, immune function, antioxidation, and ileal microbial populations in broiler chickens. Poult. Sci. 2021, 100, 101028. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Li, H.; Qiu, Q.; Wang, Y. Condensed Tannins Improved Immune Functions and Reduced Environmental Pollution of Captive Sichuan Black Goat Kids Under Cadmium Toxicity. Biol. Trace Elem. Res. 2022, 200, 4325–4331. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Xie, K.; Yi, J.; Song, Z.; Zhang, H.; He, X. The effects of magnolol supplementation on growth performance, meat quality, oxidative capacity, and intestinal microbiota in broilers. Poult. Sci. 2022, 101, 101722. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Jin, X.; Camerlink, I.; Tong, M.; Su, J.; Zhao, F.; Yan, S.; Shi, B. Effects of Yucca schidigera extract on growth performance and antioxidative function of small intestine in broilers. J. Anim. Physiol. Anim. Nutr. 2019, 103, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Messaili, S.; Colas, C.; Fougère, L.; Destandau, E. Combination of molecular network and centrifugal partition chromatography fractionation for targeting and identifying Artemisia annua L. antioxidant compounds. J. Chromatogr. A 2020, 1615, 460785. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Zhu, G.Y.; Sheng, X.H.; Xing, K.; Venema, K.; Wang, X.G.; Xiao, L.F.; Guo, Y.; Ni, H.M.; Zhu, N.H.; et al. Dietary supplementation with selenomethionine enhances antioxidant capacity and selenoprotein gene expression in layer breeder roosters. Poult. Sci. 2022, 101, 102113. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.K.; Kim, H.G.; Han, J.M.; Lee, J.S.; Lee, J.S.; Chung, S.H.; Son, C.G. Hepatoprotective Effect of Terminalia chebula against t-BHP-Induced Acute Liver Injury in C57/BL6 Mice. Evid. Based Complement. Alternat. Med. 2015, 2015, 517350. [Google Scholar] [CrossRef]
- Ahmadi-Naji, R.; Heidarian, E.; Ghatreh-Samani, K. Evaluation of the effects of the hydroalcoholic extract of Terminalia chebula fruits on diazinon-induced liver toxicity and oxidative stress in rats. Avicenna J. Phytomed. 2017, 7, 454–466. [Google Scholar] [PubMed]
- Farina, M.; Vieira, L.E.; Buttari, B.; Profumo, E.; Saso, L. The Nrf2 Pathway in Ischemic Stroke: A Review. Molecules 2021, 26, 5001. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Xiong, Q.; Lin, K.; Liang, Z.; Zhou, M.; Tian, X.; Xu, C.; Ru, Q. Terminalia chebula extracts ameliorate methamphetamine-induced memory deficits via activating the ERK and Nrf2 pathway. Brain Res. Bull. 2022, 184, 76–87. [Google Scholar] [CrossRef]
- Lin, K.; Zhou, M.; Leng, C.; Tao, X.; Zhou, R.; Li, Y.; Sun, B.; Shu, X.; Liu, W. Neuroprotective Effect of Polyphenol Extracts from Terminalia chebula Retz. against Cerebral Ischemia-Reperfusion Injury. Molecules 2022, 27, 6449. [Google Scholar] [CrossRef]
- Zhou, R.; Lin, K.; Leng, C.; Zhou, M.; Zhang, J.; Li, Y.; Liu, Y.; Ye, X.; Xu, X.; Sun, B.; et al. Chebulic Acid Prevents Hypoxia Insult via Nrf2/ARE Pathway in Ischemic Stroke. Nutrients 2022, 14, 5390. [Google Scholar] [CrossRef] [PubMed]
- Xun, W.; Shi, L.; Zhou, H.; Hou, G.; Cao, T.; Zhao, C. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic Escherichia coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef]
- Dubrovsky, G.; Dunn, J.C.Y. Mechanisms for intestinal regeneration. Curr. Opin. Pediatr. 2018, 30, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, R.; Liu, Q.; Gong, Y.; Ou, Y.; Qi, Q.; Xie, Y.; Wang, X.; Hu, C.; Jiang, S.; et al. Effects of dietary supplementation with dandelion tannins or soybean isoflavones on growth performance, antioxidant function, intestinal morphology, and microbiota composition in Wenchang chickens. Front. Vet. Sci. 2023, 9, 1073659. [Google Scholar] [CrossRef]
- Jing, C.; Niu, J.; Liu, Y.; Jiao, N.; Huang, L.; Jiang, S.; Yan, L.; Yang, W.; Li, Y. Tannic Acid Extracted from Galla chinensis Supplementation in the Diet Improves Intestinal Development through Suppressing Inflammatory Responses via Blockage of NF-κB in Broiler Chickens. Animals 2022, 12, 2397. [Google Scholar] [CrossRef]
- Song, B.; Li, P.; Yan, S.; Liu, Y.; Gao, M.; Lv, H.; Lv, Z.; Guo, Y. Effects of Dietary Astragalus Polysaccharide Supplementation on the Th17/Treg Balance and the Gut Microbiota of Broiler Chickens Challenged With Necrotic Enteritis. Front. Immunol. 2022, 13, 781934. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, X.; Wang, Y.; Guo, Y.; Zhu, P.; Li, G.; Zhang, J.; Ma, Q.; Zhao, L. Dietary ellagic acid ameliorated Clostridium perfringens-induced subclinical necrotic enteritis in broilers via regulating inflammation and cecal microbiota. J. Anim. Sci. Biotechnol. 2022, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Meimandipour, A.; Shuhaimi, M.; Soleimani, A.F.; Azhar, K.; Hair-Bejo, M.; Kabeir, B.M.; Javanmard, A. Muhammad Anas, and A. M. Yazid. Selected microbial groups and short-chain fatty acids profile in a simulated chicken cecum supplemented with two strains of Lactobacillus. Poult. Sci. 2010, 89, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, J.; Chen, X.; Nie, C.; Zhao, J.; Guan, W.; Lei, L.; He, T.; Chen, Y.; Johnston, L.J.; et al. Combination of Clostridium butyricum and Corn Bran Optimized Intestinal Microbial Fermentation Using a Weaned Pig Model. Front. Microbiol. 2018, 9, 3091. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; Boyen, F.; Gantois, I.; Timbermont, L.; Bohez, L.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. Supplementation of coated butyric acid in the feed reduces colonization and shedding of Salmonella in poultry. Poult. Sci. 2005, 84, 1851–1856. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–12. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Vital, M.; Howe, A.C.; Tiedje, J.M. Revealing the bacterial butyrate synthesis pathways by analyzing (meta)genomic data. mBio 2014, 5, e00889. [Google Scholar] [CrossRef]
- Pandit, R.J.; Hinsu, A.T.; Patel, N.V.; Koringa, P.G.; Jakhesara, S.J.; Thakkar, J.R.; Shah, T.M.; Limon, G.; Psifidi, A.; Guitian, J.; et al. Microbial diversity and community composition of caecal microbiota in commercial and indigenous Indian chickens determined using 16s rDNA amplicon sequencing. Microbiome 2018, 6, 115. [Google Scholar] [CrossRef]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken Gut Microbiota: Importance and Detection Technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Hu, J.; Cheng, H.W. Research Note: Probiotic, Bacillus subtilis, alleviates neuroinflammation in the hippocampus via the gut microbiota-brain axis in heat-stressed chickens. Poult. Sci. 2023, 102, 102635. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Zhou, T.; Ma, P.; Xie, B.; Li, W.; Gong, S.; Xue, F. Mechanism determination on the interactive effects between host immunity and gut microbiome to resist consecutive hydrogen sulfide inhalation of laying hens. Poult. Sci. 2023, 102, 102694. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Zhang, L.; Zhang, B.; Kong, L.; Pan, X.; Goossens, T.; Song, Z. Dietary sodium butyrate improves female broiler breeder performance and offspring immune function by enhancing maternal intestinal barrier and microbiota. Poult. Sci. 2023, 102, 102658. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Pedroso, A.A.; Mallo, J.J.; Puyalto, M.; Kim, W.K.; Applegate, T.J. Sodium butyrate improved performance while modulating the cecal microbiota and regulating the expression of intestinal immune-related genes of broiler chickens. Poult. Sci. 2017, 96, 3981–3993. [Google Scholar] [CrossRef]
- Goel, A.; Kim, B.J.; Ncho, C.M.; Jeong, C.M.; Gupta, V.; Jung, J.Y.; Ha, S.Y.; Lee, D.H.; Yang, J.K.; Choi, Y.H. Dietary Supplementation of Shredded, Steam-Exploded Pine Particles Decreases Pathogenic Microbes in the Cecum of Acute Heat-Stressed Broilers. Animals 2021, 11, 2252. [Google Scholar] [CrossRef] [PubMed]
- Wexler, A.G.; Goodman, A.L. An insider’s perspective: Bacteroides as a window into the microbiome. Nat. Microbiol. 2017, 2, 17026. [Google Scholar] [CrossRef]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef]
- Zhang, H.; Li, M.; Zhang, K.; Ding, X.; Bai, S.; Zeng, Q.; Chu, L.; Hou, D.; Xuan, Y.; Yin, H.; et al. Effect of benzoic acid, Enterococcus faecium, and essential oil complex on intestinal microbiota of laying hens under coccidia and Clostridium perfringens challenge. Poult. Sci. 2023, 102, 102490. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Piao, M.; Song, Y. Dietary Quercetin Increases Colonic Microbial Diversity and Attenuates Colitis Severity in Citrobacter rodentium-Infected Mice. Front. Microbiol. 2019, 10, 1092. [Google Scholar] [CrossRef] [PubMed]
- Wassie, T.; Cheng, B.; Zhou, T.; Gao, L.; Lu, Z.; Xie, C.; Wu, X. Microbiome-metabolome analysis reveals alterations in the composition and metabolism of caecal microbiota and metabolites with dietary Enteromorpha polysaccharide and Yeast glycoprotein in chickens. Front. Immunol. 2022, 13, 996897. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Diets |
|---|---|
| Control group | basal diet |
| Treatment group 1 | basal diet + 200 mg/kg |
| Treatment group 2 | basal diet + 400 mg/kg |
| Treatment group 3 | basal diet + 600 mg/kg |
| Items 1 | Age (Days) | |
|---|---|---|
| Starter | Finisher | |
| (1 to 28 Days) | (29 to 56 Days) | |
| Ingredient (%) | ||
| Corn | 55.57 | 66.40 |
| Wheat bran | 0.60 | 0.50 |
| Soybean meal | 35.00 | 22.51 |
| Soya oil | 2.62 | 1.94 |
| Fish meal | 1.40 | 5.00 |
| HCl-lysine | 0.10 | 0.02 |
| DL-methionine | 0.30 | 0.09 |
| Limestone | 1.14 | 0.82 |
| Dicalcium phosphate | 1.96 | 1.43 |
| Sodium chloride | 0.3 | 0.3 |
| Vitamin–mineral premix 2 | 1.00 | 1.00 |
| Nutrient content 3 (%) | ||
| ME (MJ/kg) | 12.12 | 12.55 |
| CP | 21.18 | 19.00 |
| Ca | 1.00 | 0.90 |
| Total P | 0.74 | 0.70 |
| Available P | 0.45 | 0.45 |
| Digestible Lys | 1.24 | 1.03 |
| Digestible Met | 0.62 | 0.43 |
| Parameters 2 | TCE Level 3 (mg/kg) | SEM 4 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | Anova | Linear | Quadratic | ||
| Initial BW (g) | 31.17 | 31.04 | 31.08 | 31.22 | 0.130 | 0.869 | 0.581 | 0.723 |
| Final BW (kg) | 1.76 c | 1.91 b | 1.84 a | 1.86 a | 0.014 | <0.001 | 0.049 | 0.004 |
| Starter period, d 1 to 28 | ||||||||
| ADG (g) | 19.15 | 21.30 | 20.36 | 20.09 | 0.300 | 0.079 | 0.499 | 0.100 |
| ADFI (g) | 36.76 | 38.51 | 38.00 | 37.88 | 0.415 | 0.526 | 0.452 | 0.412 |
| F/G (feed/gain, g/g) | 1.90 | 1.84 | 1.83 | 1.85 | 0.016 | 0.331 | 0.205 | 0.177 |
| Finisher period, d 29 to 56 | ||||||||
| ADG (g) | 43.21 b | 44.21 a | 44.20 a | 44.75 a | 0.152 | <0.001 | <0.001 | <0.001 |
| ADFI (g) | 99.77 b | 106.54 a | 99.43 b | 103.63 ab | 0.902 | 0.006 | 0.591 | 0.683 |
| F/G (feed/gain, g/g) | 2.35 | 2.32 | 2.25 | 2.29 | 0.015 | 0.092 | 0.050 | 0.079 |
| Overall, from d 1 to 56 | ||||||||
| ADG (g) | 30.82 c | 33.61 b | 32.28 a | 32.71 a | 0.249 | <0.001 | 0.049 | 0.004 |
| ADFI (g) | 69.24 b | 72.35 a | 68.32 b | 70.34 ab | 0.488 | 0.012 | 0.872 | 0.855 |
| F/G (feed/gain, g/g) | 2.22 b | 2.15 a | 2.13 a | 2.15 a | 0.009 | <0.001 | 0.002 | <0.001 |
| Parameters 2 | TCE Level 3 (mg/kg) | SEM 4 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | Anova | Linear | Quadratic | ||
| Serum | ||||||||
| IgA (g/L) | 0.26 | 0.27 | 0.28 | 0.28 | 0.078 | 0.670 | 0.204 | 0.451 |
| IgM (g/L) | 0.69 | 0.70 | 0.78 | 0.77 | 0.017 | 0.103 | 0.028 | 0.086 |
| IgG (g/L) | 1.97 | 1.96 | 1.99 | 2.05 | 0.054 | 0.943 | 0.586 | 0.820 |
| IL-1β (pg/mL) | 76.89 | 68.99 | 58.98 | 62.01 | 3.345 | 0.241 | 0.066 | 0.134 |
| IL-2 (pg/mL) | 260.02 | 284.77 | 288.27 | 286.44 | 6.805 | 0.435 | 0.179 | 0.256 |
| IL-4 (pg/mL) | 48.18 | 51.28 | 54.76 | 55.29 | 1.261 | 0.142 | 0.021 | 0.068 |
| IL-6 (pg/mL) | 49.04 b | 46.77 b | 39.09 a | 38.20 a | 1.263 | <0.001 | <0.001 | <0.001 |
| IL-10 (pg/mL) | 120.24 b | 133.01 ab | 143.37 a | 141.90 ab | 3.883 | 0.125 | 0.026 | 0.055 |
| Liver | ||||||||
| IgA (μg/mL prot) | 28.32 | 30.16 | 31.26 | 31.43 | 0.857 | 0.582 | 0.176 | 0.367 |
| IgM (μg/mL prot) | 85.96 | 86.63 | 89.48 | 88.75 | 1.648 | 0.873 | 0.460 | 0.750 |
| IgG (μg/mL prot) | 177.24 | 178.35 | 202.53 | 184.52 | 4.809 | 0.137 | 0.294 | 0.360 |
| IL-1β (pg/mg prot) | 12.27 a | 11.90 ab | 11.55 ab | 10.9 b | 0.182 | 0.057 | 0.005 | 0.021 |
| IL-2 (pg/mg prot) | 39.09 b | 39.88 ab | 41.25 ab | 44.43 a | 0.822 | 0.095 | 0.014 | 0.039 |
| IL-4 (pg/mg prot) | 6.70 b | 7.34 ab | 7.41 ab | 8.13 a | 0.172 | 0.021 | 0.002 | 0.011 |
| IL-6 (pg/mg prot) | 4.59 a | 4.10 b | 4.23 ab | 4.10 b | 0.076 | 0.062 | 0.049 | 0.068 |
| IL-10 (pg/mg prot) | 10.43 | 10.55 | 10.96 | 11.09 | 0.187 | 0.563 | 0.155 | 0.373 |
| Spleen | ||||||||
| IgA (μg/mL prot) | 25.00 | 26.98 | 27.07 | 27.04 | 0.759 | 0.745 | 0.371 | 0.549 |
| IgM (μg/mL prot) | 77.16 b | 77.45 b | 80.81 ab | 82.35 a | 0.802 | 0.042 | 0.005 | 0.021 |
| IgG (ug/mL prot) | 217.43 b | 240.19 a | 241.29 a | 238.54 a | 3.714 | 0.059 | 0.050 | 0.027 |
| IL-1β (pg/mg prot) | 13.80 a | 12.23 ab | 10.53 b | 10.30 b | 0.528 | 0.052 | 0.006 | 0.021 |
| IL-2 (pg/mg prot) | 37.18 | 37.82 | 38.15 | 37.62 | 0.339 | 0.808 | 0.600 | 0.618 |
| IL-4 (pg/mg prot) | 6.34 | 6.74 | 7.24 | 6.98 | 0.193 | 0.423 | 0.411 | 0.244 |
| IL-6 (pg/mg prot) | 4.10 a | 3.21 b | 3.38 b | 3.27 b | 0.103 | 0.002 | 0.008 | 0.002 |
| IL-10 (pg/mg prot) | 9.73 b | 11.11 ab | 11.71 a | 11.52 a | 0.288 | 0.052 | 0.017 | 0.019 |
| Parameters 2 | TCE Level 3 (mg/kg) | SEM 4 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | Anova | Linear | Quadratic | ||
| Serum | ||||||||
| CAT (U/mL) | 16.13 | 16.93 | 18.79 | 17.11 | 0.403 | 0.114 | 0.189 | 0.124 |
| GSH-Px (U/mL) | 950.83 b | 1048.39 ab | 1213.82 a | 1228.17 a | 39.343 | 0.021 | 0.002 | 0.009 |
| T-AOC (U/mL) | 0.49 b | 0.50 b | 0.52 ab | 0.55 a | 0.007 | 0.007 | <0.001 | 0.002 |
| MDA (nmol/mL) | 2.95 | 2.80 | 2.90 | 3.02 | 0.097 | 0.717 | 0.731 | 0.767 |
| SOD (U/mL) | 55.99 | 61.36 | 57.04 | 65.32 | 3.060 | 0.902 | 0.399 | 0.688 |
| Liver | ||||||||
| CAT (U/mg) | 59.37 | 68.31 | 78.87 | 70.14 | 5.027 | 0.623 | 0.352 | 0.451 |
| GSH-Px (U/mg) | 55.94 | 60.51 | 62.48 | 61.80 | 3.079 | 0.892 | 0.490 | 0.728 |
| T-AOC (U/mg) | 0.57b | 0.64 a | 0.64 a | 0.65 a | 0.011 | 0.017 | 0.010 | 0.007 |
| MDA (nmol/mg) | 0.57 | 0.55 | 0.54 | 0.47 | 0.038 | 0.818 | 0.359 | 0.637 |
| SOD (U/mg) | 14.98 | 16.29 | 16.20 | 17.76 | 0.530 | 0.342 | 0.081 | 0.225 |
| Spleen | ||||||||
| CAT (U/mg) | 12.10 b | 13.69 ab | 14.32 ab | 17.24 a | 0.667 | 0.037 | 0.004 | 0.016 |
| GSH-Px (U/mg) | 96.05 b | 108.62 b | 117.68 ab | 135.94 a | 4.585 | 0.008 | <0.001 | 0.003 |
| T-AOC (U/mg) | 0.13 | 0.14 | 0.19 | 0.27 | 0.028 | 0.313 | 0.066 | 0.161 |
| MDA (nmol/mg) | 0.98 | 0.84 | 0.77 | 0.87 | 0.035 | 0.177 | 0.194 | 0.085 |
| SOD (U/mg) | 8.65 | 10.05 | 12.16 | 12.04 | 0.643 | 0.157 | 0.029 | 0.080 |
| Parameters 2 | TCE Level 3 (mg/kg) | SEM 4 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | Anova | Linear | Quadratic | ||
| Duodenum | ||||||||
| VH (μm) | 826.56 b | 831.74 b | 912.78 a | 862.22 b | 9.976 | 0.002 | 0.032 | 0.033 |
| CD (μm) | 140.65 b | 132.81 b | 109.99 a | 111.07 a | 4.051 | 0.004 | <0.001 | 0.003 |
| VH/CD (μm: μm) | 5.67 b | 6.14 b | 7.83 a | 7.74 a | 0.301 | 0.008 | 0.002 | 0.006 |
| Jejunum | ||||||||
| VH (μm) | 787.15 | 802.01 | 814.81 | 794.68 | 27.894 | 0.989 | 0.891 | 0.947 |
| CD (μm) | 98.13 | 90.99 | 90.63 | 89.06 | 4.137 | 0.886 | 0.469 | 0.734 |
| VH/CD (μm: μm) | 8.18 | 8.84 | 9.31 | 9.06 | 0.289 | 0579 | 0.236 | 0.373 |
| Ileum | ||||||||
| VH (μm) | 448.25 b | 545.92 b | 683.10 a | 496.95 b | 26.746 | 0.005 | 0.245 | 0.008 |
| CD (μm) | 67.61 | 61.92 | 60.54 | 59.95 | 2.636 | 0.750 | 0.312 | 0.543 |
| VH/CD (μm: μm) | 6.67 c | 8.93 b | 11.78 a | 8.47 bc | 0.480 | <0.001 | 0.053 | <0.001 |
| Parameters 2 | TCE Level 3 (mg/kg) | SEM 4 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | Anova | Linear | Quadratic | ||
| Acetic acid (mg/g) | 2.46 | 2.48 | 2.68 | 2.64 | 0.038 | 0.084 | 0.026 | 0.084 |
| Propionic acid (mg/g) | 0.59 | 0.78 | 0.77 | 0.64 | 0.045 | 0.350 | 0.760 | 0.189 |
| Butyric acid (mg/g) | 0.43 | 0.46 | 0.60 | 0.62 | 0.038 | 0.200 | 0.037 | 0.119 |
| Isobutyric acid (mg/g) | 0.14 | 0.11 | 0.10 | 0.10 | 0.007 | 0.254 | 0.063 | 0.127 |
| Valeric acid (mg/g) | 0.16 | 0.15 | 0.12 | 0.13 | 0.011 | 0.538 | 0.156 | 0.350 |
| Isovaleric acid (mg/g) | 0.16 b | 0.14 a | 0.15 ab | 0.16 b | 0.003 | 0.035 | 0.920 | 0.042 |
| Total SCFAs (mg/g) | 3.95 | 4.13 | 4.43 | 4.28 | 0.093 | 0.301 | 0.115 | 0.196 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.; Liu, S.; Wang, F.; Wang, T.; Yin, L.; Chen, J.; Fu, C. Effects of Dietary Terminalia chebula Extract on Growth Performance, Immune Function, Antioxidant Capacity, and Intestinal Health of Broilers. Animals 2024, 14, 746. https://doi.org/10.3390/ani14050746
Cheng Y, Liu S, Wang F, Wang T, Yin L, Chen J, Fu C. Effects of Dietary Terminalia chebula Extract on Growth Performance, Immune Function, Antioxidant Capacity, and Intestinal Health of Broilers. Animals. 2024; 14(5):746. https://doi.org/10.3390/ani14050746
Chicago/Turabian StyleCheng, Ying, Shida Liu, Fang Wang, Tao Wang, Lichen Yin, Jiashun Chen, and Chenxing Fu. 2024. "Effects of Dietary Terminalia chebula Extract on Growth Performance, Immune Function, Antioxidant Capacity, and Intestinal Health of Broilers" Animals 14, no. 5: 746. https://doi.org/10.3390/ani14050746