
Difructose Anhydride and Passive Immunity Effects on Passive Immune Transfer and Performance of Feeding Difructose Anhydride to Neonatal Calves

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
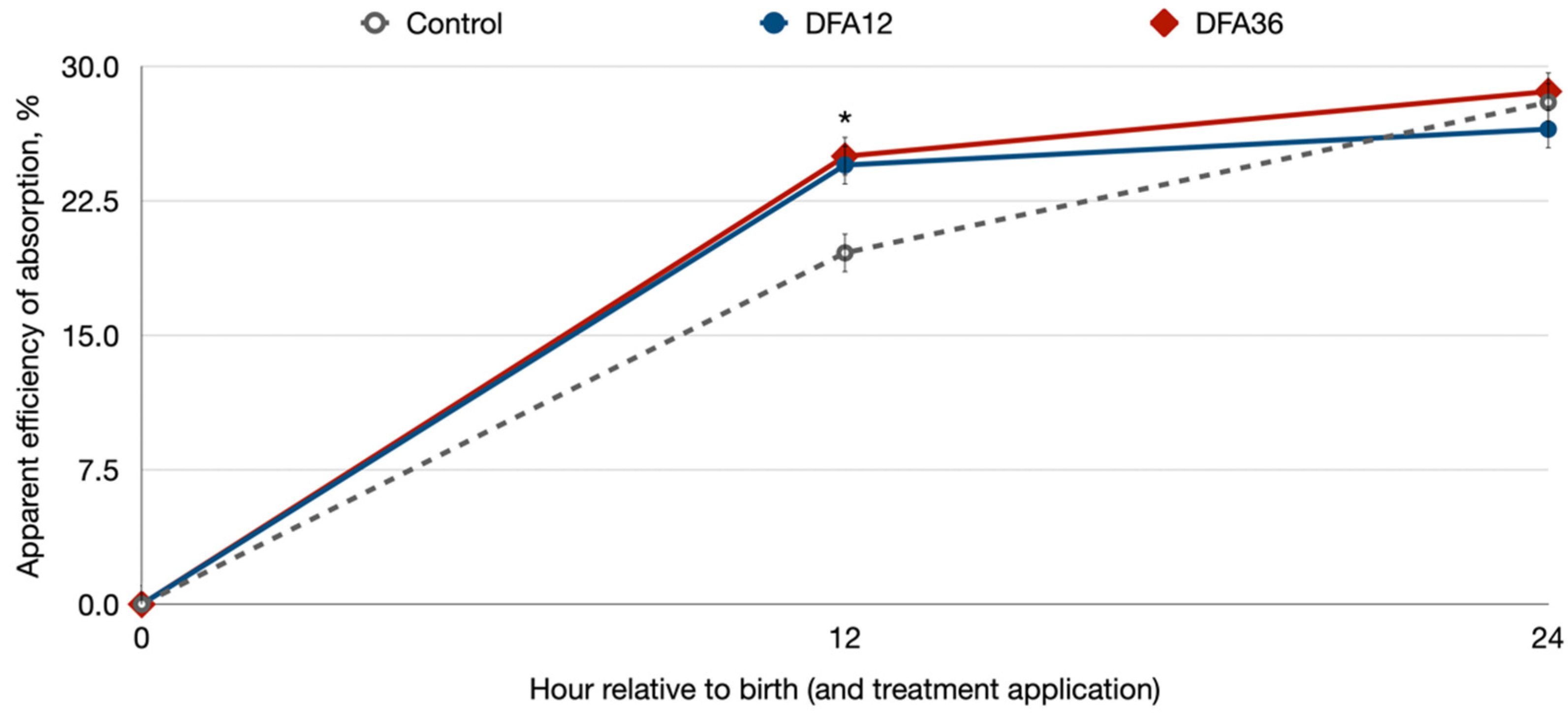
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Beam, A.L.; Lombard, J.E.; Kopral, C.A.; Garber, L.P.; Winter, A.L.; Hicks, J.A.; Schlater, J.L. Prevalence of failure of passive transfer of immunity in newborn heifer calves and associated management practices on US dairy operations. J. Dairy Sci. 2009, 92, 3973–3980. [Google Scholar] [CrossRef]
- Panousis, N.; Kritsepi-Konstantinou, M.; Kalaitzakis, E.; Giadinis, N.; Valergakis, G.E. Prevalence of failure of passive transfer of immunoglobulins in Holstein calves in Northern Greece and association with management practices. J. Hell. Vet. Med. Soc. 2013, 64, 193–200. [Google Scholar] [CrossRef]
- Staněk, S.; Nejedlá, E.; Fleischer, P.; Pechová, A.; Šlosárková, S. Prevalence of failure of passive transfer of immunity in dairy calves in the Czech Republic. Acta Univ. Agric. Silvic. Mendel. Brun. 2019, 67, 163–172. [Google Scholar] [CrossRef]
- NAHMS. Dairy Cattle Management Practices in the United States; USDA APHIS Veterinary Services: Ft. Collins, CO, USA, 2014.
- McGuirk, S.M.; Collins, M. Managing the production, storage and delivery of colostrum. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 593–603. [Google Scholar] [CrossRef]
- Fischer, A.J.; Song, Y.; He, Z.; Haines, D.M.; Guan, L.L.; Steele, M.A. Effect of delaying colostrum feeding on passive trans- fer and intestinal bacterial colonization in neonatal male Holstein calves. J. Dairy Sci. 2018, 101, 3099–3109. [Google Scholar] [CrossRef]
- Lateur-Rowet, H.J.M.; Breukink, H.J. The failure of the oesophageal groove reflex, when fluids are given with an oesophageal feeder to newborn and young calves. Vet. Q. 1983, 5, 68–74. [Google Scholar] [CrossRef]
- Chapman, H.W.; Butler, D.G.; Newell, M. The route of liquids administered to calves by esophageal feeder. Can. J. Vet. Res. 1986, 50, 84–87. [Google Scholar]
- Lopez, A.J.; Yohe, T.T.; Echeverry-Munera, J.; Nagorske, M.; Renaud, D.L.; Steele, M.A. Effects of a low- or high-frequency colostrum feeding protocol on immunoglobulin G absorption in newborn calves. J. Dairy Sci. 2022, 105, 6318–6326. [Google Scholar] [CrossRef]
- Adams, G.D.; Bush, L.J.; Horner, J.L.; Staley, T.E. Two methods for administering colostrum to newborn calves. J. Dairy Sci. 1985, 68, 773–775. [Google Scholar] [CrossRef]
- Besser, T.E.; Gay, C.C.; Pritchett, L. Comparison of three methods of feeding colostrum to dairy calves. J. Am. Vet. Med. Assoc. 1991, 198, 419–442. [Google Scholar]
- Godden, S.M.; Haines, D.M.; Konkol, K.; Peterson, J. Improving passive transfer of immunoglobulins in calves. II: Interaction between feeding method and volume of colostrum fed. J. Dairy Sci. 2009, 92, 1758–1764. [Google Scholar] [CrossRef]
- Lazarevic, M.; Spring, P.; Shabanovic, M.; Tokic, V.; Tucker, L.A. Effect of gut active carbohydrates on plasma IgG concentrations in piglets and calves. Animal 2010, 4, 938–943. [Google Scholar] [CrossRef]
- Kamada, H.; Nonaka, I.; Ueda, Y.; Murai, M. Selenium addition to colostrum increases immunoglobulin G absorption by new- born calves. J. Dairy Sci. 2007, 90, 5665–5670. [Google Scholar] [CrossRef]
- Morrill, K.M.; Marston, S.P.; Whitehouse, N.L.; Van Amburgh, M.E.; Schwab, C.G.; Haines, D.M.; Erickson, P.S. Anionic salts in the prepartum diet and addition of sodium bicarbonate to colostrum replacer, and their effects on immunoglobulin G absorption in the neonate. J. Dairy Sci. 2010, 93, 2067–2075. [Google Scholar] [CrossRef]
- Pierce, A.E.; Feinstein, A. Biophysical and immunological studies on bovine immune globulins with evidence for selective transport within the mammary gland from maternal plasma to colostrum. Immunology 1965, 8, 106–123. [Google Scholar]
- Balfour, W.E.; Comline, R.S. The specificity of the intestinal absorption of large molecules by the new-born calf. J. Physiol. 1959, 148, 77–78. [Google Scholar]
- Staley, T.E.; Corley, L.D.; Bush, L.J.; Jones, E.W. The ultrastructure of neonatal calf intestine and absorption of heterologous proteins. Anat. Rec. 1972, 172, 559–579. [Google Scholar] [CrossRef]
- Jochims, K.; Kaup, F.-J.; Drommer, W.; Pickel, M. An immunoelectron microscopic investigation of colostral IgG absorption across the intestine of newborn calves. Res. Vet. Sci. 1994, 57, 75–80. [Google Scholar] [CrossRef]
- Kikuchi, H.; Nagura, T.; Inoue, M.; Kishida, T.; Sakurai, H.; Yokota, A.; Asano, K.; Tomita, F.; Sayama, K.; Senba, Y. Physical, chemical and physiological properties of difructose anhydride III produced from inulin by enzymatic reaction. J. Appl. Glycosci. 2004, 51, 291–296. [Google Scholar] [CrossRef]
- Suzuki, T.; Hara, H. Difructose anhydride III and sodium caprate activate paracellular transport via different intracellular events in Caco-2 cells. Life Sci. 2006, 79, 401–410. [Google Scholar] [CrossRef]
- Sato, T.; Hanada, M.; Ohtani, M.; Nakai, T.; Teramura, M.; Sadoya, H.; Takahashi, T.; Hongo, A. Short communication: Effect of difructose anhydride III on serum immunoglobulin G concentration in newborn calves. J. Dairy Sci. 2012, 95, 5336–5339. [Google Scholar] [CrossRef] [PubMed]
- Htun, A.; Sato, T.; Fukuma, N.; Hanada, M. Effects of difructose anhydride III on serum immunoglobulin G concentration and health status of newborn Holstein calves during the preweaning period. J. Dairy Sci. 2018, 101, 3226–3232. [Google Scholar] [CrossRef] [PubMed]
- Quigley, J.D.; Drewry, J.J.; Martine, K.R. Estimation of plasma volume in Holstein and Jersey calves. J. Dairy Sci. 1998, 81, 1308–1312. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.; Tyler, J.W.; Chigerwe, M.; Dawes, M.E.; Middleton, R. Effect of delayed colostrum collection on colostral IgG concentration in dairy cows. J. Am. Vet. Med. Assoc. 2005, 226, 1375–1377. [Google Scholar] [CrossRef] [PubMed]
- Conneely, M.; Berry, D.P.; Sayers, R.; Murphy, J.P.; Lorenz, I.; Doherty, M.L.; Kennedy, E. Factors associated with the concentration of immunoglobulin G in the colostrum of dairy cows. Animal 2013, 7, 1824–1832. [Google Scholar] [CrossRef]
- Kekana, T.W.; Marume, U.; Nherera-Chokuda, F.V. Prepartum supplementation of Moringa oleifera leaf meal: Effects on health of the dam, colostrum quality, and acquisition of immunity in the calf. J. Dairy Sci. 2022, 105, 5813–5821. [Google Scholar] [CrossRef]
- Quigley, J.D.; Kost, C.J.; Wolfe, T.M. Absorption of protein and igg in calves fed a colostrum supplement or replacer. J. Dairy Sci. 2002, 85, 1243–1248. [Google Scholar] [CrossRef]
- Gelsinger, S.L.; Heinrichs, A.J. Comparison of immune responses in calves fed heat-treated or unheated colostrum. J. Dairy Sci. 2017, 100, 4090–4101. [Google Scholar] [CrossRef]
- Besser, T.E.; Garmedia, A.E.; McGuire, T.C.; Gay, C.C. Effect of colostral immunoglobulin G1 and immunoglobulin M concentrations on immunoglobulin absorption in calves. J. Dairy Sci. 1985, 68, 2033–2037. [Google Scholar] [CrossRef]
- Saldana, D.J.; Gelsinger, S.L.; Jones, C.M.; Heinrichs, A.J. Effect of different heating times of high-, medium-, and low-quality colostrum on immunoglobulin G absorption in dairy calves. J. Dairy Sci. 2019, 102, 2068–2074. [Google Scholar] [CrossRef]
- Mineo, H.; Amano, M.; Chiji, H.; Shigematsu, N.; Tomita, F.; Hara, H. Indigestible disaccharides open tight junctions and enhance net calcium, magnesium, and zinc absorption in isolated rat small and large intestinal epithelium. Dig. Dis. Sci. 2004, 49, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Htun, A.; Sato, T.; Hanada, M. Effect of difructose anhy- dride III supplementation on passive immunoglobulin G transfer and serum immunoglobulin G concentration in newborn Holstein calves fed pooled colostrum. J. Dairy Sci. 2016, 99, 5701–5706. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Treatment 1 | p-Value 2 | ||||||
|---|---|---|---|---|---|---|---|
| Control | DFA12 | DFA36 | SE 3 | T | t | Txt | |
| Initial BW, kg | 42.3 | 42.0 | 42.3 | 0.87 | 0.97 | - | - |
| Final BW, kg (42 d of life) | 71.4 | 71.8 | 70.2 | 1.51 | 0.76 | - | - |
| Milk replacer intake, g/d | 888 | 890 | 886 | 1.44 | 0.16 | <0.01 | 0.35 |
| Starter intake, g/d | 274 | 303 | 247 | 32.1 | 0.46 | <0.01 | 0.49 |
| Dry matter intake, g/d | 969 | 1004 | 941 | 38.3 | 0.51 | <0.01 | 0.83 |
| Average daily gain, g/d | 707 | 717 | 690 | 27.6 | 0.78 | <0.01 | 0.92 |
| Feed efficiency, % 4 | 72.9 | 71.9 | 73.5 | 3.35 | 0.93 | <0.01 | 0.81 |
| Treatment 1 | p-Value 2 | ||||||
|---|---|---|---|---|---|---|---|
| Control | DFA12 | DFA36 | SE 3 | T | t | Txt | |
| Colostrum consumed first 24 h, L | 4.39 | 4.28 | 4.25 | 0.16 | 0.80 | - | - |
| Colostrum IgG, g/L 4 | 116.0 | 116.3 | 96.23 | 0.05 | 0.58 | - | - |
| IgG consumption first 24 h, g | 508.9 | 498.0 | 409.2 | 1.12 | 0.77 | - | - |
| Serum IgG, g/L 4 | 28.6 | 29.3 | 29.8 | 1.32 | 0.26 | <0.01 | 0.66 |
| AEA of IgG, % 5 | 23.4 | 25.4 | 26.7 | 1.05 | 0.20 | <0.01 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escartín, M.; Rialp, N.; Bach, A. Difructose Anhydride and Passive Immunity Effects on Passive Immune Transfer and Performance of Feeding Difructose Anhydride to Neonatal Calves. Animals 2024, 14, 35. https://doi.org/10.3390/ani14010035
Escartín M, Rialp N, Bach A. Difructose Anhydride and Passive Immunity Effects on Passive Immune Transfer and Performance of Feeding Difructose Anhydride to Neonatal Calves. Animals. 2024; 14(1):35. https://doi.org/10.3390/ani14010035
Chicago/Turabian StyleEscartín, Miguel, Núria Rialp, and Alex Bach. 2024. "Difructose Anhydride and Passive Immunity Effects on Passive Immune Transfer and Performance of Feeding Difructose Anhydride to Neonatal Calves" Animals 14, no. 1: 35. https://doi.org/10.3390/ani14010035