
Pre-Partum Supplementation with Polyunsaturated Fatty Acids on Colostrum Characteristics and Lamb Immunity and Behavior after a Mild Post-Weaning Aversive Handling Period

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Management Pre-Partum
2.2. Animals, Management and Measurements from Birth until Weaning
2.2.1. Ewes
2.2.2. Lambs
2.3. Animals and Aversive Handling Period (AHP) Post-Weaning
2.4. Behavior Tests Post-AHP
2.5. Inflammatory and Lymphocyte Proliferation Challenge (IC) Post-AHP
2.6. Statistical Analysis
3. Results
3.1. Pre-Partum Concentrate Intake
3.2. Ewe Hematology and Plasmatic Values at Birth
3.3. Birth to Weaning Period
3.3.1. Colostrum Physical Measurements, Chemical and Fatty Acid Composition
3.3.2. Colostrum SCC, Total Solids and IgG Concentration
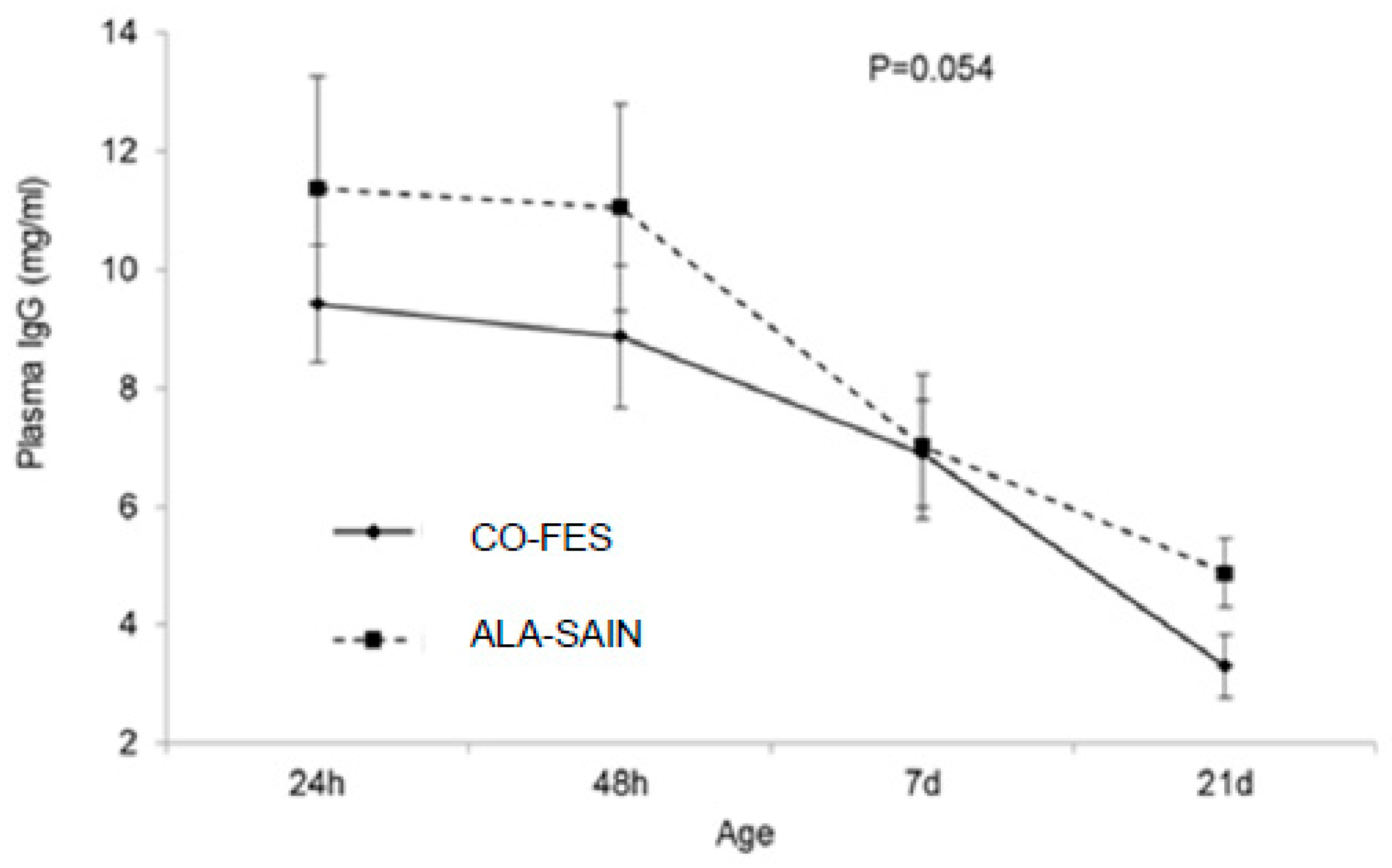
3.3.3. Lamb Serum IgG Concentration and AEA
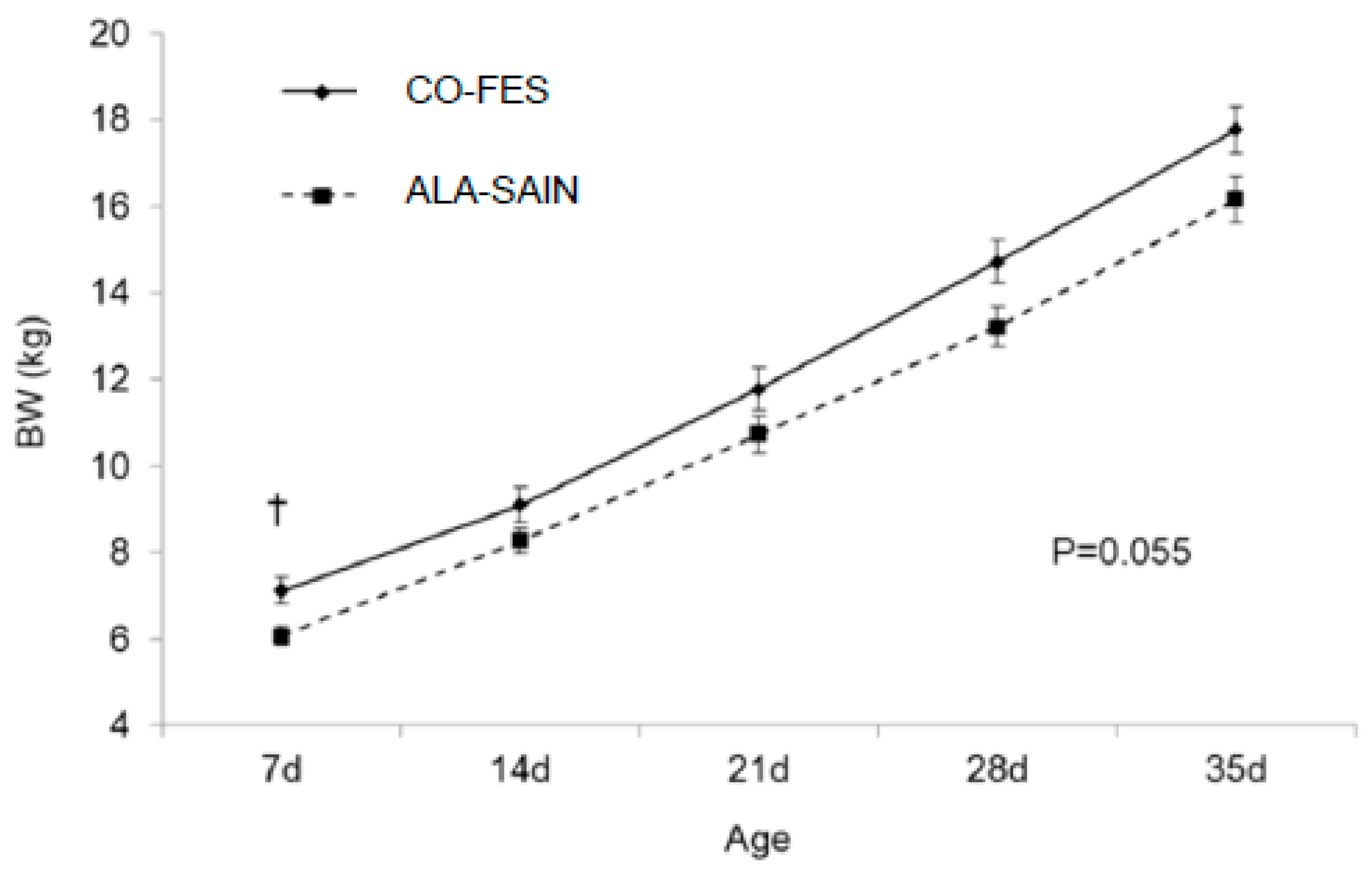
3.3.4. Lamb BW from Birth until Weaning
3.4. Post-Weaning Period
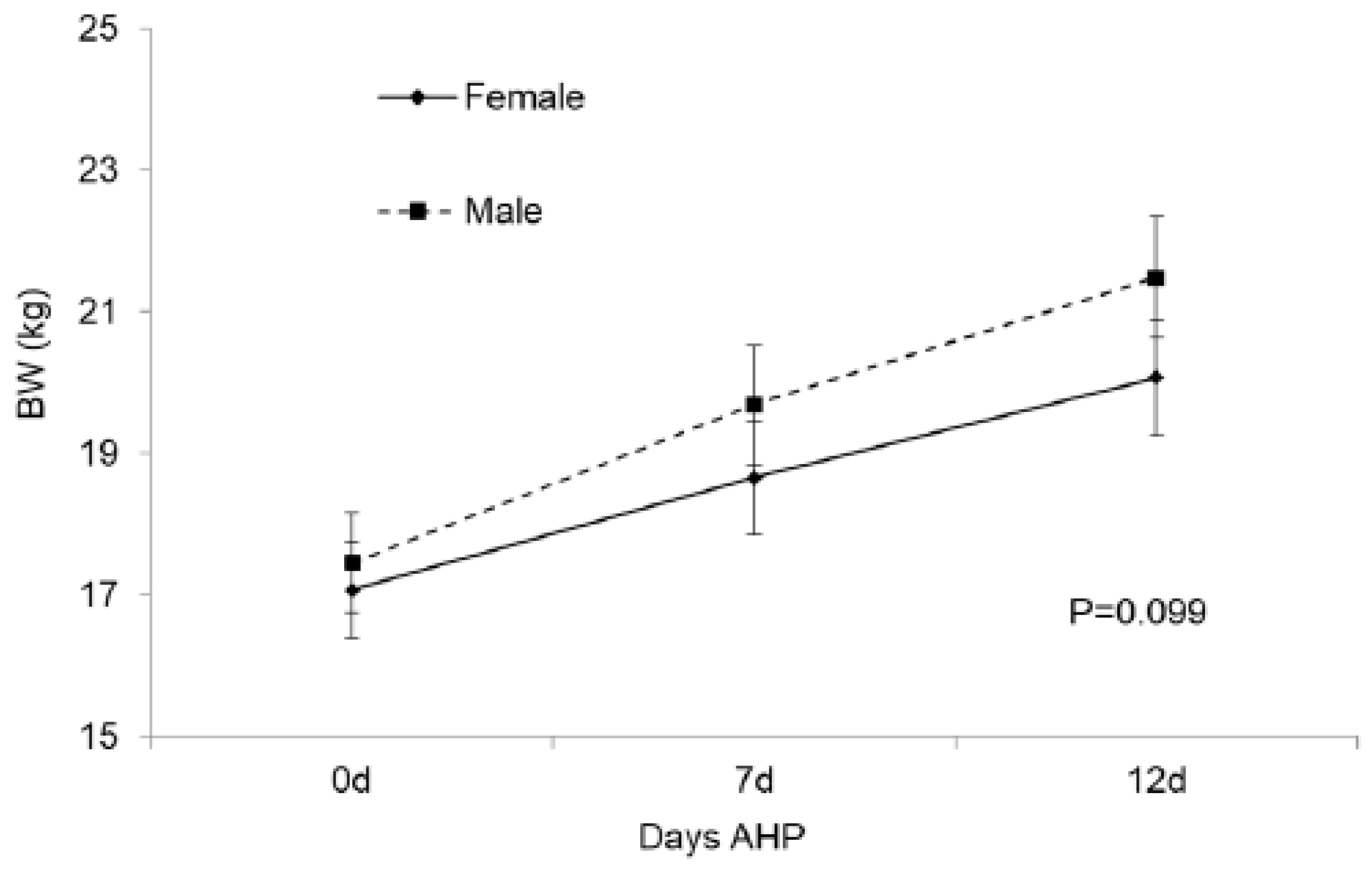
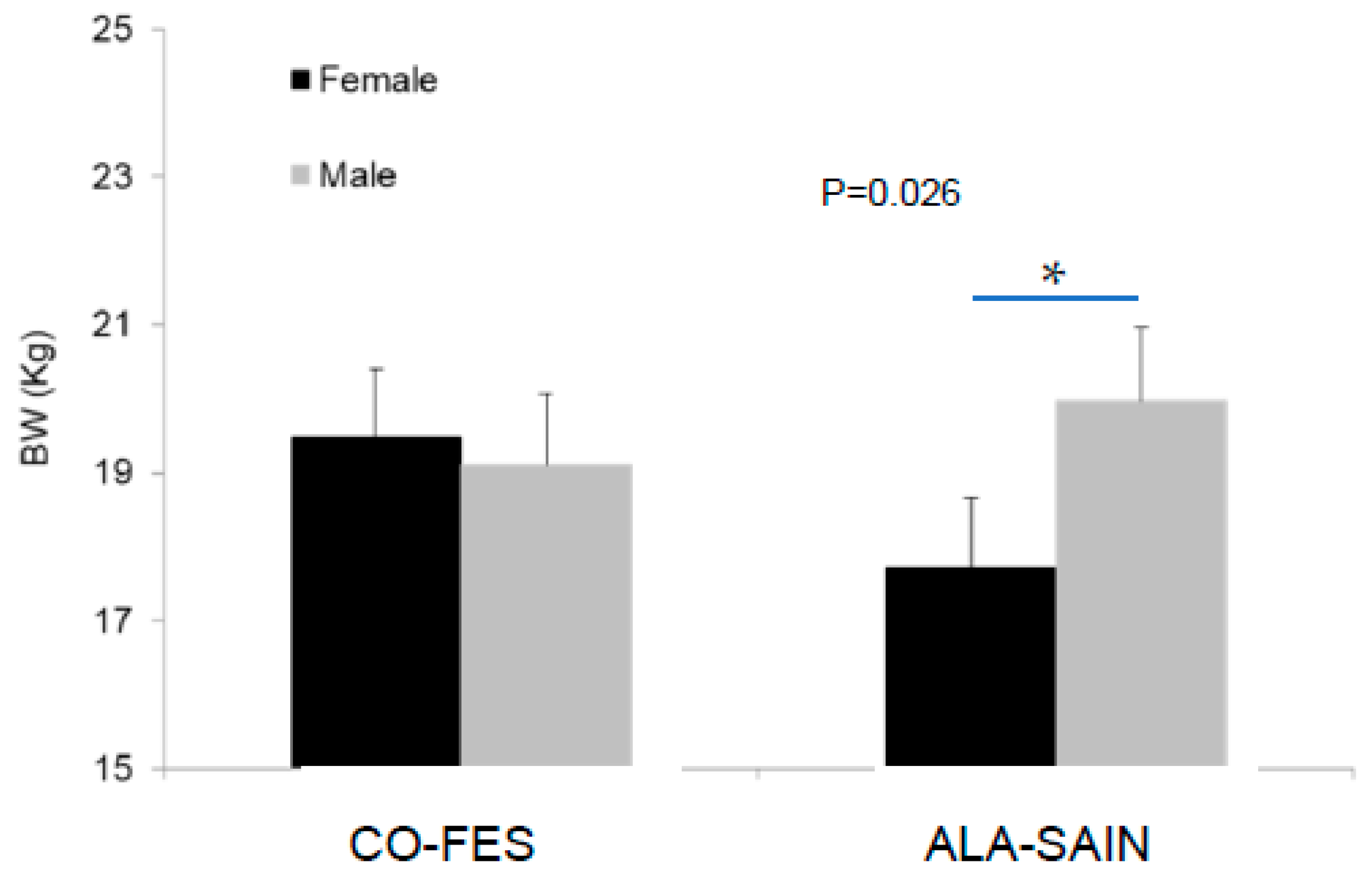
3.4.1. Lamb BW and Plasma Cortisol Concentration during Post-Weaning AHP
3.4.2. Lamb Behavior and Plasma Cortisol Concentrations after Behavior Tests
3.4.3. Inflammatory and Lymphocyte Proliferation Challenge (IC) Post-AHP
4. Discussion
4.1. Ewe Hematology and Plasmatic Values at Lambing
4.2. Colostrum Physical Measurements, Chemical and Fatty Acid Composition
4.3. Colostrum SCC, Total Solids and IgG Concentration
4.4. Lamb Serum IgG Concentration and AEA
4.5. Lamb BW from Birth until Weaning
4.6. Lamb BW and Plasma Cortisol Concentration during Post-Weaning AHP
4.7. Lamb Behavior and Plasma Cortisol Concentrations after Behavior Tests
4.8. Inflammatory and Lymphocyte Proliferation Challenge (IC) Post-AHP
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulbert, L.E.; Moisá, S.J. Stress, immunity, and the management of calves. J. Dairy Sci. 2016, 99, 3199–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, K.M.; Barker, J.P.B. Fetal programming and adult health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Khanal, P.; Nielsen, M.O. Impacts of prenatal nutrition on animal production and performance: A focus on growth and metabolic and endocrine function in sheep. J. Anim. Sci. Biotechnol. 2017, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acuti, G.; Trabalza-Marinucci, M.; Cagiola, M.; Pela, M.; Curina, G.; Moscati, L. Extruded linseed supplementation in the diet of dairy sheep: The effects on immune response and oxidative stress as affected by the physiological state. Small Rumin. Res. 2012, 106S, S21–S28. [Google Scholar] [CrossRef]
- Caroprese, M.; Giannenas, I.; Fthenakis, G.C. Interactions between nutritional approaches and defences against microbial diseases in small ruminants. Vet. Microbiol. 2015, 181, 8–14. [Google Scholar] [CrossRef]
- King, G.J. Comparative placentation in ungulates. J. Exp. Zool. 1993, 266, 588–602. [Google Scholar] [CrossRef]
- Campbell, S.G.; Siegel, M.J.; Knowlton, B.J. Sheep immunoglobulins and their transmission to the neonatal lamb. N. Z. Vet. J. 1977, 25, 361–365. [Google Scholar] [CrossRef]
- Castro, N.; Capote, J.; Bruckmaier, R.M.; Argüello, A. Management effects on colostrogenesis in small ruminants: A review. J. Appl. Anim. Res. 2011, 39, 85–93. [Google Scholar] [CrossRef]
- Hernández-Castellano, L.E.; Suárez-Trujillo, A.; Martell-Jaizme, D.; Cugno, G.; Argüello, A.; Castro, N. The effect of colostrum period management on BW and immune system in lambs: From birth to weaning. Animal 2015, 10, 1672–1679. [Google Scholar] [CrossRef] [Green Version]
- Wathes, D.C.; Abayasekara, D.R.E.; Aitken, R.J. Polyunsaturated fatty acids in male and female reproduction. Biol. Rep. 2007, 77, 190–201. [Google Scholar] [CrossRef]
- Garcia, M.; Greco, L.F.; Favoreto, M.G.; Marsola, R.S.; Wang, D.; Shin, J.H.; Block, E.; Thatcher, W.W.; Santos, J.E.P.; Staples, C.R. Effect of supplementing essential fatty acids to pregnant nonlactating Holstein cows and their preweaned calves on calf performance, immune response, and health. J. Dairy Sci. 2014, 97, 5045–5064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annett, R.W.; Carson, A.F.; Dawson, L.E.R. Effects of digestible undegradable protein (DUP) supply and fish oil supplementation of ewes during late pregnancy on colostrum production and lamb output. Anim. Feed Sci. Technol. 2008, 146, 270–288. [Google Scholar] [CrossRef]
- Macías-Cruz, U.; Mejía-Vázquez, A.; Vicente-Pérez, R.; Correa-Calderón, A.; Robinson, P.H.; Mellado, M.; Meza-Herrera, C.A.; Guerra-Liera, J.E.; Avendaño-Reyes, L. Effects of soybean oil inclusion in the pre-lambing diet on udder size, colostrum secretion, and offspring thermoregulation and growth in hairbreed ewes. Livest. Sci. 2017, 204, 7–15. [Google Scholar] [CrossRef]
- Gulliver, C.E.; Friend, M.A.; King, B.J.; Clayton, E.H. The role of omega-3 polyunsaturated fatty acids in reproduction of sheep and cattle. Anim. Reprod. Sci. 2012, 131, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Capper, J.L.; Wilkinson, R.G.; Mackenzie, A.M.; Sinclair, L.A. Polyunsaturated fatty acid supplementation during pregnancy alters neonatal behavior in sheep. J. Nutr. 2006, 136, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Coleman, D.N.; Murphy, K.D.; Relling, A.R. Prepartum fatty acid supplementation in sheep. II. Supplementation of eicosapentaenoic acid and docosahexaenoic acid during late gestation alters the fatty acid profile of plasma, colostrum, milk and adipose tissue, and increases lipogenic gene expression of adipose tissue. J. Anim. Sci. 2018, 96, 1181–1204. [Google Scholar]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Surlis, C.; Earley, B.; McGee, M.; Keogh, K.; Cormican, P.; Blackshields, G.; Tiernan, K.; Dunn, A.; Morrison, S.; Arguello, A.; et al. Blood immune transciptome analysis if artificially fed dairy calves and naturally suckled beef calves from birth to 7 days of age. Sci. Rep. 2018, 8, 15461. [Google Scholar] [CrossRef]
- Duerkop, B.A.; Vaishnava, S.; Hooper, L.V. Immune responses to the microbiota at the intestinal mucosal surface. Immunity 2009, 31, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Forsythe, P.; Bienenstock, J. Immunomodulation by commensal and probiotic bacteria. Immunol. Investig. 2010, 39, 429–448. [Google Scholar] [CrossRef]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blecha, F. Immune system response to stress. In The Biology of Animal Stress; Moberg, G.P., Mench, J.A., Eds.; CAB International: Wallingford, UK, 2000; pp. 1111–1121. [Google Scholar]
- Radeski, M.; O’Shea, H.; De Meneghi, D.; Ilieski, V. Positioning animal welfare in the One Health concept through evaluation of an Animal Welfare Center in Skopje, Macedonia. Front. Vet. Sci. 2018, 4, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalo, C.; Linage, B.; Carriedo, J.A.; de la Fuente, F.; San Primitivo, F. Evaluation of the overall accuracy of the DeLaval cell counter for somatic cell counts in ovine milk. J. Dairy Sci. 2006, 89, 4613–4619. [Google Scholar] [CrossRef]
- Real Decreto 752/2011. de 27 de Mayo, por el que se Establece la Normativa Básica de Control que Deben Cumplir los Agentes del Sector de Leche Cruda de Oveja y Cabra. Ministerio de la Presidencia, Boletín Oficial del Estado Nº 137, de 9 de junio de 2011 (BOE-A-2011-9995). Available online: https://www.boe.es/buscar/doc.php?id=BOE-A-2011-9995 (accessed on 5 May 2022).
- Kramer, J.K.G.; Blackadar, C.B.; Zhou, J. Evaluation of two GC columns (60-m SUPELCOWAX 10 and 100-m CP Sil 88) for analysis of milkfat with emphasis on CLA, 18:1, 18:2 and 18:3 isomers, and short- and long-chain FA. Lipids 2002, 37, 823–835. [Google Scholar] [CrossRef]
- Precht, D.; Molkentin, J.; Destaillats, F.; Wolff, R. Comparative studies on individual isomeric 18:1 acids in cow, goat, and ewe milk fats by low-temperature high-resolution capillary gas–liquid chromatography. Lipids 2001, 36, 827–832. [Google Scholar] [CrossRef]
- Cruz-Hernandez, C.; Deng, Z.; Zhou, J.; Hill, A.R.; Yurawecz, M.P.; Delmonte, P.; Mossoba, M.M.; Dugan, M.; Kramer, J. Methods to analyze conjugated linoleic acids (CLA) and trans-18:1 isomers in dairy fats using a combination of GC, silver ion TLC-GC, and silver ion HPLC. J. AOAC Int. 2004, 87, 545–560. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.C.; Alves, N.G.; Ascari, I.J.; Junqueira, F.B.; Coutinho, A.S.; Lima, R.R.; Pérez, J.R.O.; De Paula, S.O.; Furusho-Garcia, I.F.; Abreu, L.R. Colostrum composition of Santa Inês sheep and passive transfer of immunity to lambs. J. Dairy Sci. 2015, 98, 3706–3716. [Google Scholar] [CrossRef] [Green Version]
- Quigley, J.D.; Drewry, J.J. Nutrient and immunity transfer from cow to calf pre- and postcalving. J. Dairy Sci. 1998, 81, 2779–2790. [Google Scholar] [CrossRef]
- Destrez, A.; Deiss, V.; Leterrier, C.; Boivin, X.; Boissy, A. Long-term exposure to unpredictable and uncontrollable aversive events alters fearfulness in sheep. Animal 2013, 7, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Averós, X.; Marchewka, J.; Beltrán de Heredia, I.; Zanella, A.J.; Ruiz, R.; Estevez, I. Space allowance during gestation and early maternal separation: Effects on the fear response and social motivation of lambs. Appl. Anim. Behav. Sci. 2015, 163, 98–109. [Google Scholar] [CrossRef]
- Sanchez, C.; Estevez, I. The Chickitizer Software Program; University of Maryland: College Park, ML, USA, 1998. [Google Scholar]
- Averós, X.; Lorea, A.; Beltrán de Heredia, I.; Arranz, J.; Ruiz, R.; Estevez, I. Space availability and movement patterns in dairy sheep during pregnancy. PLoS ONE 2014, 9, e94767. [Google Scholar] [CrossRef] [PubMed]
- Turchi, P. Quantitative Analysis Movement: Measuring and Modelling Population Redistribution in Animals and Plants; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Leone, E.H.; Estevez, I. Space use according to the distribution of resources and level of competition. Poult. Sci. 2008, 87, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Estevez, I.; Mallapur, A.; Miller, C.; Christman, M.C. Short- and long-term movement patterns in complex confined environments in broiler chickens: The effects of distribution of cover panels and food resources. Poult. Sci. 2010, 89, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marini, D.; Pippia, J.; Colditz, I.G.; Hinch, G.; Petherick, J.C.; Lee, C. Randomised trial of the bioavailability and efficacy of orally administered flunixin, carprofen and ketoprofen in a pain model in sheep. Aust. Vet. J. 2015, 93, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Winer, B.J. Statistical Principles in Experimental Design, 2nd ed.; McGraw Hill, Inc.: New York, NY, USA, 1971. [Google Scholar]
- Seal, B.C.; Lillehoj, H.S.; Donovan, D.M.; Gay, C.G. Alternatives to antibiotics: A symposium on the challenges and solutions for animal production. Anim. Health Res. Rev. 2013, 14, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Jolazadeh, A.R.; Mohammadabadi, T.; Dehghan-banadaky, M.; Chaji, M.; Garcia, M. Effect of supplementation fat during the last 3 weeks of uterine life and the preweaning period on performance, ruminal fermentation, blood metabolites, passive immunity and health of the newborn calf. Br. J. Nutr. 2019, 122, 1346–1358. [Google Scholar] [CrossRef]
- Pesántez-Pacheco, J.L.; Heras-Molina, A.; Torres-Rovira, L.; Sanz-Fernández, M.V.; García-Contreras, C.; Vázquez-Gómez, M.; Feyjoo, P.; Cáceres, E.; Frías-Mateo, M.; Hernández, F.; et al. Influence of maternal factors (weight, body condition, parity, and pregnancy rank) on plasma metabolites of dairy ewes and their lambs. Animals 2019, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Musk, G.C.; James, A.; Kemp, M.W.; Ritchie, S.; Ritchie, A.; Laurence, M. Pregnant sheep in a farm environmental do not develop anaemia. Animals 2017, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, N.S.; Panayi, G.S. Inhibition of interleukin-2 production by retroplacental sera; a possible mechanism for human fetal allograft survival. Am. J. Reprod. Immunol. Microbiol. 1985, 9, 6–11. [Google Scholar] [CrossRef]
- Kessler, E.C.; Bruckmaier, R.M.; Gross, J.J. Immunoglobulin G content and colostrum composition of different goat and sheep breeds in Switzerland and Germany. J. Dairy Sci. 2019, 102, 5542–5549. [Google Scholar] [CrossRef]
- Bauman, D.E.; Harvatine, K.J.; Lock, A.L. Nutrigenomics, Rumen-Derived Bioactive Fatty Acids, and the Regulation of Milk Fat Synthesis. Ann. Rev. Nutr. 2011, 3, 299–319. [Google Scholar] [CrossRef] [PubMed]
- Toral, P.G.; Bernard, L.; Belenguer, A.; Rouel, J.; Hervás, G.; Chilliard, Y.; Frutos, P. Comparison of ruminal lipid metabolism in dairy cows and goats fed diets supplemented with starch, plant oil, or fish oil. J. Dairy Sci. 2016, 99, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Banchero, G.E.; Perez, E.; Clariget, R.; Bencina, R.; Lindsay, D.R.; Milton, J.T.B.; Martin, G.B. Endocrine and metabolic factors involved in the effect of nutrition on the production of colostrum in female sheep. Reprod. Nutr. Dev. 2006, 46, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Banchero, G.E.; Quintans, G.; Martin, G.B.; Milton, J.T.B.; Lindsay, D.R. Nutrition and colostrum production in sheep. 2. Metabolic and hormonal responses to different energy sources in the final stages of pregnancy. Reprod. Fertil. Dev. 2004, 16, 645–653. [Google Scholar] [CrossRef]
- Pascual, A.; Pineda-Quiroga, C.; Goiri, I.; Atxaerandio, R.; Ruiz, R.; García-Rodríguez, A. Effects of feeding UFA-rich cold-pressed oilseed cakes and sainfoin on dairy ewes’ milk fatty acid profile and curd sensory properties. Small Rumin. Res. 2019, 175, 96–103. [Google Scholar] [CrossRef]
- Bauman, D.E.; Mather, I.H.; Wall, R.J.; Lock, A.L. Major advances associated with the biosynthesis of milk. J. Dairy Sci. 2006, 89, 1235–1243. [Google Scholar] [CrossRef]
- Dewanckele, L.; Toral, P.G.; Vlaeminck, B.; Fievez, V. Invited review: Role of rumen biohydrogenation intermediates and rumen microbes in diet-induced milk fat depression: An update. J. Dairy Sci. 2020, 103, 7655–7681. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.N.; Kritchevsky, D.; Pariza, M.W. Conjugated linoleic acid and atherosclerosis in rabbits. Atherosclerosis 1994, 108, 19–25. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, S.R.; Ahn, J.Y.; Cho, I.J.; Yoon, C.S.; Ha, T.Y. Dietary conjugated linoleic acid reduces lipid peroxidation by increasing oxidative stability in rats. J. Nutr. Sci. Vitaminol. 2005, 51, 8–15. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, M.; Bassaganya-Riera, J.; Mohede, I.C.M. Immunomodulatory properties of conjugated linoleic acid. Am. J. Clin. Nutr. 2004, 79, 1199S–1206S. [Google Scholar] [CrossRef] [Green Version]
- Castro, N.; Capote, J.; Martín, D.; Argüello, A. The influence of dietary conjugated linoleic acid on blood serum and colostrum immunoglobulin G concentration in female goats before and after parturition. J. Anim. Physiol. Anim. Nutr. 2006, 90, 429–431. [Google Scholar] [CrossRef]
- Hunter, A.G.; Reneau, J.K.; Williams, J.B. Factors affecting IgG concentration in day-old lambs. J. Anim. Sci. 1977, 45, 1146–1151. [Google Scholar] [CrossRef] [PubMed]
- Ciupercescu, D.D. Dynamics of serum immunoglobulin concentrations in sheep during pregnancy and lactation. Res. Vet. Sci. 1977, 22, 23–27. [Google Scholar] [CrossRef]
- Logan, E.F.; Penhale, W.J.; Jones, R.A. Changes in the serum immunoglobulin levels of colostrum-fed calves during the first 12 weeks postpartum. Res. Vet. Sci. 1972, 14, 394–397. [Google Scholar] [CrossRef]
- Renaud, D.L.; Waalderbos, K.M.; Beavers, L.; Duffield, T.F.; Leslie, K.E.; Windeyer, M.C. Risk factors associated with failed transfer of passive immunity in male and female dairy calves: A 2008 retrospective cross-sectional study. J. Dairy Sci. 2020, 103, 3521–3528. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L.; McClure, K.E.; Parker, C.F. Fat fed to pregnant and lactating ewes on milk composition, lamb plasma fatty acids and growth. J. Anim. Sci. 1977, 45, 1152–1159. [Google Scholar] [CrossRef] [Green Version]
- Coleman, D.N.; Rivera-Acevedo, K.C.; Relling, A.R. Prepartum fatty acid supplementation in sheep I. Eicosapentaenoic and docosahexaenoic acid supplementation do not modify ewe and lamb metabolic status and performance through weaning. J. Anim. Sci. 2018, 96, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Destrez, A.; Deiss, V.; Lévy, F.; Calandreau, L.; Lee, C.; Chaillou-Sagon, E.; Boissy, A. Chronic stress induces pessimistic-like judgment and learning deficits in sheep. Appl. Anim. Behav. Sci. 2013, 148, 28–36. [Google Scholar] [CrossRef]
- Braastad, B.O.; Osadchuk, L.V.; Lund, G.; Bakken, M. Effects of prenatal handling stress on adrenal weight and function and behaviour in novel situations in blue fox cubs (Alopex lagopus). Appl. Anim. Behav. Sci. 1998, 57, 157–169. [Google Scholar] [CrossRef]
- Pedernera, M.; Pérez-Sánchez, L.A.; Romero-Aguilar, L.D.; Aguirre, V.; Flores-Pérez, I.; Vázquez, R.; Orihuela, A. Effects of high concentrate supplementation of Saint Croix sheep during peripartum on neonatal lamb behaviour. J. Appl. Anim. Res. 2017, 46, 720–724. [Google Scholar] [CrossRef] [Green Version]
- Coulon, M.; Wellman, C.L.; Marjara, I.S.; Janczak, A.M.; Zanella, A.J. Early adverse experience alters dendritic spine density and gene expression in prefrontal cortex and hippocampus in lambs. Psychoneuroendocrinology 2013, 38, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Coulon, M.; Hild, S.; Schroeer, A.; Janczak, A.M.; Zanella, A.J. Gentle vs. aversive handling of pregnant ewes: II. Physiology and behavior of the lambs. Physiol. Behav. 2011, 103, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Veissier, I.; Boissy, A.; Nowak, R.; Orgeur, P.; Poindron, P. Ontogeny of social awareness in domestic herbivores. Appl. Anim. Behav. Sci. 1998, 57, 233–245. [Google Scholar] [CrossRef]
- Vandenheede, M.; Bouissou, M.F. Sex differences in fear reactions in sheep. Appl. Anim. Behav. Sci. 1993, 37, 39–55. [Google Scholar] [CrossRef]
- Mekaouche, M.; Givalois, L.; Barbanel, G.; Siaud, P.; Maurel, D.; Malaval, F.; Bristow, A.F.; Boissin, J.; Assenmacher, I.; Ixart, G. Chronic restraint enhances interleukin-1-beta release in the basal state and after and endotoxin challenge, independently of adrenocorticotropin and corticosterone release. Neuroimmunomodulation 1994, 1, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Batuman, O.A.; Sajewski, D.; Ottenweller, J.E.; Pitman, D.L.; Natelson, B.H. Effects of repeated stress on T cell numbers and function in rats. Brain Behav. Immun. 1990, 4, 105–117. [Google Scholar] [CrossRef]
- Coppinger, T.R.; Minton, J.E.; Reddy, P.G.; Blecha, F. Repeated restraint and isolation stress in lambs increases pituitary-adrenal secretions and reduces cell-mediated immunity. J. Anim. Sci. 1991, 69, 2808–2814. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.H.; Cantrell, D.A. Signaling and function of interleukin-2 in T lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [Google Scholar] [CrossRef]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Pung, O.J.; Tucker, A.N.; Vore, S.J.; Luster, M.I. Influence of estrogen on host resistance: Increased susceptibility of mice to Listeria monocytogenes correlates with depressed production of interleukin 2. Infect. Immun. 1985, 50, 91–96. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Natal Diet | |||||
|---|---|---|---|---|---|
| CO-FES 1 | ALA-SAIN 2 | ||||
| Variable | Mean | SE | Mean | SE | p-Value |
| Erythrocytes (×106 cells/mm3) | 8.56 | 0.52 | 9.74 | 0.57 | 0.318 |
| Hematocrit (%) 3 | 35.11 | 1.02 | 37.25 | 0.85 | 0.328 |
| Hemoglobin (g/dl) | 10.80 | 0.54 | 11.31 | 0.55 | 0.578 |
| MCV (fl) 3,4 | 39.82 | 1.27 | 38.33 | 1.00 | 0.195 |
| Leucocytes (×103 cells/mm3) 3 | 5.23 | 0.77 | 5.28 | 0.30 | 0.887 |
| Eosinophils (%) | 0.10 | 0.37 | 0.12 | 0.49 | 0.979 |
| Neutrophils (%) | 47.56 | 9.24 | 51.99 | 8.34 | 0.803 |
| Lymphocytes (%) | 46.37 | 9.18 | 41.51 | 8.12 | 0.780 |
| Monocytes (%) | 5.29 | 4.05 | 4.84 | 3.53 | 0.953 |
| N:L 5 | 1.46 | 0.50 | 1.66 | 0.45 | 0.794 |
| Pre-Natal Diet | |||||
|---|---|---|---|---|---|
| CO-FES 1 | ALA-SAIN 2 | ||||
| Variable | Mean | SE | Mean | SE | p-Value |
| Physical measurements | |||||
| Volume (L) 3 | 1410 | 191 | 1501 | 232 | 0.814 |
| Weight (kg) 3 | 1431 | 193 | 1528 | 235 | 0.818 |
| Density (g/L) 3 | 1041 | 1.86 | 1042 | 2.19 | 0.323 |
| pH 3 | 6.47 | 0.10 | 6.40 | 0.08 | 0.527 |
| Chemical composition | |||||
| Fat (%) 3 | 11.99 | 1.24 | 11.82 | 0.97 | 0.085 |
| Total protein (%) | 14.44 | 0.97 | 12.22 | 0.88 | 0.148 |
| Ash (%) 3 | 0.96 | 0.04 | 1.00 | 0.03 | 0.628 |
| Lactose (%) 3 | 3.12 | 0.21 | 3.33 | 0.24 | 0.416 |
| Dry extract (%) | 32.26 | 1.91 | 27.43 | 1.74 | 0.115 |
| Pre-Natal Diet | |||||
|---|---|---|---|---|---|
| CO-FES 1 | ALA-SAIN 2 | ||||
| Variable | Mean | SE | Mean | SE | p-Value |
| Unsaturated fatty acids (UFA; mg/g total FA) | |||||
| c9-14:1 3 | 1.68 | 0.26 | 2.53 | 0.59 | 0.303 |
| t9-16:1 | 1.09 | 0.21 | 1.83 | 0.21 | 0.046 |
| c7-16:1 | 2.15 | 0.19 | 1.94 | 0.20 | 0.493 |
| c9-16:1 | 9.59 | 1.28 | 12.69 | 1.27 | 0.150 |
| ∑ 16:1 3 | 13.54 | 0.61 | 16.10 | 1.15 | 0.042 |
| c9-17:1 | 2.49 | 0.29 | 2.23 | 0.30 | 0.571 |
| ∑ t6- to t9-18:1 3 | 4.61 | 0.20 | 4.88 | 0.33 | 0.849 |
| t10-18:1 | 4.47 | 0.30 | 3.15 | 0.29 | 0.014 |
| t11-18:1 3 | 12.47 | 0.90 | 25.43 | 3.14 | 0.006 |
| t12-18:1 | 2.33 | 0.19 | 2.48 | 0.19 | 0.615 |
| t13- + t14-18:1 4 | 1.10 | 0.22 | 2.50 | 0.22 | 0.002 |
| c9-18:1 3 | 233.2 | 13.02 | 206.1 | 14.47 | 0.062 |
| t15- + c10-18:1 | 1.54 | 0.13 | 1.57 | 0.13 | 0.854 |
| c11-18:1 3 | 3.27 | 0.16 | 3.78 | 0.14 | 0.388 |
| c12-18:1 | 1.45 | 0.11 | 2.03 | 0.13 | 0.099 |
| t16-18:1 3 | 1.18 | 0.07 | 1.78 | 0.15 | 0.010 |
| ∑ C18:1 trans | 26.33 | 3.96 | 39.71 | 3.92 | 0.053 |
| ∑ C18:1 cis 3 | 237.95 | 13.22 | 211.90 | 14.58 | 0.069 |
| ∑trans-MUFA 3 | 27.08 | 1.50 | 42.37 | 4.29 | 0.041 |
| ∑cis-MUFA | 265.7 | 16.64 | 222.3 | 17.33 | 0.114 |
| ∑MUFA 3 | 284.9 | 12.9 | 277.4 | 13.6 | 0.235 |
| c9, c12-18:2 (18:2n-6) | 18.04 | 1.21 | 19.38 | 1.19 | 0.493 |
| c9, c12, c15-18:3 (18:3n-3, ALA) | 4.88 | 0.74 | 9.82 | 0.73 | <0.001 |
| c11, c14, c17-20:3 (20:3n-3) | 2.25 | 0.12 | 2.04 | 0.11 | 0.260 |
| 7c,10c,13c,16c,19c-22:5 (22:5n-3) | 1.74 | 0.10 | 1.91 | 0.10 | 0.268 |
| 4c,7c,10c,13c,16c,19c-22:6 (22:6n-3) | 1.27 | 0.16 | 1.61 | 0.16 | 0.212 |
| ∑PUFA | 39.50 | 2.49 | 52.01 | 2.46 | 0.008 |
| c9, t11-18:2 (c9,t11-CLA or RA) 5 | 8.56 | 0.93 | 13.24 | 0.92 | 0.007 |
| ∑CLA | 9.21 | 1.01 | 14.77 | 1.00 | 0.005 |
| Ratio t11-18:1/t10-18:1 | 3.03 | 0.49 | 7.46 | 0.48 | <0.001 |
| ∑n-3 3 | 10.26 | 0.40 | 16.10 | 0.70 | 0.004 |
| ∑n-6 | 20.10 | 1.26 | 21.78 | 1.25 | 0.411 |
| n-6:n-3 3 | 1.91 | 0.10 | 1.38 | 0.05 | 0.008 |
| Pre-Natal Diet | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CO-FES 1 | ALA-SAIN 2 | ||||||||||
| At Birth | 24 h Post-Lambing | At Birth | 24 h Post-Lambing | ||||||||
| Variable | Mean | SE | Mean | SE | Mean | SE | Mean | SE | p-Value Pre-Natal Diet | p-Value Time | p-Value Pre-Natal Diet × Time |
| SCC 3 (×103 cells/mL) 4 | 410.7 | 153.8 | 198.7 | 55.7 | 486.7 | 204.1 | 624.5 | 301.9 | 0.548 | 0.587 | 0.385 |
| Total Solids (° Bx) | 23.90 | 1.25 | 18.10 | 1.50 | 22.03 | 1.18 | 14.95 | 1.18 | 0.179 | <0.001 | 0.513 |
| IgG (mg/mL) | 47.48 | 3.43 | 9.65 | 3.43 | 40.78 | 3.41 | 6.07 | 3.41 | 0.214 | <0.001 | 0.627 |
| Pre-Natal Diet | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CO-FES 1 | ALA-SAIN 2 | ||||||||||
| Female | Male | Female | Male | ||||||||
| Variable | Mean | SE | Mean | SE | Mean | SE | Mean | SE | p-Value Pre-Natal Diet | p-Value Sex | p-Value Pre-Natal Diet × Sex |
| Social isolation test | |||||||||||
| Passive stand (%) 4 | 58.5 | 9.3 | 48.3 | 9.7 | 46.5 | 9.1 | 41.9 | 9.5 | 0.058 | 0.129 | 0.612 |
| Move (%) 4 | 17.9 | 5.0 | 16.0 | 4.7 | 18.0 | 4.9 | 18.2 | 5.4 | 0.707 | 0.790 | 0.736 |
| Explore (%) 4 | 14.8 | 4.2 | 16.6 | 4.6 | 26.3 | 5.1 | 16.7 | 4.6 | 0.156 | 0.385 | 0.238 |
| Escape attempts (%) 4 | 7.6 | 4.0 | 17.8 | 7.7 | 7.5 | 3.7 | 21.0 | 0.9 | 0.738 | 0.002 | 0.719 |
| Total distance (m) | 2.9 | 0.5 | 3.0 | 0.5 | 2.6 | 0.4 | 2.7 | 0.5 | 0.529 | 0.741 | 0.974 |
| Angular dispersion 3 | 0.27 | 0.02 | 0.33 | 0.03 | 0.31 | 0.04 | 0.42 | 0.06 | 0.170 | 0.034 | 0.693 |
| Excretions (n) 5 | 0.0 | 0.0 | 0.2 | 0.2 | 0.2 | 0.2 | 0.3 | 0.3 | 0.977 | 0.977 | 0.978 |
| Vocalizations (n) 5 | 63.3 | 6.0 | 74.3 | 7.1 | 73.6 | 6.7 | 77.9 | 7.7 | 0.096 | 0.064 | 0.398 |
| Social motivation test | |||||||||||
| Passive stand (%) 4 | 50.4 | 9.7 | 51.8 | 9.9 | 47.2 | 9.3 | 36.6 | 9.1 | 0.058 | 0.325 | 0.276 |
| Move (%) 4 | 11.3 | 3.1 | 11.9 | 3.3 | 11.6 | 2.8 | 14.4 | 3.9 | 0.627 | 0.576 | 0.746 |
| Escape attempts (%) 4 | 1.4 | 2.2 | 1.3 | 2.1 | 0.2 | 0.4 | 2.0 | 3.1 | 0.288 | 0.186 | 0.223 |
| Interact with lambs (%) 4 | 27.1 | 8.9 | 21.6 | 7.8 | 41.9 | 10.5 | 37.9 | 10.5 | 0.002 | 0.266 | 0.773 |
| Latency to the fence (s) 3 | 2.2 | 0.4 | 2.4 | 0.5 | 4.2 | 1.2 | 1.5 | 0.3 | 0.896 | 0.1337 | 0.090 |
| Time close to lambs (%) 4 | 92.1 b | 5.2 | 85.3 c | 10.2 | 98.4 a | 1.3 | 89.6 bc | 7.6 | <0.001 | <0.001 | <0.001 |
| Total distance (m) | 2.0 | 0.3 | 1.7 | 0.3 | 1.7 | 0.3 | 2.4 | 0.3 | 0.575 | 0.451 | 0.105 |
| Angular dispersion 3 | 0.21 | 0.01 | 0.26 | 0.04 | 0.33 | 0.03 | 0.31 | 0.07 | 0.051 | 0.839 | 0.251 |
| Excretions (n) 5 | 0.8 a | 0.3 | 0.1 ab | 0.3 | 0.0 b | 0.3 | 0.3 ab | 0.3 | 0.168 | 0.228 | 0.022 |
| Vocalizations (n) 5 | 14.8 | 3.1 | 11.6 | 2.7 | 14.2 | 3.0 | 11.0 | 2.6 | 0.712 | 0.079 | 0.968 |
| Pre-Natal Diet | Sex | Hours Post-IC 3 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CO-FES 1 | ALA-SAIN 2 | Male | Female | 24 h | 48 h | 72 h | ||||||||||||||
| Variable | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | p-Value Pre-Natal Diet | p-Value Sex | p-Value Time | p-Value Pre-Natal Diet × Sex | p-Value Pre-Natal Diet × Time | p-Value Sex × Time |
| Skinfold thickness (mm) 4 | 8.6 | 0.3 | 8.7 | 0.3 | 9.0 | 0.3 | 8.4 | 0.3 | 9.7 a | 0.4 | 8.9 a | 0.3 | 7.5 b | 0.2 | 0.523 | 0.481 | <0.001 | 0.923 | 0.381 | 0.626 |
| Skin temperature (°C) 4 | 32.0 | 0.3 | 32.7 | 0.2 | 32.0 | 0.2 | 32.7 | 0.3 | 32.6 a | 0.3 | 32.4 a | 0.3 | 32.0 b | 0.3 | 0.823 | 0.452 | 0.038 | 0.171 | 0.792 | 0.994 |
| Pre-Natal Diet | Sex | Hours Post-IC 3 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CO-FES 1 | ALA-SAIN 2 | Male | Female | 0 h | 24 h | 48 h | ||||||||||||||
| Variable | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | p-Value Pre-Natal Diet | p-Value Sex | p-Value Time | P-Value Pre-Natal Diet × Sex | p-Value Pre-Natal Diet × Time | p-Value Sex × Time |
| Cortisol (ng/mL) 4 | 83.2 | 10.1 | 74.9 | 8.0 | 84.5 | 10.4 | 74.7 | 8.0 | 98.6 | 11.4 | 72.8 | 10.5 | 65.8 | 10.5 | 0.047 | 0.548 | 0.051 | 0.638 | 0.837 | 0.766 |
| IL-2 (pg/mL) 4 | 51.4 | 0.2 | 53.8 | 0.9 | 53.2 | 1.0 | 52.1 | 0.4 | 52.0 | 0.6 | 53.0 | 1.3 | 52.7 | 0.6 | 0.003 | 0.084 | 0.600 | 0.050 | 0.306 | 0.078 |
| IL-10 (pg/mlL) 4 | 14.9 | 0.5 | 14.3 | 0.4 | 14.1 | 0.6 | 15.0 | 0.4 | 14.2 | 0.5 | 14.3 | 0.8 | 15.3 | 0.6 | 0.318 | 0.112 | 0.300 | 0.718 | 0.435 | 0.454 |
| IL-1β (pg/mL) 4 | 40.1 | 2.8 | 43.2 | 3.9 | 41.8 | 3.0 | 41.5 | 3.7 | 47.9 | 6.2 | 39.1 | 1.3 | 38.0 | 3.3 | 0.540 | 0.682 | 0.269 | 0.501 | 0.789 | 0.309 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Averós, X.; Granado-Tajada, I.; Arranz, J.; Beltrán de Heredia, I.; González, L.; Ruiz, R.; García-Rodríguez, A.; Atxaerandio, R. Pre-Partum Supplementation with Polyunsaturated Fatty Acids on Colostrum Characteristics and Lamb Immunity and Behavior after a Mild Post-Weaning Aversive Handling Period. Animals 2022, 12, 1780. https://doi.org/10.3390/ani12141780
Averós X, Granado-Tajada I, Arranz J, Beltrán de Heredia I, González L, Ruiz R, García-Rodríguez A, Atxaerandio R. Pre-Partum Supplementation with Polyunsaturated Fatty Acids on Colostrum Characteristics and Lamb Immunity and Behavior after a Mild Post-Weaning Aversive Handling Period. Animals. 2022; 12(14):1780. https://doi.org/10.3390/ani12141780
Chicago/Turabian StyleAverós, Xavier, Itsasne Granado-Tajada, Josune Arranz, Ignacia Beltrán de Heredia, Laura González, Roberto Ruiz, Aser García-Rodríguez, and Raquel Atxaerandio. 2022. "Pre-Partum Supplementation with Polyunsaturated Fatty Acids on Colostrum Characteristics and Lamb Immunity and Behavior after a Mild Post-Weaning Aversive Handling Period" Animals 12, no. 14: 1780. https://doi.org/10.3390/ani12141780