
Mito-TEMPO Improves the Meiosis Resumption and Mitochondrial Function of Vitrified Sheep Oocytes via the Recovery of Respiratory Chain Activity
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Collection of COCs
2.3. Vitrification and COC Warming
2.4. In Vitro Maturation
2.5. Oocyte Survival and the First Polar Body Extrusion (PBE)
2.6. Immunofluorescence (IF) Staining
2.7. Intracellular ROS and GSH Level Assay
2.8. Quantification of Mitochondrial Function Staining
2.9. Quantification of Mitochondrial Membrane Potentials (∆Ψm)
2.10. ATP Content Determination
2.11. Quantification of Cytoplasmic Ca2+ ([Ca2+]c), Mitochondrial Ca2+ ([Ca2+]m), and Endoplasmic Reticulum Ca2+ ([Ca2+]ER)
2.12. Building a Single-Cell RNA Library and Sequencing the Transcriptome
2.13. RNA Isolation and Real-Time Quantitative PCR (RT-qPCR)
2.14. Statistical Analysis
3. Results
3.1. Mito-TEMPO Improved the Oocyte Viability and Meiotic Maturation after Vitrification
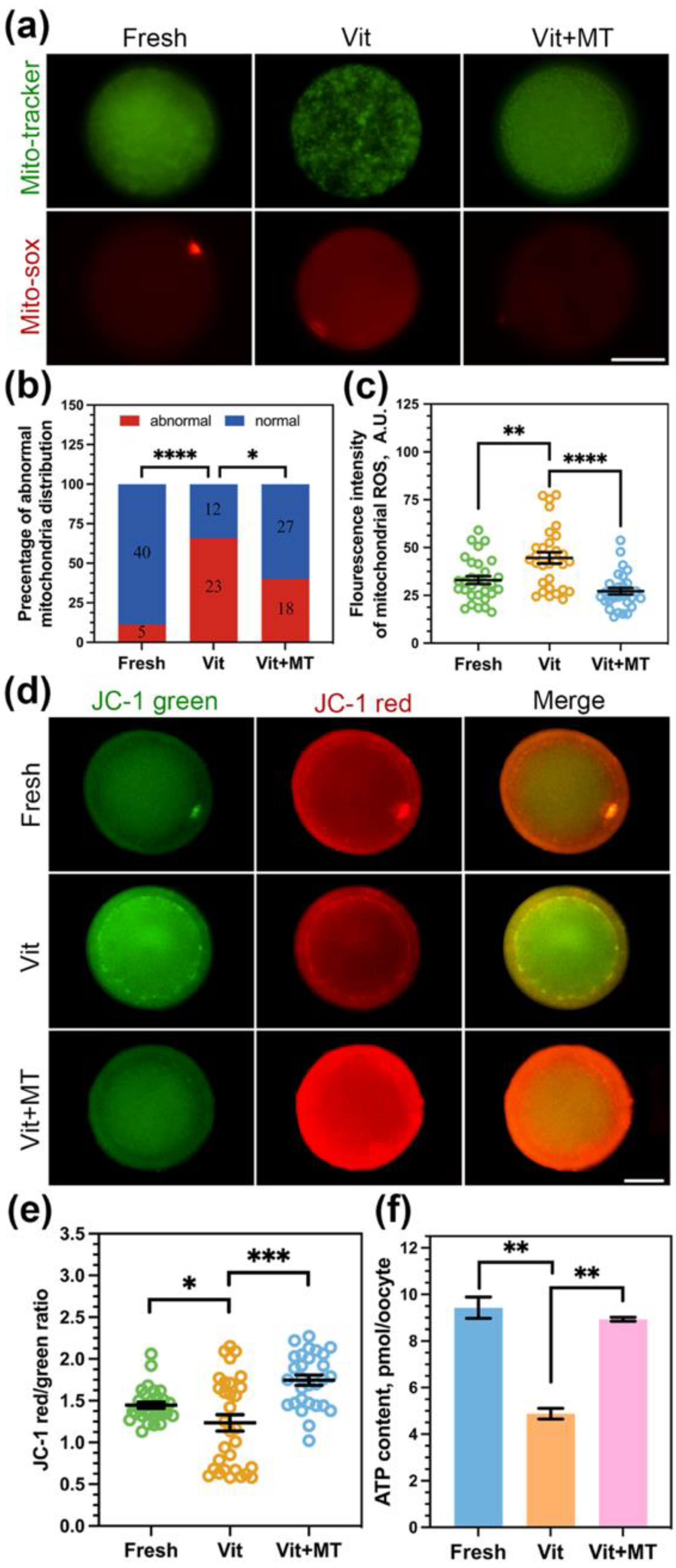
3.2. Mito-TEMPO Inhibits Oxidative Stress after Vitrification
3.3. Mito-TEMPO Restores Mitochondrial Function in Vitrified Oocytes
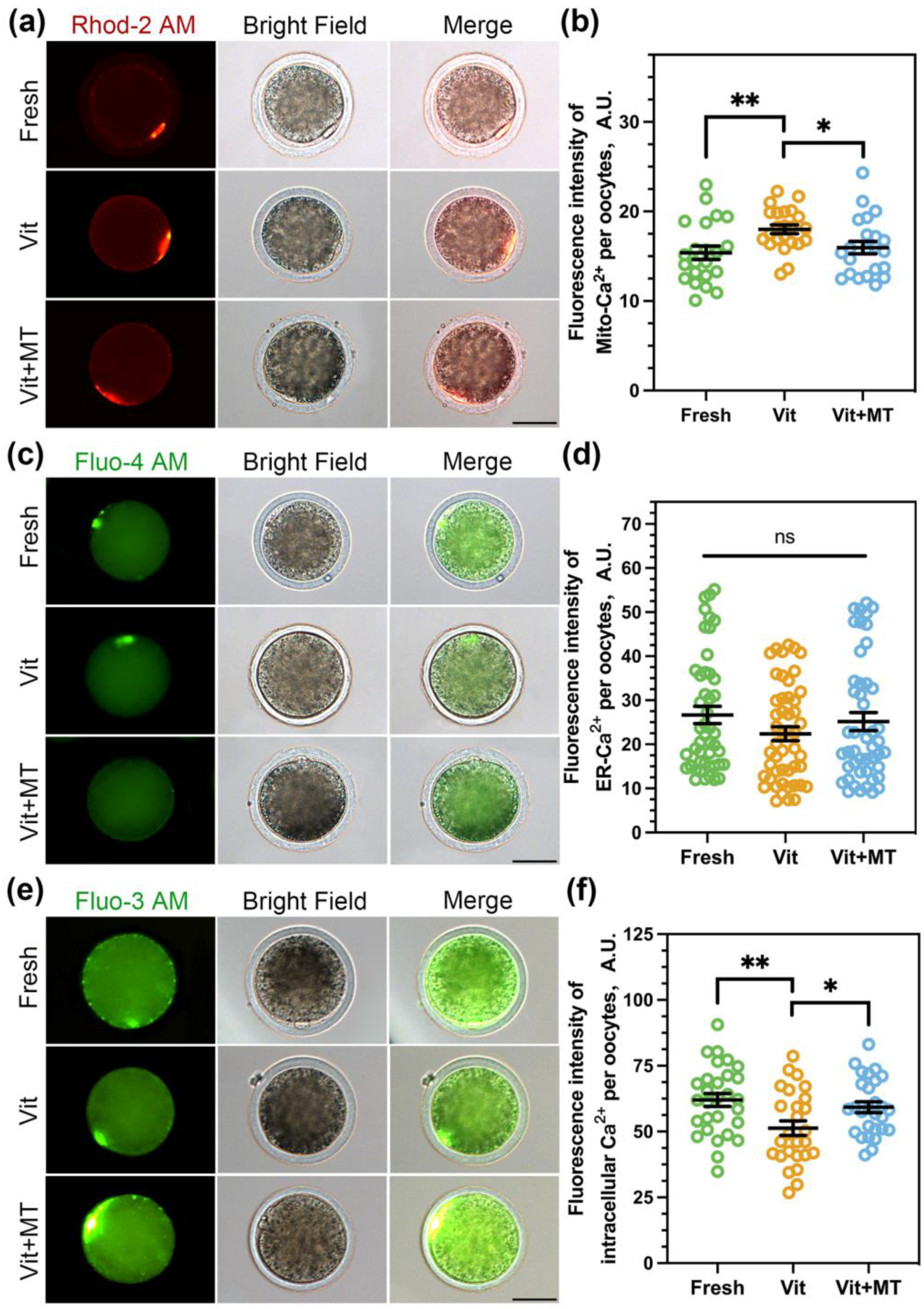
3.4. Mito-TEMPO Restores Mitochondrial and Cytoplasmic Ca2+ Levels in Vitrified Oocytes
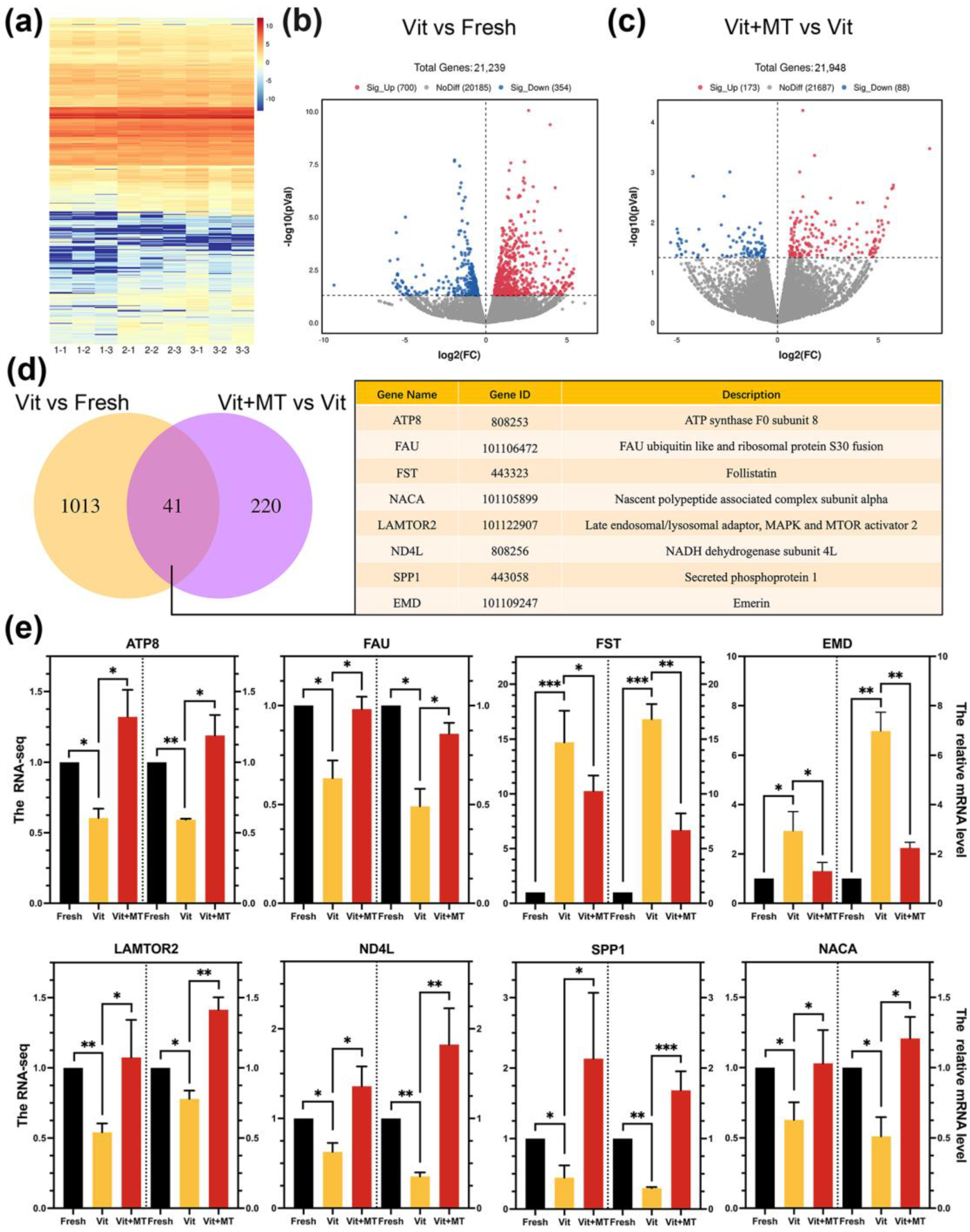
3.5. Investigating the Target Effectors of Mito-TEMPO in Vitrified Oocytes Using Single-Cell Transcriptome Analysis
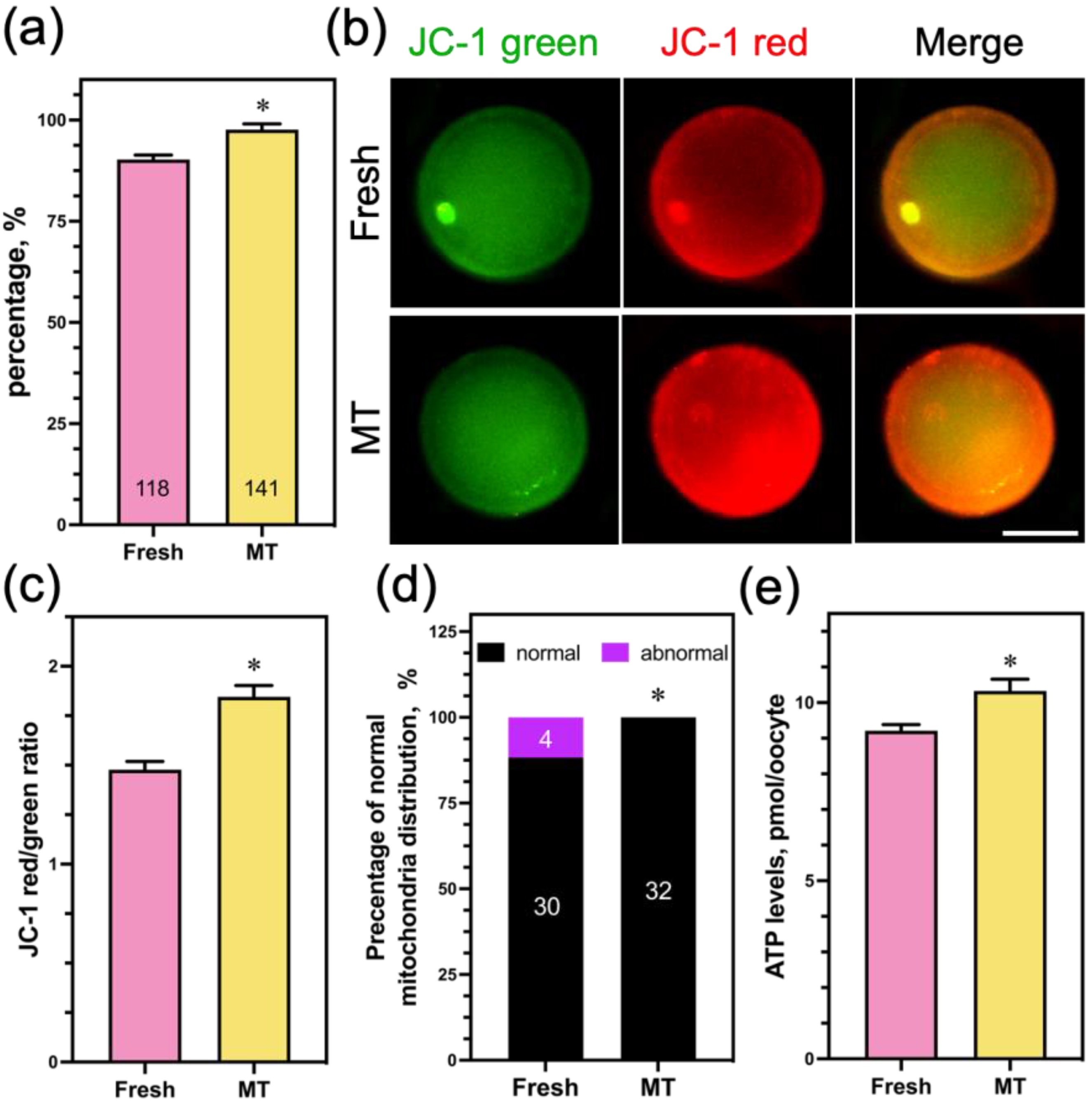
3.6. Mito-TEMPO at a Concentration of 1 μM Had No Potential Toxic Effect on Oocyte Mitochondrial Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quan, G.; Lv, C.; Liang, J.; Zhao, X.; Wu, G. The Assessment of Various Factors Affecting the Postwarming Viability and Developmental Capability of Goat Metaphase II Oocytes Vitrified by Cryotop. Biopreserv. Biobank. 2022, 20, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Galeati, G.; Spinaci, M.; Vallorani, C.; Bucci, D.; Porcu, E.; Tamanini, C. Pig oocyte vitrification by cryotop method: Effects on viability, spindle and chromosome configuration and in vitro fertilization. Anim. Reprod. Sci. 2011, 127, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Guo, N.; Tan, M.H.; Li, Y.F. Effect of mouse oocyte vitrification on mitochondrial membrane potential and distribution. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Bogliolo, L.; Murrone, O.; Piccinini, M.; Ariu, F.; Ledda, S.; Tilocca, S.; Albertini, D.F. Evaluation of the impact of vitrification on the actin cytoskeleton of in vitro matured ovine oocytes by means of Raman microspectroscopy. J. Assist. Reprod. Genet. 2015, 32, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Konc, J.; Kanyo, K.; Kriston, R.; Somoskoi, B.; Cseh, S. Cryopreservation of embryos and oocytes in human assisted reproduction. Biomed. Res. Int. 2014, 2014, 307268. [Google Scholar] [CrossRef]
- Coello, A.; Campos, P.; Remohi, J.; Meseguer, M.; Cobo, A. A combination of hydroxypropyl cellulose and trehalose as supplementation for vitrification of human oocytes: A retrospective cohort study. J. Assist. Reprod. Genet. 2016, 33, 413–421. [Google Scholar] [CrossRef]
- Choi, J.K.; Huang, H.; He, X. Improved low-CPA vitrification of mouse oocytes using quartz microcapillary. Cryobiology 2015, 70, 269–272. [Google Scholar] [CrossRef]
- Ekpo, M.D.; Xie, J.; Hu, Y.; Liu, X.; Liu, F.; Xiang, J.; Zhao, R.; Wang, B.; Tan, S. Antifreeze Proteins: Novel Applications and Navigation towards Their Clinical Application in Cryobanking. Int. J. Mol. Sci. 2022, 23, 2639. [Google Scholar] [CrossRef]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef]
- Martinez-Diez, M.; Santamaria, G.; Ortega, A.D.; Cuezva, J.M. Biogenesis and dynamics of mitochondria during the cell cycle: Significance of 3’UTRs. PLoS ONE 2006, 1, e107. [Google Scholar] [CrossRef]
- Soares, M.; Sousa, A.P.; Fernandes, R.; Ferreira, A.F.; Almeida-Santos, T.; Ramalho-Santos, J. Aging-related mitochondrial alterations in bovine oocytes. Theriogenology 2020, 157, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Sendra, L.; Garcia-Mares, A.; Herrero, M.J.; Alino, S.F. Mitochondrial DNA Replacement Techniques to Prevent Human Mitochondrial Diseases. Int. J. Mol. Sci. 2021, 22, 551. [Google Scholar] [CrossRef] [PubMed]
- Fields, M.; Marcuzzi, A.; Gonelli, A.; Celeghini, C.; Maximova, N.; Rimondi, E. Mitochondria-Targeted Antioxidants, an Innovative Class of Antioxidant Compounds for Neurodegenerative Diseases: Perspectives and Limitations. Int. J. Mol. Sci. 2023, 24, 3739. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Wipf, P.; Stoyanovsky, D.; Greenberger, J.S.; Borisenko, G.; Belikova, N.A.; Yanamala, N.; Samhan Arias, A.K.; Tungekar, M.A.; Jiang, J.; et al. Mitochondrial targeting of electron scavenging antioxidants: Regulation of selective oxidation vs random chain reactions. Adv. Drug Deliv. Rev. 2009, 61, 1375–1385. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, Y.; Li, L.; Liu, S.; Wang, C.; Yuan, Y.; Yang, G.; Chen, Y.; Cheng, J.; Lu, Y.; et al. Mitochondrial ROS promote mitochondrial dysfunction and inflammation in ischemic acute kidney injury by disrupting TFAM-mediated mtDNA maintenance. Theranostics 2021, 11, 1845–1863. [Google Scholar] [CrossRef] [PubMed]
- Yousefian, I.; Zare-Shahneh, A.; Kohram, H. Addition of MitoTEMPO to the maturation medium improves in vitro maturation competence of bovine oocytes. Vet. Res. Forum 2022, 13, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, S.Y.; Kim, J.W.; Yang, S.G.; Kim, M.J.; Jegal, H.G.; Kim, I.S.; Choo, Y.K.; Koo, D.B. Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro. Int. J. Mol. Sci. 2018, 19, 3422. [Google Scholar] [CrossRef] [PubMed]
- Zarei, F.; Daghigh-Kia, H.; Masoudi, R. Supplementation of ram’s semen extender with Mito-TEMPO II: Quality evaluation and flow cytometry study of post-thawed spermatozoa. Andrologia 2022, 54, e14299. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, Y.; Bai, H.; Liu, J.; Li, J.; Wu, B. Mitochondria-targeted antioxidant MitoTEMPO improves the post-thaw sperm quality. Cryobiology 2018, 80, 26–29. [Google Scholar] [CrossRef]
- Masoudi, R.; Asadzadeh, N.; Sharafi, M. The mitochondria-targeted antioxidant Mito-TEMPO conserves rooster’s cooled semen quality and fertility potential. Theriogenology 2020, 156, 236–241. [Google Scholar] [CrossRef]
- Zarei, F.; Kia, H.D.; Masoudi, R.; Moghaddam, G.; Ebrahimi, M. Supplementation of ram’s semen extender with Mito-TEMPO I: Improvement in quality parameters and reproductive performance of cooled-stored semen. Cryobiology 2021, 98, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Leoni, G.G.; Bebbere, D.; Berlinguer, F.; Mossa, F.; Bogliolo, L.; Madeddu, M.; Ledda, S.; Naitana, S. Vitrification devices affect structural and molecular status of in vitro matured ovine oocytes. Mol. Reprod. Dev. 2007, 74, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhang, L.; Qi, Q.; Li, J.; Yan, F.; Hou, J. Growth hormone treatment improves the development of follicles and oocytes in prepubertal lambs. J. Ovarian Res. 2023, 16, 132. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, M.; Machado, S.A.; Stojkovic, P.; Zakhartchenko, V.; Hutzler, P.; Goncalves, P.B.; Wolf, E. Mitochondrial distribution and adenosine triphosphate content of bovine oocytes before and after in vitro maturation: Correlation with morphological criteria and developmental capacity after in vitro fertilization and culture. Biol. Reprod. 2001, 64, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Pan, B.; Qazi, I.H.; Yang, H.; Guo, S.; Yang, J.; Zhang, Y.; Zeng, C.; Zhang, M.; Han, H.; et al. Melatonin Improves In Vitro Development of Vitrified-Warmed Mouse Germinal Vesicle Oocytes Potentially via Modulation of Spindle Assembly Checkpoint-Related Genes. Cells 2019, 8, 1009. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Fan, L.H.; Li, A.; Dong, F.; Hou, Y.; Schatten, H.; Sun, Q.Y.; Ou, X.H. Effects of various calcium transporters on mitochondrial Ca2+ changes and oocyte maturation. J. Cell. Physiol. 2021, 236, 6548–6558. [Google Scholar] [CrossRef] [PubMed]
- Zhuan, Q.; Li, J.; Du, X.; Zhang, L.; Meng, L.; Luo, Y.; Zhou, D.; Liu, H.; Wan, P.; Hou, Y.; et al. Antioxidant procyanidin B2 protects oocytes against cryoinjuries via mitochondria regulated cortical tension. J. Anim. Sci. Biotechnol. 2022, 13, 95. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Du, M.; Zhuan, Q.; Luo, Y.; Li, J.; Hou, Y.; Zeng, S.; Zhu, S.; Fu, X. Melatonin rescues the aneuploidy in mice vitrified oocytes by regulating mitochondrial heat product. Cryobiology 2019, 89, 68–75. [Google Scholar] [CrossRef]
- Ahmadi, E.; Shirazi, A.; Shams-Esfandabadi, N.; Nazari, H. Antioxidants and glycine can improve the developmental competence of vitrified/warmed ovine immature oocytes. Reprod. Domest. Anim. 2019, 54, 595–603. [Google Scholar] [CrossRef]
- Mo, X.H.; Fu, X.W.; Yuan, D.S.; Wu, G.Q.; Jia, B.Y.; Cheng, K.R.; Du, M.; Zhou, Y.H.; Yue, M.X.; Hou, Y.P.; et al. Effect of meiotic status, cumulus cells and cytoskeleton stabilizer on the developmental competence of ovine oocytes following vitrification. Small Rumin. Res. 2014, 117, 151–157. [Google Scholar] [CrossRef]
- Mullen, S.F.; Fahy, G.M. A chronologic review of mature oocyte vitrification research in cattle, pigs, and sheep. Theriogenology 2012, 78, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Al-Zubaidi, U.; Adhikari, D.; Cinar, O.; Zhang, Q.H.; Yuen, W.S.; Murphy, M.P.; Rombauts, L.; Robker, R.L.; Carroll, J. Mitochondria-targeted therapeutics, MitoQ and BGP-15, reverse aging-associated meiotic spindle defects in mouse and human oocytes. Hum. Reprod. 2021, 36, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Gadau, S.D.; Serra, E.; Zinellu, A.; Carru, C.; Porcu, C.; Naitana, S.; Berlinguer, F.; Leoni, G.G. A recovery time after warming restores mitochondrial function and improves developmental competence of vitrified ovine oocytes. Theriogenology 2018, 110, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Castillo, E.; Diaz, F.A.; Talbot, S.A.; Bondioli, K.R. Effect of bovine oocyte vitrification with EGTA and post-warming recovery with resveratrol on meiotic spindle, mitochondrial function, reactive oxygen species, and developmental competence. Theriogenology 2023, 196, 59–67. [Google Scholar] [CrossRef]
- Xu, X.; Yang, B.; Zhang, H.; Feng, X.; Hao, H.; Du, W.; Zhu, H.; Khan, A.; Khan, M.Z.; Zhang, P.; et al. Effects of beta-Nicotinamide Mononucleotide, Berberine, and Cordycepin on Lipid Droplet Content and Developmental Ability of Vitrified Bovine Oocytes. Antioxidants 2023, 12, 991. [Google Scholar] [CrossRef]
- Dalton, C.M.; Carroll, J. Biased inheritance of mitochondria during asymmetric cell division in the mouse oocyte. J. Cell. Sci. 2013, 126, 2955–2964. [Google Scholar] [CrossRef]
- Zhou, D.; Zhuan, Q.; Luo, Y.; Liu, H.; Meng, L.; Du, X.; Wu, G.; Hou, Y.; Li, J.; Fu, X. Mito-Q promotes porcine oocytes maturation by maintaining mitochondrial thermogenesis via UCP2 downregulation. Theriogenology 2022, 187, 205–214. [Google Scholar] [CrossRef]
- Walkon, L.L.; Strubbe-Rivera, J.O.; Bazil, J.N. Calcium Overload and Mitochondrial Metabolism. Biomolecules 2022, 12, 1891. [Google Scholar] [CrossRef]
- Glancy, B.; Balaban, R.S. Role of mitochondrial Ca2+ in the regulation of cellular energetics. Biochemistry 2012, 51, 2959–2973. [Google Scholar] [CrossRef]
- Peng, T.I.; Jou, M.J. Oxidative stress caused by mitochondrial calcium overload. Ann. N. Y. Acad. Sci. 2010, 1201, 183–188. [Google Scholar] [CrossRef]
- van der Reest, J.; Nardini Cecchino, G.; Haigis, M.C.; Kordowitzki, P. Mitochondria: Their relevance during oocyte ageing. Ageing Res. Rev. 2021, 70, 101378. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Guo, Y.; Yu, R.; Wang, J.; Liu, Y.; Chen, H.; Pang, W.; Yang, G.; Chu, G.; Gao, L. Ru360 protects against vitrification-induced oocyte meiotic defects by restoring mitochondrial function. Theriogenology 2023, 204, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.; Fearnley, I.M.; Skehel, J.M.; Shannon, R.J.; Hirst, J.; Walker, J.E. Bovine complex I is a complex of 45 different subunits. J. Biol. Chem. 2006, 281, 32724–32727. [Google Scholar] [CrossRef] [PubMed]
- Mastrorocco, A.; Ciani, E.; Nicassio, L.; Roelen, B.A.J.; Minervini, F.; Dell’Aquila, M.E. Beauvericin alters the expression of genes coding for key proteins of the mitochondrial chain in ovine cumulus-oocyte complexes. Mycotoxin Res. 2021, 37, 1–9. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, D.; Ju, S.; Sun, L.; Zhang, S.; Wu, C.; Rui, R.; Dai, J. Mitophagy is involved in the mitochondrial dysfunction of vitrified porcine oocytes. Mol. Reprod. Dev. 2021, 88, 427–436. [Google Scholar] [CrossRef]
- Yang, S.G.; Bae, J.W.; Park, H.J.; Koo, D.B. Mito-TEMPO protects preimplantation porcine embryos against mitochondrial fission-driven apoptosis through DRP1/PINK1-mediated mitophagy. Life Sci. 2023, 315, 121333. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | NCBI Reference Sequences | Primer Sequence (5′ to 3′) | Annealing Temperature (°C) |
|---|---|---|---|
| GADPH | NM_001190390 | F: GGTTGTCTCCTGCGACTTCA R: CAGGGCCTTGAGGATGGAAA | 55 |
| ATP8 | XR_003587932.1 | F: GCCACAACTAGACACATCAACG R: AGGGGTAATGAAAGAGGCAAA | 55 |
| FAU | XM_004019665.4 | F: TAGCTTCGTTGGAGGGCATC R: TTAGGGCCCTTCTTCTTGCC | 55 |
| FST | XM_012096672.3 | F: TATGGGACTTCAAGGTTGGC R: GGTGTCTTCCGAAATGGAGT | 50 |
| NACA | XM_012174719.3 | F: CAACAATGCCCTCTGGCAAC R: TTCTGAGCGAGCAACTGGAG | 50 |
| LAMTOR2 | XM_004009674.3 | F: AGACCGTTGGCTTCGGAATG R: TGATGCTGCTACTTGGGTGAG | 55 |
| ND4L | XR_003587932.1 | F: TCACAGTATCCCTCACAGGACT R: CTCGCAAGCTGCGAAAACTA | 55 |
| SPP1 | NM_001009224.1 | F: TCCGCCCTTCCAGTTAAACC R: TCAGGGGTTTCAGCATCGTC | 55 |
| EMD | XM_027963104.1 | F: CGCCAGTACAACATCCCACA R: ACGGACGCCGAATCTAAGTC | 55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Dilixiati, A.; Zhang, L.; Aihemaiti, A.; Song, Y.; Zhao, G.; Fu, X.; Wang, X.; Wusiman, A. Mito-TEMPO Improves the Meiosis Resumption and Mitochondrial Function of Vitrified Sheep Oocytes via the Recovery of Respiratory Chain Activity. Animals 2024, 14, 152. https://doi.org/10.3390/ani14010152
Zhao X, Dilixiati A, Zhang L, Aihemaiti A, Song Y, Zhao G, Fu X, Wang X, Wusiman A. Mito-TEMPO Improves the Meiosis Resumption and Mitochondrial Function of Vitrified Sheep Oocytes via the Recovery of Respiratory Chain Activity. Animals. 2024; 14(1):152. https://doi.org/10.3390/ani14010152
Chicago/Turabian StyleZhao, Xi, Airixiati Dilixiati, Luyao Zhang, Aikebaier Aihemaiti, Yukun Song, Guodong Zhao, Xiangwei Fu, Xuguang Wang, and Abulizi Wusiman. 2024. "Mito-TEMPO Improves the Meiosis Resumption and Mitochondrial Function of Vitrified Sheep Oocytes via the Recovery of Respiratory Chain Activity" Animals 14, no. 1: 152. https://doi.org/10.3390/ani14010152