
C-Type Natriuretic Peptide Pre-Treatment Improves Maturation Rate of Goat Oocytes by Maintaining Transzonal Projections, Spindle Morphology, and Mitochondrial Function
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. IACUC Statement
2.2. Antibodies and Chemicals
2.3. COCs Isolation and In Vitro Maturation
2.4. Immunofluorescence Staining
2.5. Measurement of cGMP and cAMP Levels in Oocytes
2.6. Measurement of Intra-Oocyte Reactive Oxygen Species (ROS) and Glutathione (GSH) Levels
2.7. Oocyte MitoTracker Staining
2.8. Measurement of Mitochondrial Membrane Potential (MMP)
2.9. Real-Time Quantitative PCR Analysis
2.10. Statistical Analysis
2.11. Experimental Design
3. Results
3.1. CNP Treatment Maintained Meiotic Arrest and Improved Oocyte Maturation
3.2. CNP Treatment before IVM Maintained TZPs of COCs
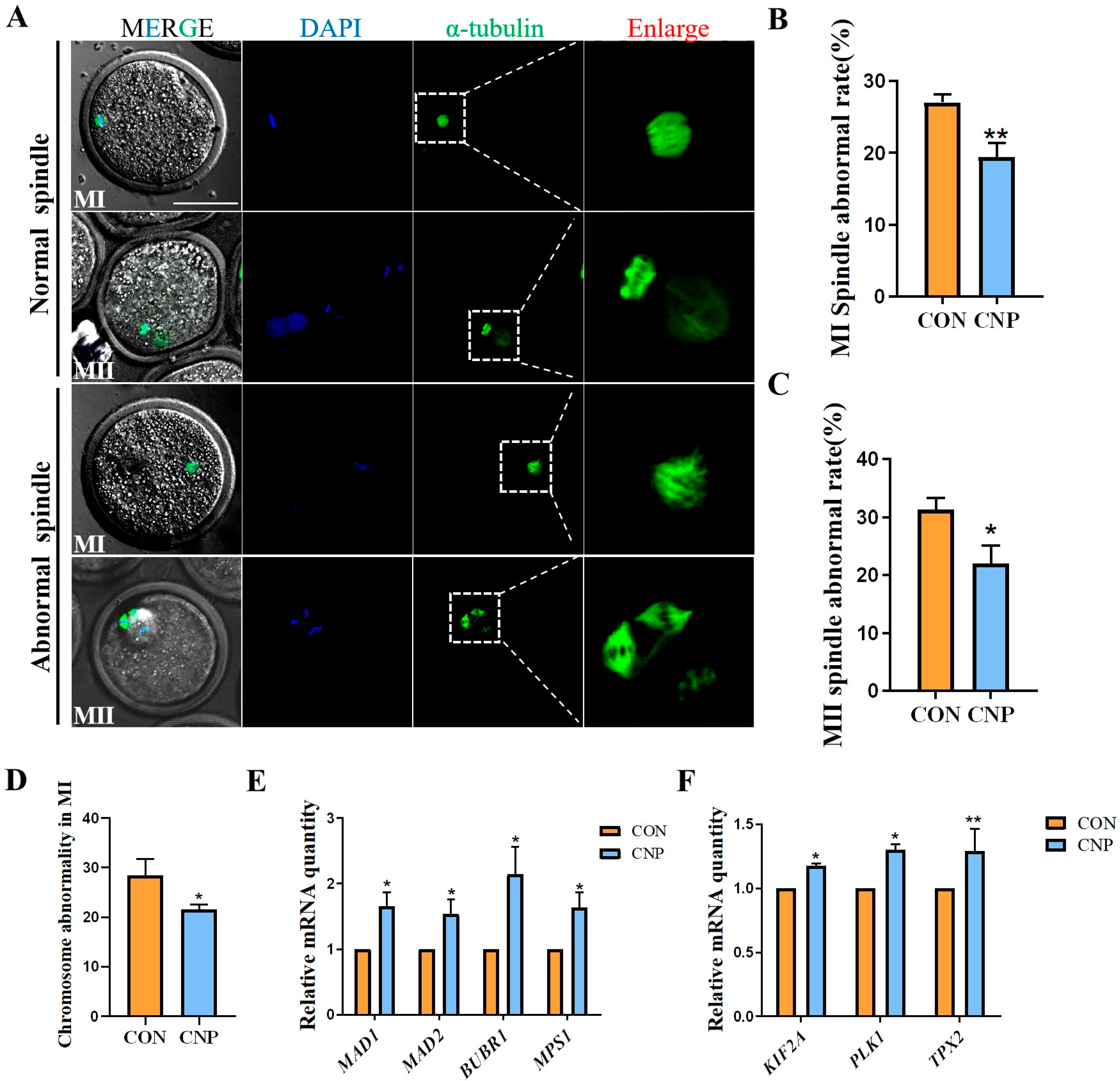
3.3. CNP Treatment before IVM Improved Spindle Assembly in Oocytes
3.4. CNP Treatment before IVM Enhanced the Antioxidant Capacity of Oocytes
3.5. CNP Treatment before IVM Improved Oocytes’ Mitochondrial Membrane Potential and Mitochondria Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krisher, R.L. In vivo and in vitro environmental effects on mammalian oocyte quality. Annu. Rev. Anim. Biosci. 2013, 1, 393–417. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Vaquer, A.; Barragan, M.; Rodriguez, A.; Vassena, R. Altered cytoplasmic maturation in rescued in vitro matured oocytes. Hum. Reprod. 2019, 34, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Su, Y.Q.; Sugiura, K.; Xia, G.; Eppig, J.J. Granulosa cell ligand NPPC and its receptor NPR2 maintain meiotic arrest in mouse oocytes. Science 2010, 330, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.P.; Ratzan, W.J.; Freudzon, M.; Mehlmann, L.M.; Krall, J.; Movsesian, M.A.; Wang, H.; Ke, H.; Nikolaev, V.O.; Jaffe, L.A. Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte. Development 2009, 136, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, H.; Liu, W.; Yang, Y.; Wang, X.; Zhang, Z.; Guo, Q.; Wang, C.; Xia, G. Natriuretic peptides improve the developmental competence of in vitro cultured porcine oocytes. Reprod. Biol. Endocrinol. 2017, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Wang, X. Effects of C-type natriuretic peptide on meiotic arrest and developmental competence of bovine oocyte derived from small and medium follicles. Sci. Rep. 2020, 10, 18213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wei, Q.; Cai, J.; Zhao, X.; Ma, B. Effect of C-Type Natriuretic Peptide on Maturation and Developmental Competence of Goat Oocytes Matured In Vitro. PLoS ONE 2015, 10, e0132318. [Google Scholar] [CrossRef]
- Romero, S.; Sanchez, F.; Lolicato, F.; Van Ranst, H.; Smitz, J. Immature Oocytes from Unprimed Juvenile Mice Become a Valuable Source for Embryo Production When Using C-Type Natriuretic Peptide as Essential Component of Culture Medium. Biol. Reprod. 2016, 95, 64. [Google Scholar] [CrossRef]
- Coticchio, G.; Dal Canto, M.; Mignini Renzini, M.; Guglielmo, M.C.; Brambillasca, F.; Turchi, D.; Novara, P.V.; Fadini, R. Oocyte maturation: Gamete-somatic cells interactions, meiotic resumption, cytoskeletal dynamics and cytoplasmic reorganization. Hum. Reprod. Update 2015, 21, 427–454. [Google Scholar] [CrossRef]
- Baena, V.; Terasaki, M. Three-dimensional organization of transzonal projections and other cytoplasmic extensions in the mouse ovarian follicle. Sci. Rep. 2019, 9, 1262. [Google Scholar] [CrossRef]
- Marchais, M.; Gilbert, I.; Bastien, A.; Macaulay, A.; Robert, C. Mammalian cumulus-oocyte complex communication: A dialog through long and short distance messaging. J. Assist. Reprod. Genet. 2022, 39, 1011–1025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.; Feng, X.; Zhang, S.; Xu, X.; Li, L.; Niu, S.; Bo, Y.; Wang, C.; Li, Z.; et al. Oocyte-derived microvilli control female fertility by optimizing ovarian follicle selection in mice. Nat. Commun. 2021, 12, 2523. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, M.; Crichton, J.H.; Playfoot, C.J.; Adams, I.R. Oocyte development, meiosis and aneuploidy. Semin. Cell Dev. Biol. 2015, 45, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Amaro, A.C.; Samora, C.P.; Holtackers, R.; Wang, E.; Kingston, I.J.; Alonso, M.; Lampson, M.; McAinsh, A.D.; Meraldi, P. Molecular control of kinetochore-microtubule dynamics and chromosome oscillations. Nat. Cell Biol. 2010, 12, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Lara-Gonzalez, P.; Pines, J.; Desai, A. Spindle assembly checkpoint activation and silencing at kinetochores. Semin. Cell Dev. Biol. 2021, 117, 86–98. [Google Scholar] [CrossRef]
- Trebichalska, Z.; Kyjovska, D.; Kloudova, S.; Otevrel, P.; Hampl, A.; Holubcova, Z. Cytoplasmic maturation in human oocytes: An ultrastructural study dagger. Biol. Reprod. 2021, 104, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Cree, L.M.; Samuels, D.C.; de Sousa Lopes, S.C.; Rajasimha, H.K.; Wonnapinij, P.; Mann, J.R.; Dahl, H.H.; Chinnery, P.F. A reduction of mitochondrial DNA molecules during embryogenesis explains the rapid segregation of genotypes. Nat. Genet. 2008, 40, 249–254. [Google Scholar] [CrossRef]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef]
- Barr, C.S.; Rhodes, P.; Struthers, A.D. C-type natriuretic peptide. Peptides 1996, 17, 1243–1251. [Google Scholar] [CrossRef]
- Walther, T.; Stepan, H. C-type natriuretic peptide in reproduction, pregnancy and fetal development. J. Endocrinol. 2004, 180, 17–22. [Google Scholar] [CrossRef]
- Hsueh, A.J.; Kawamura, K.; Cheng, Y.; Fauser, B.C. Intraovarian control of early folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Santiquet, N.; Papillon-Dion, E.; Djender, N.; Guillemette, C.; Richard, F.J. New elements in the C-type natriuretic peptide signaling pathway inhibiting swine in vitro oocyte meiotic resumption. Biol. Reprod. 2014, 91, 16. [Google Scholar] [CrossRef] [PubMed]
- Clarke, H.J. Transzonal projections: Essential structures mediating intercellular communication in the mammalian ovarian follicle. Mol. Reprod. Dev. 2022, 89, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, L.; El-Hayek, S.; Carvalho, K.F.; Wang, W.; Yang, Q.; Granados-Aparici, S.; Mondadori, R.; Bordignon, V.; Clarke, H.J. Epidermal growth factor receptor signaling uncouples germ cells from the somatic follicular compartment at ovulation. Nat. Commun. 2021, 12, 1438. [Google Scholar] [CrossRef] [PubMed]
- El-Hayek, S.; Yang, Q.; Abbassi, L.; FitzHarris, G.; Clarke, H.J. Mammalian Oocytes Locally Remodel Follicular Architecture to Provide the Foundation for Germline-Soma Communication. Curr. Biol. 2018, 28, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Severson, A.F.; von Dassow, G.; Bowerman, B. Oocyte Meiotic Spindle Assembly and Function. Curr. Top. Dev. Biol. 2016, 116, 65–98. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Zhou, C.; Yuan, M.; Miao, Y.; Zhao, X.; Ma, B. Effect of C-type natriuretic peptide on maturation and developmental competence of immature mouse oocytes in vitro. Reprod. Fertil. Dev. 2017, 29, 319–324. [Google Scholar] [CrossRef]
- Sun, S.C.; Kim, N.H. Spindle assembly checkpoint and its regulators in meiosis. Hum. Reprod. Update 2012, 18, 60–72. [Google Scholar] [CrossRef]
- Marston, A.L.; Wassmann, K. Multiple Duties for Spindle Assembly Checkpoint Kinases in Meiosis. Front. Cell Dev. Biol. 2017, 5, 109. [Google Scholar] [CrossRef]
- Wu, Z.; Pan, B.; Qazi, I.H.; Yang, H.; Guo, S.; Yang, J.; Zhang, Y.; Zeng, C.; Zhang, M.; Han, H.; et al. Melatonin Improves In Vitro Development of Vitrified-Warmed Mouse Germinal Vesicle Oocytes Potentially via Modulation of Spindle Assembly Checkpoint-Related Genes. Cells 2019, 8, 1009. [Google Scholar] [CrossRef]
- Akera, T. Tubulin post-translational modifications in meiosis. Semin. Cell Dev. Biol. 2023, 137, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Yang, X.; Liu, K. Treatment of cattle oocytes with C-type natriuretic peptide before in vitro maturation enhances oocyte mitochondrial function. Anim. Reprod. Sci. 2021, 225, 106685. [Google Scholar] [CrossRef] [PubMed]
- Soto-Heras, S.; Menendez-Blanco, I.; Catala, M.G.; Izquierdo, D.; Thompson, J.G.; Paramio, M.T. Biphasic in vitro maturation with C-type natriuretic peptide enhances the developmental competence of juvenile-goat oocytes. PLoS ONE 2019, 14, e0221663. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, M.; Singh, S.; Singh, A.P.; Dasgupta, S. Mitochondrial fusion and fission: The fine-tune balance for cellular homeostasis. FASEB J. 2021, 35, e21620. [Google Scholar] [CrossRef]
- Sanchez, F.; Romero, S.; De Vos, M.; Verheyen, G.; Smitz, J. Human cumulus-enclosed germinal vesicle oocytes from early antral follicles reveal heterogeneous cellular and molecular features associated with in vitro maturation capacity. Hum. Reprod. 2015, 30, 1396–1409. [Google Scholar] [CrossRef]
- Persad, K.L.; Lopaschuk, G.D. Energy Metabolism on Mitochondrial Maturation and Its Effects on Cardiomyocyte Cell Fate. Front. Cell Dev. Biol. 2022, 10, 886393. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Classification | Gene | Primer Seq (5′-3′) | Fragment Size (bp) | Annealing Temperature |
|---|---|---|---|---|
| Reference gene | ACTB | F: CGCAGACAGGATGCAGAAAG | 148 | 60 °C |
| R: GCTGATCCACATCTGCTGGA | ||||
| TZP assembly-related genes | DAAM1 | F: TGATGTCACCTGCGAGTTGG | 78 | 60 °C |
| R: CGCTCCGGGTACAATGGAAT | ||||
| FSCN1 | F: AAGTACTGGACGCTGACGAC | 133 | ||
| R: CCGTCACGAACTTGCCATTG | ||||
| MYO10 | F: TGGAGTCGCTCAACTTCGAC | 118 | ||
| R: TTGAAGCTAGGCTTCTCCGC | ||||
| Spindle assembly-related genes | KIF2A | F: GCACAACAGAATGCACGTAG | 132 | 60 °C |
| R: GTTTGTAGCATCAACATCCTGGG | ||||
| PLK1 | F: GATGCTTCAGCCAGATCCCA | 200 | ||
| R: CGGGGTTCTCCATGCCTTTAT | ||||
| TPX2 | F: GGAGAGGTGCCCAAGTTCAA | 283 | ||
| R:GCTGAAAAGGTTCCTGAACGATAA | ||||
| SAC-related genes | MAD1 | F: TGGAGGGAGGATCTGGACTG | 226 | 60 °C |
| R: ATCTCGCGCTCGTAGTTCCT | ||||
| MAD2 | F: TGCTTTTGAAACGAGTGGCG | 175 | ||
| R: ACCAAATGAGAAGAACTCGGC | ||||
| BUBR1 | F: TCCCAGCACAGACAATTCCA | 168 | ||
| R: CCCTACACGGCTGATTGGAG | ||||
| MPS1 | F: CGGCAGATTCCAGAGCAGAA | 140 | ||
| R: TCATCCAAGGCACTGTTGCT | ||||
| Mitochondrial oxidative metabolism-related genes | PGC-1α | F: AAGCCAACCAAGATAACC | 157 | 60 °C |
| R: TACAACTCAGACTGCTCGGG | ||||
| SIRT1 | F: GGCTTACAGGGCCTATCCAG | 124 | ||
| R: CACCAAACAGAAGGTTATCTCGG | ||||
| NRF1 | F: AGGCTGGGGCAAAGAAAG | 303 | ||
| R: CCAACCTGGATAAGCGAGAC | ||||
| NRF2 | F: CCAACTACTCCCAGGTAGCCC | 227 | ||
| R: AGCAGTGGCAACCTGAACG | ||||
| Mitochondria fusion and division-related genes | DRP1 | F: GAGAAGAAAATGGAGTTGAAGCA | 232 | 60 °C |
| R: CACCTACAGGCACCTTGGTC | ||||
| FIS1 | F: GGAACTACCGGCTCAAGGAAT | 206 | ||
| R: GGACACAGCAAGTCCGATGA | ||||
| OPA1 | F: AGCTTCTGACCTACTTCTCTTGT | 110 | ||
| R: TCTCTTTGTCTGACACCTTTCTGT | ||||
| MFN1 | F: CGGGCACAGATGTCACTA | 151 | ||
| R: GGCTTGGAAAGTCGCTCA | ||||
| MFN2 | F: TGCATGAAACTGGCGCGAT | 282 | ||
| R: TGATGCCCCTCACTTTGGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, R.; Pan, M.; Yin, L.; Zhang, Y.; Tang, Y.; Lu, S.; Gao, Y.; Wei, Q.; Han, B.; Ma, B. C-Type Natriuretic Peptide Pre-Treatment Improves Maturation Rate of Goat Oocytes by Maintaining Transzonal Projections, Spindle Morphology, and Mitochondrial Function. Animals 2023, 13, 3880. https://doi.org/10.3390/ani13243880
Xu R, Pan M, Yin L, Zhang Y, Tang Y, Lu S, Gao Y, Wei Q, Han B, Ma B. C-Type Natriuretic Peptide Pre-Treatment Improves Maturation Rate of Goat Oocytes by Maintaining Transzonal Projections, Spindle Morphology, and Mitochondrial Function. Animals. 2023; 13(24):3880. https://doi.org/10.3390/ani13243880
Chicago/Turabian StyleXu, Rui, Menghao Pan, Lu Yin, Yiqian Zhang, Yaju Tang, Sihai Lu, Yan Gao, Qiang Wei, Bin Han, and Baohua Ma. 2023. "C-Type Natriuretic Peptide Pre-Treatment Improves Maturation Rate of Goat Oocytes by Maintaining Transzonal Projections, Spindle Morphology, and Mitochondrial Function" Animals 13, no. 24: 3880. https://doi.org/10.3390/ani13243880