
Winter Epibiotic Community of the Red King Crab Paralithodes camtschaticus in Sayda Bay (Barents Sea)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
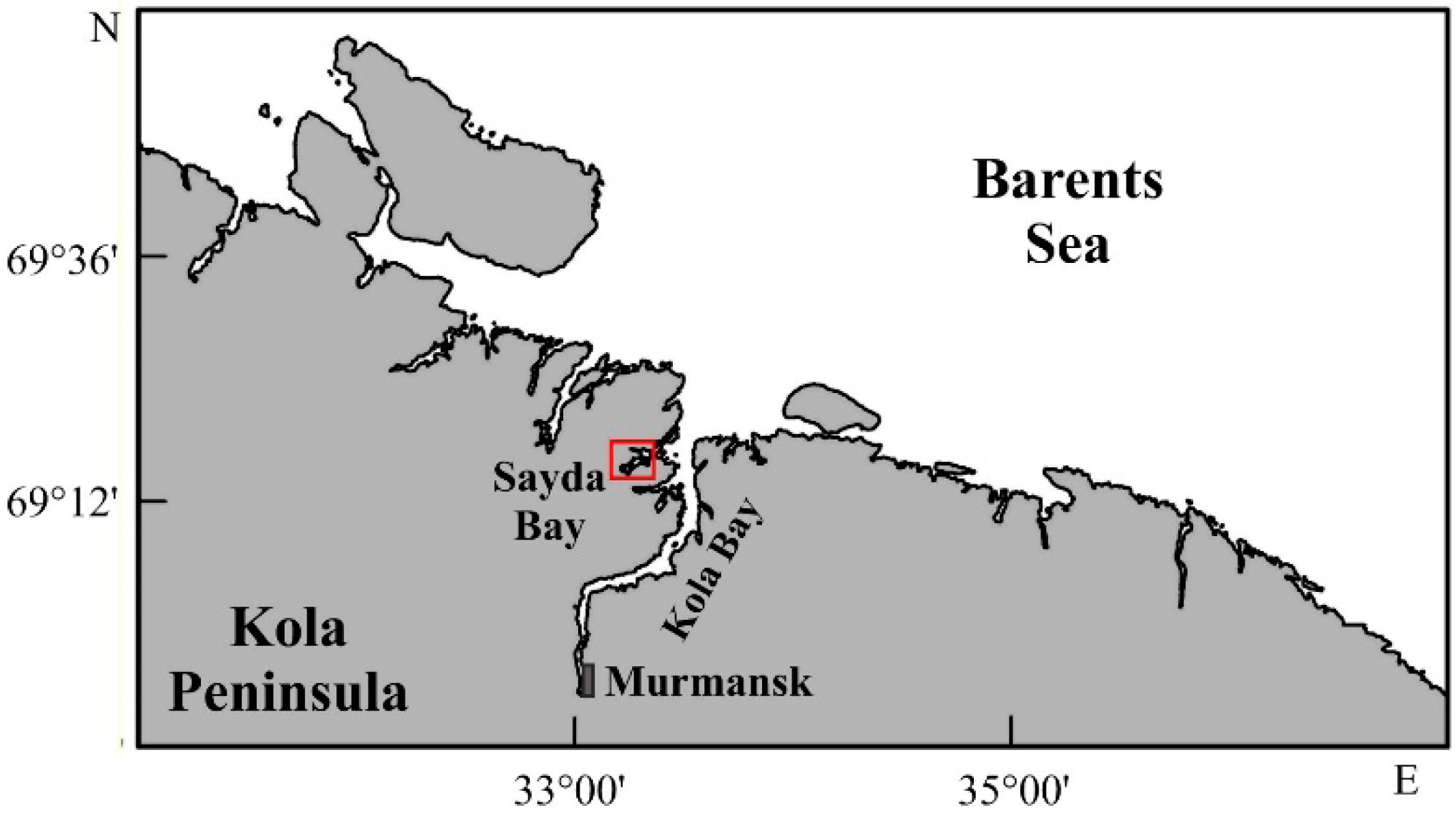
2.1. Study Area
2.2. Field Sampling and Data Analysis
2.3. Statistical Analysis
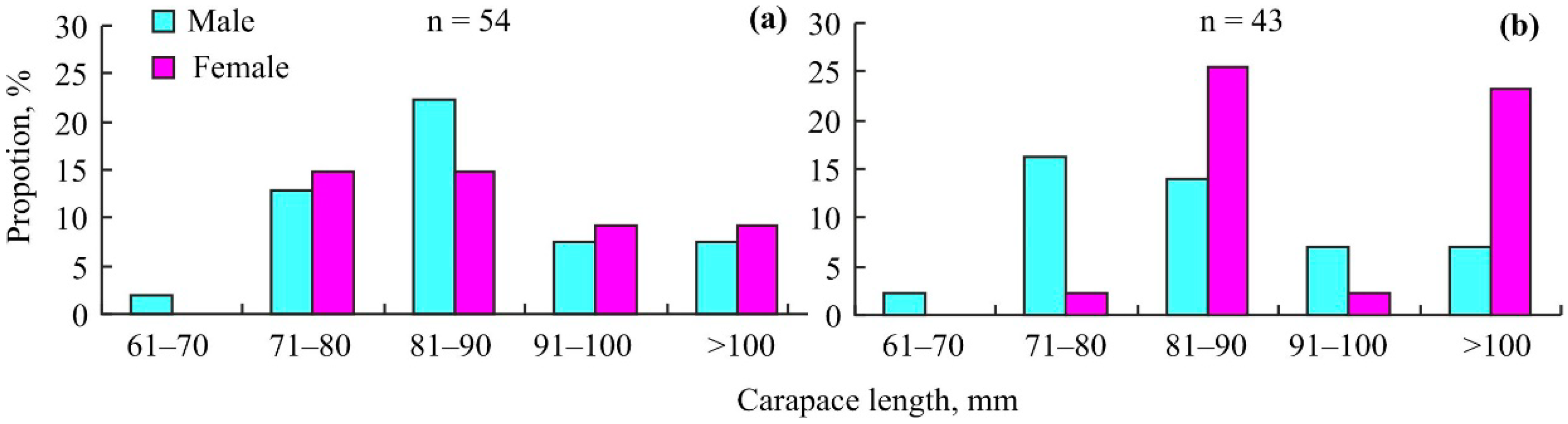
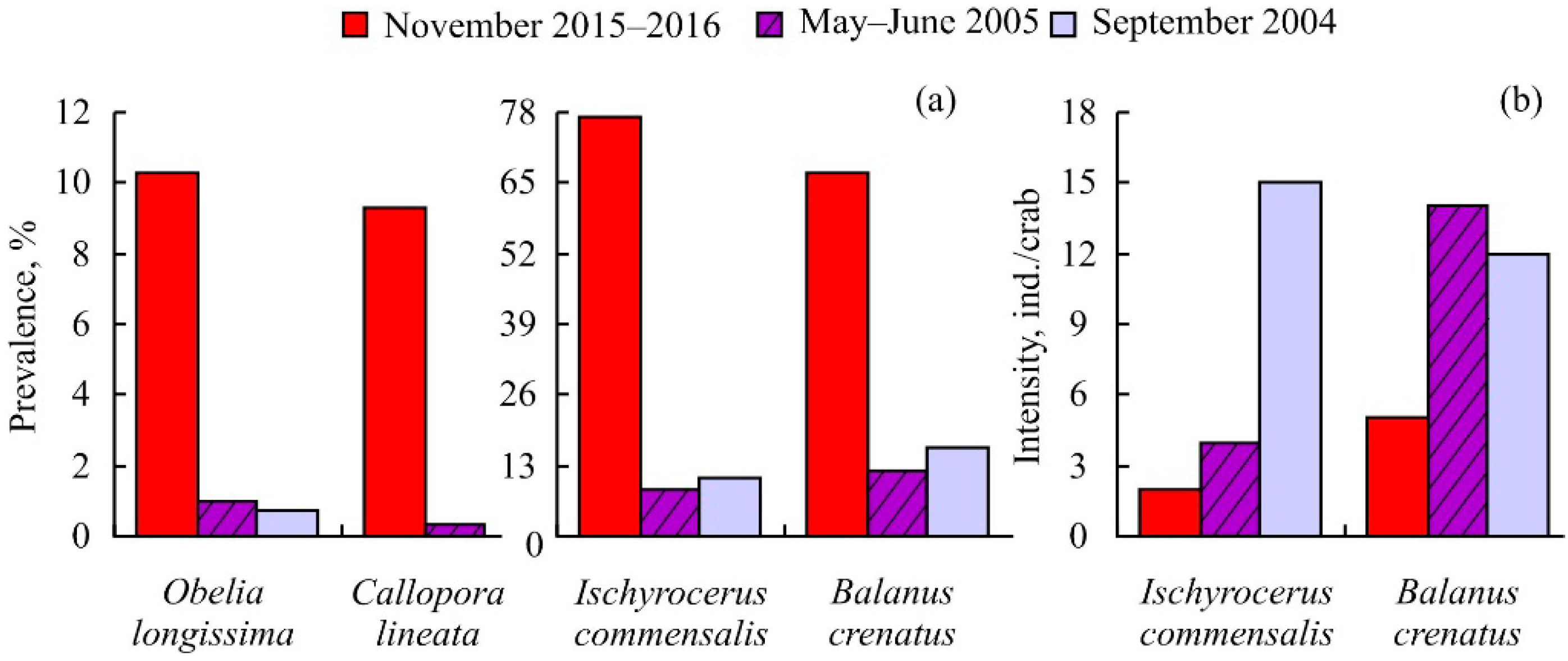
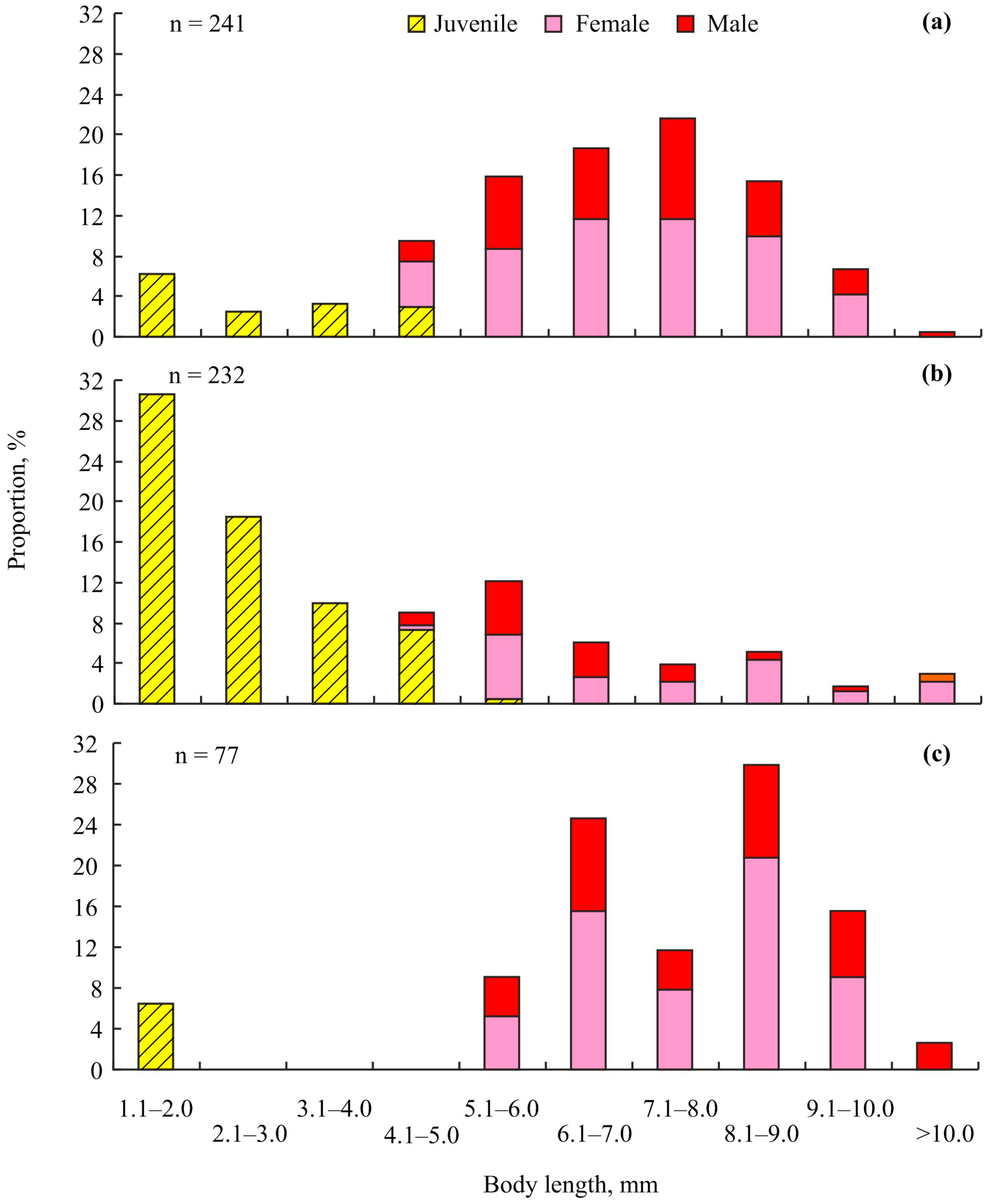
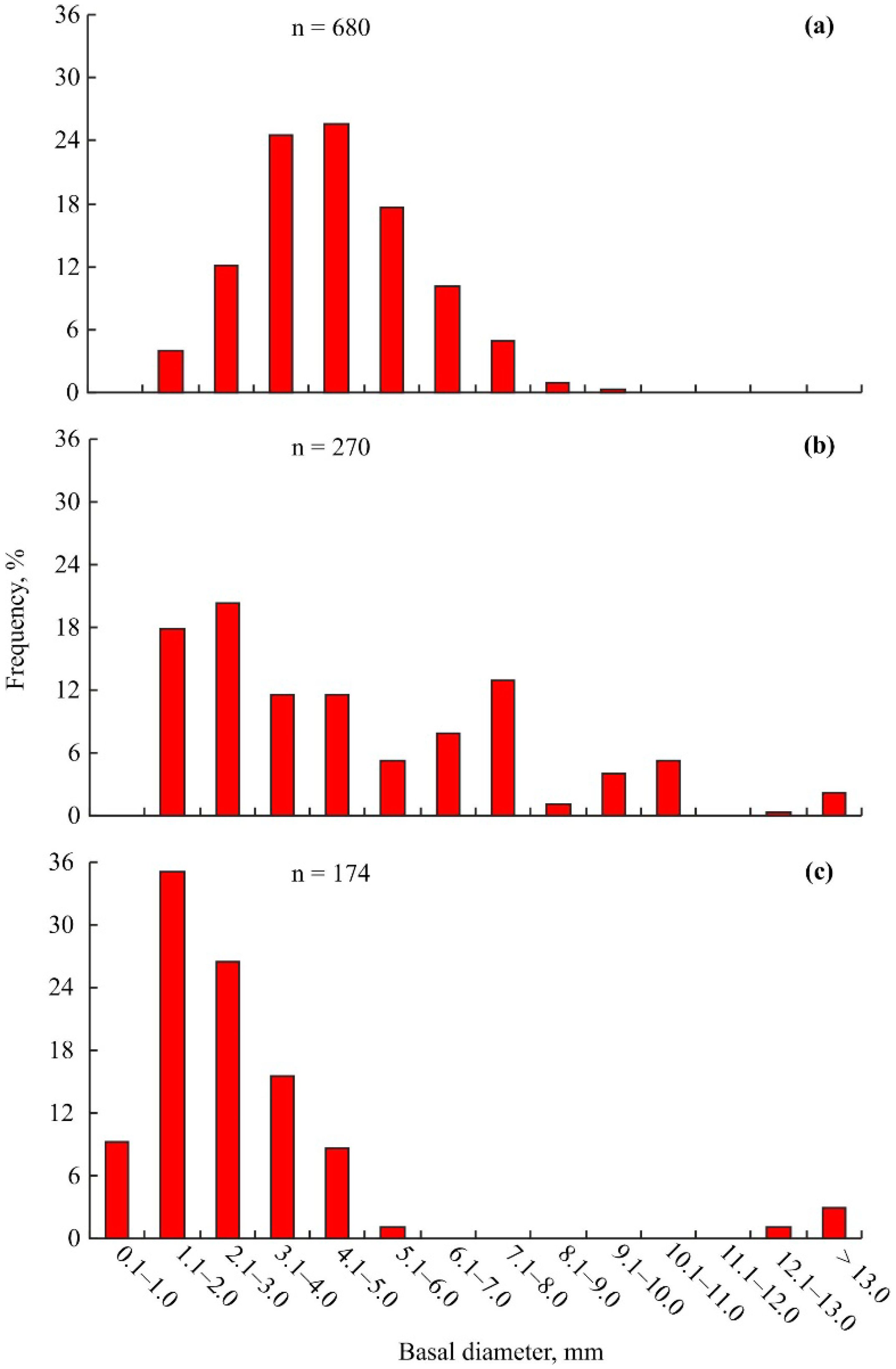
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (Red King) Crab in the Barents Sea: Peculiarities of Biology, Perspectives of Fishery; KSC RAS Press: Apatity, Russia, 2002. (In Russian) [Google Scholar]
- Bakanev, S.V.; Stesko, A.V. Red king crab. In Materials Justifying Changes to the Previously Approved Total Allowable Catch in Fishing Areas in Inland Seas of the Russian Federation, on the Continental Shelf of the Russian Federation, in the Exclusive Economical Zone of the Russian Federation, in the Azov and Caspian Seas in 2021; FGBUN VNIRO (Polar Branch): Murmansk, Russia, 2021; pp. 2–16. (In Russian) [Google Scholar]
- Dvoretsky, V.G.; Dvoretsky, A.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Renewal of the recreational red king crab fishery in Russian waters of the Barents Sea: Potential benefits and costs. Mar. Policy 2022, 136, 104916. [Google Scholar] [CrossRef]
- Abelló, P.; Villanueva, R.; Gili, J.M. Epibiosis in deep-sea crab populations as indicator of biological and behavioural characteristics of the host. J. Mar. Biol. Assoc. UK 1990, 70, 687–695. [Google Scholar] [CrossRef]
- Parapar, J.; Fernández, L.; González-Gurriarán, E.; Muíno, R. Epibiosis and masking material in the spider crab Maja squinado (Decapoda: Majidae) in the Ria de Arousa (Galicia, NW Spain). Cah. Biol. Mar. 1997, 38, 221–234. [Google Scholar]
- Dick, M.H.; Donaldson, W.E.; Vining, I.W. Epibionts of the tanner crab Chionoecetes bairdi in the region of Kodiak Island, Alaska. J. Crust. Biol. 1998, 18, 519–528. [Google Scholar] [CrossRef]
- Fernandez-Leborans, G. Epibiosis in Crustacea: An overview. Crustaceana 2010, 83, 549–640. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Distribution of amphipods Ischyrocerus on the red king crab, Paralithodes camtschaticus: Possible interactions with the host in the Barents Sea. Estuar. Coast. Shelf Sci. 2009, 82, 390–396. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef]
- Mantelatto, F.L.; O’Brien, J.J.; Biagi, R. Parasites and symbionts of crabs from Ubatuba Bay, Sao Paulo State, Brazil. Compar. Parasitol. 2003, 70, 211–214. [Google Scholar] [CrossRef]
- Williams, J.D.; McDermott, J.J. Hermit crab biocoenoses; a worldwide review of the diversity and natural history of hermit crab associates. J. Exp. Mar. Biol. Ecol. 2004, 305, 1–128. [Google Scholar] [CrossRef]
- Fernandez-Leborans, G.; Davila, P.; Cerezo, E.; Contreras, C. Epibiosis and hyperepibiosis on Pagurus bernhardus (Crustacea: Decapoda) from the west Coast of Scotland. J. Mar. Biol. Assoc. UK 2013, 93, 1351–1362. [Google Scholar] [CrossRef]
- Campos, J.; Ribas, F.; Bio, A.; Freitas, V.; Souza, A.T.; van der Veer, H.W. Body condition and energy content of the shore crab Carcinus maenas L. in a temperate coastal system: The cost of barnacle epibiosis. Biofouling 2022, 38, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Bakay, Y.; Kuzmin, S.A.; Utevsky, S.Y. Ecological and parasitologic investigations on the Barents Sea red king crab Paralithodes camtschaticus (the first results). ICES CM 1998, 4, 14. [Google Scholar]
- Jansen, P.A.; Mackenzie, K.; Hemmingsen, W. Some parasites and commensals of red king crab Paralithodes camtschaticus in the Barents Sea. Bull. Europ. Assoc. Fish Pathol. 1998, 18, 46–49. [Google Scholar]
- Hemmingsen, W.; Jansen, P.A.; MacKenzie, K. Crabs, leeches and trypanosomes: An unholy trinity? Mar. Pollut. Bull. 2005, 50, 336–339. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Some aspects of the biology of the amphipods Ischyrocerus anguipes associated with the red king crab, Paralithodes camtschaticus, in the Barents Sea. Polar Biol. 2009, 32, 463–469. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Matishov, G.G. (Ed.) Kola Bay: Oceanography, Ecosystems, and Pollutants; KSC RAS Press: Apatity, Russia, 1997. (In Russian) [Google Scholar]
- Dvoretskiy, A.G.; Dvoretskiy, V.G. Epibionts of the red king crab Paralithodes camtschaticus (Tilesius, 1815) in Sayda Inlet of Kola Bay. Inland Water Biol. 2009, 2, 276–279. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs; Alaska Sea Grant College Program, University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population biology of Ischyrocerus commensalis, a crab-associated amphipod, in the southern Barents Sea: A multi-annual summer study. Mar. Ecol. 2011, 32, 498–508. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Pereladov, M.V. Some aspects of distribution and behavior of red king crabs (Paralithodes camtschaticus) in the Barents Sea shallow coastal water. Tr. VNIRO 2003, 142, 103–119. (In Russian) [Google Scholar]
- Talberg, N.B. Comparative characteristics of the features of red king crab migrations in coastal waters Barents and Okhotsk Seas. Tr. VNIRO 2005, 144, 91–101. [Google Scholar]
- Kuznetsov, V.V. The Biology of Abundant and the Most Common Species of Crustaceans in the Barents and White Seas; Nauka: Moscow, Russia, 1964. (In Russian) [Google Scholar]
- McGaw, I.J. Epibionts of sympatric species of Cancer crabs in Barkley sound, British Columbia. J. Crust. Biol. 2006, 26, 85–93. [Google Scholar] [CrossRef]
- Savoie, L.; Miron, J.; Biron, M. Fouling community of the snow crab Chionoecetes opilio in Sydney Bight, Canada: Preliminary observations in relation to sampling period and depth/geographical location. Cah. Biol. Mar. 2007, 48, 347–359. [Google Scholar]
- Sainte-Marie, B.; Berube, I.; Brillon, S.; Hazel, F. Observations on the growth of the sculptured shrimp Sclerocrangon boreas (Decapoda: Caridea). J. Crustac. Biol. 2006, 26, 55–62. [Google Scholar] [CrossRef]
- Kuris, A.M.; Blau, S.F.; Paul, A.J.; Shields, J.D.; Wickham, D.E. Infestation by brood symbionts and their impact on egg mortality in the red king crab, Paralithodes camtschatica, in Alaska: Geographic and temporal variation. Can. J. Fish. Aquat. Sci. 1991, 48, 559–568. [Google Scholar] [CrossRef]
- Klitin, A.K. The Red King Crab along the Shores of Sakhalin and the Kuril Islands: Biology, Distribution, and Functional Structure of the Area; Natsrybresursy: Moscow, Russia, 2003. (In Russian) [Google Scholar]
- Costa, T.M.; Christofoletti, R.A.; Pinheiro, M.A. Epibionts on Arenaeus cribrarius (Brachyura: Portunidae) from Brazil. Zoologia 2010, 27, 387–394. [Google Scholar] [CrossRef]
- Newell, R.C.; Branch, G.M. The influence of temperature on the maintenance of metabolic energy balance in marine invertebrates. Adv. Mar. Biol. 1980, 17, 329–396. [Google Scholar]
- Gosselin, L.A.; Qian, P.Y. Juvenile mortality in benthic marine invertebrates. Mar. Ecol. Prog. Ser. 1997, 146, 265–282. [Google Scholar] [CrossRef]
- Reitzel, A.M.; Miner, B.G.; McEdward, L.R. Relationships between spawning date and larval development time for benthic marine invertebrates: A modeling approach. Mar. Ecol. Prog. Ser. 2004, 280, 13–23. [Google Scholar] [CrossRef]
- Firtha, L.B.; Knights, A.M.; Bella, S.S. Air temperature and winter mortality: Implications for the persistence of the invasive mussel, Perna viridis, in the intertidal zone of the south-eastern United States. J. Exp. Mar. Biol. Ecol. 2011, 400, 250–256. [Google Scholar] [CrossRef]
- Paul, J.; Paul, A. Encrusting Barnacles as Ageable Tags on Gulf of Alaska Chionoecetes bairdi (Decapoda); Alaska Sea Grant College Program, University of Alaska: Fairbanks, AK, USA, 1986. [Google Scholar]
- Jeffries, W.B.; Voris, H.K.; Poovachiranon, S. Age of the mangrove crab Scylla serrata at colonization by stalked barnacles of the genus Octolasmis. Biol. Bull. 1992, 182, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Lovrich, G.A.; Calcagno, J.A.; Smith, B.D. The barnacle Notobalanus flosculus as an indicator of the intermolt period of the male lithodid crab Paralomis granulosa. Mar. Biol. 2003, 143, 143–156. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Prevalence, % | Mean Intensity, ind. per Crab | ||
|---|---|---|---|---|
| Mean ± SE | Min | Max | ||
| Hydrozoa | ||||
| Coryne hincksii Bonnevie, 1898 | 1.03 | – | – | – |
| Obelia longissima (Pallas, 1766) | 10.31 | – | – | – |
| Nemertini | ||||
| Nemertini g. sp.1 | 1.03 | 1.0 ± 0.0 | 1 | 1 |
| Polychaeta | ||||
| Circeis armoricana Saint-Joseph, 1894 | 4.12 | 1.5 ± 0.3 | 1 | 2 |
| Bivalvia | ||||
| Mytilus edulis L., 1758 | 3.09 | 1.7 ± 0.3 | 1 | 2 |
| Copepoda | ||||
| Ectinosoma normani Scott T. & A., 1894 | 3.09 | 1.0 ± 0.0 | 1 | 1 |
| Mesochra pygmaea (Claus, 1863) | 4.12 | 1.0 ± 0.0 | 1 | 1 |
| Tisbe furcata (Baird, 1837) | 4.12 | 1.0 ± 0.0 | 1 | 1 |
| Amphipoda | ||||
| Ischyrocerus commensalis Chevreux, 1900 | 77.32 | 1.7 ± 0.1 | 1 | 4 |
| Cirripedia | ||||
| Balanus crenatus Brugiere, 1789 | 67.01 | 14.3 ± 2.7 | 1 | 94 |
| Bryozoa | ||||
| Callopora lineata (L., 1767) | 9.28 | – | – | – |
| Scrupocellaria arctica (Smitt, 1868) | 1.03 | – | – | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, A.G.; Dvoretsky, V.G. Winter Epibiotic Community of the Red King Crab Paralithodes camtschaticus in Sayda Bay (Barents Sea). Animals 2024, 14, 100. https://doi.org/10.3390/ani14010100
Dvoretsky AG, Dvoretsky VG. Winter Epibiotic Community of the Red King Crab Paralithodes camtschaticus in Sayda Bay (Barents Sea). Animals. 2024; 14(1):100. https://doi.org/10.3390/ani14010100
Chicago/Turabian StyleDvoretsky, Alexander G., and Vladimir G. Dvoretsky. 2024. "Winter Epibiotic Community of the Red King Crab Paralithodes camtschaticus in Sayda Bay (Barents Sea)" Animals 14, no. 1: 100. https://doi.org/10.3390/ani14010100