
Digestibility of Conventional and Novel Dietary Lipids in Channel Catfish Ictalurus punctatus

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Preparation
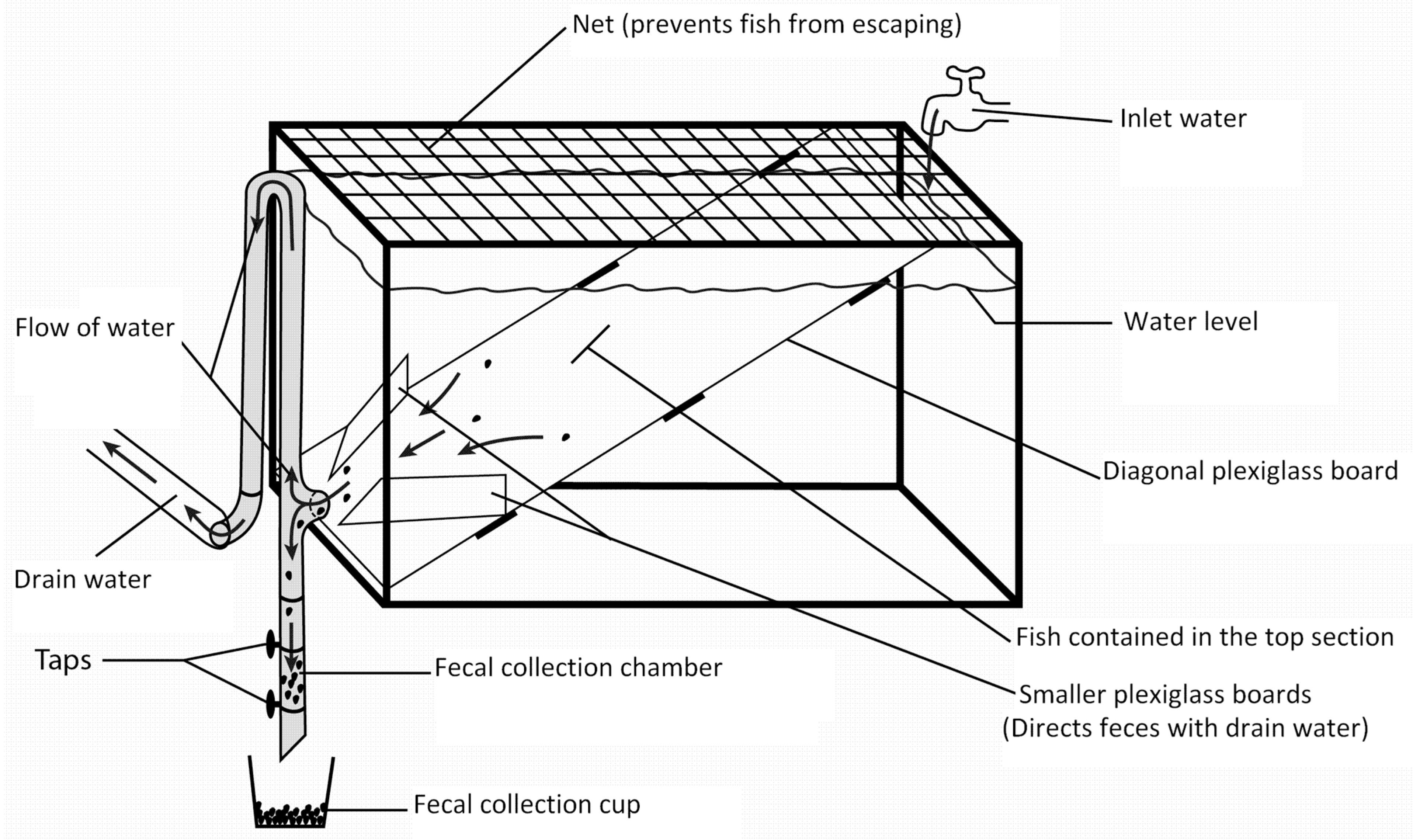
2.2. Fecal Collection System, Feeding and Fecal Collection
2.3. Proximate and Fatty Acid Analysis of Diets and Feces
2.4. Analysis of Inert Marker (Yttrium) and Calculation of Apparent Digestibility Coefficients (ADC)
3. Data Analysis
4. Results
4.1. Fatty Acid Composition of Feces
4.2. Apparent Digestibility Coefficients for Crude Protein, Lipid and Fatty Acids
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Agricultural Statistics Service (NASS). Catfish Production; Agricultural Statistics Board, United States Department of Agriculture (USDA): Washington, DC, USA, 2023; ISSN 1948-271X.
- Turchini, G.M.; Torstensen, B.E.; Ng, W.-K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Hajen, W.; Beames, R.M.; Higgs, D.; Dosanjh, B. Digestibility of various feedstuffs by post-juvenile Chinook salmon (Oncorhynchus tshawaytscha) in seawater. 1. Validation of technique. Aquaculture 1993, 112, 321–332. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient Requirements of Fish and Shrimp; National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- Stickney, R.R.; Andrews, J.W. Effects of Dietary Lipids on Growth, Food Conversion, Lipid and Fatty Acid Composition of Channel Catfish. J. Nutr. 1972, 102, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Lochmann, R.; Phillips, H. Comparison of Rice Bran Oil, Poultry Fat, and Cod Liver Oil as Supplemental Lipids in Feeds for Channel Catfish and Golden Shiner. J. Appl. Aquac. 1995, 5, 47–55. [Google Scholar] [CrossRef]
- O’neal, C.C.; Kohler, C.C. Effect of Replacing Menhaden Oil with Catfish Oil on the Fatty Acid Composition of Juvenile Channel Catfish, Ictalurus punctatus. J. World Aquac. Soc. 2008, 39, 62–71. [Google Scholar] [CrossRef]
- Robinson, E.H.; Li, M.H. Channel catfish, Ictalurus punctatus, nutrition in the United States: A historical perspective. J. World Aquac. Soc. 2019, 51, 93–118. [Google Scholar] [CrossRef]
- Martins, D.A.; Valente, L.M.; Lall, S.P. Apparent digestibility of lipid and fatty acids in fish oil, poultry fat and vegetable oil diets by Atlantic halibut, Hippoglossus hippoglossus L. Aquaculture 2009, 294, 132–137. [Google Scholar] [CrossRef]
- Hua, K.; Bureau, D.P. Development of a model to estimate digestible lipid content of salmonid fish feeds. Aquaculture 2009, 286, 271–276. [Google Scholar] [CrossRef]
- Sigurgisladottir, S.; Lall, S.P.; Parrish, C.C.; Ackman, R.G. Cholestane as a digestibility marker in the absorption of polyunsaturated fatty acid ethyl esters in Atlantic salmon. Lipids 1992, 27, 418–424. [Google Scholar] [CrossRef]
- Ng, W.-K.; Campbell, P.J.; Dick, J.R.; Bell, J.G. Interactive effects of dietary palm oil concentration and water temperature on lipid digestibility in rainbow trout, Oncorhynchus mykiss. Lipids 2003, 38, 1031–1038. [Google Scholar] [CrossRef]
- Menoyo, D.; Lopez-Bote, C.J.; Bautista, J.M.; Obach, A. Growth, digestibility and fatty acid utilization in large atlantic salmon (Salmo salar) fed varying levels of n-3 and saturated fatty acids. Aquaculture 2003, 225, 295–307. [Google Scholar] [CrossRef]
- Satoh, S.; Poe, W.E.; Wilson, R.P. Effect of Dietary n-3 Fatty Acids on Weight Gain and Liver Polar Lipid Fatty Acid Composition of Fingerling Channel Catfish. J. Nutr. 1989, 119, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Trushenski, J.T.; Rombenso, A.N.; Jackson, C.J. Reevaluating Polyunsaturated Fatty Acid Essentiality in Channel Catfish. N. Am. J. Aquac. 2020, 82, 265–277. [Google Scholar] [CrossRef]
- Suja, B.; Lochmann, R.; Sink, T.; Phillips, H.; Chen, R. Effect of diets supplemented with soybean, flaxseed, or menhaden fish oil on the growth, feed utilization, immune status, and sensory properties of channel catfish in a recirculating system at 22 °C. J. Appl. Aquac. 2012, 24, 16–31. [Google Scholar] [CrossRef]
- Faukner, J.; Rawles, S.D.; Proctor, A.A.; Sink, T.D.; Chen, R.; Phillips, H.; Lochmann, R.T. The effects of diets containing standard soybean oil, soybean oil enhanced with conjugated linoloeic acids, menhaden fish oil, or an algal docosahexaenoic acid supplement on channel catfish performance, body composition, sensory evaluation, and storage characteristics. N. Am. J. Aquac. 2013, 75, 252–265. [Google Scholar]
- Troesch, B.; Eggersdorfer, M.; Laviano, A.; Rolland, Y.; Smith, A.D.; Warnke, I.; Weimann, A.; Calder, P.C. Expert Opinion on Benefits of Long-Chain Omega-3 Fatty Acids (DHA and EPA) in Aging and Clinical Nutrition. Nutrients 2020, 12, 2555. [Google Scholar] [CrossRef]
- Twibell, R.G.; Wilson, R.P. Effects of dietary conjugated linoleic acids and total dietary lipid concentrations on growth responses of juvenile channel catfish, Ictalurus punctatus. Aquaculture 2003, 221, 621–628. [Google Scholar] [CrossRef]
- Manning, B.B.; Li, M.H.; Robinson, E.H.; Peterson, B.C. Enrichment of channel catfish (Ictalurus punctatus) fillets with conjugated linoleic acid and omega-3 fatty acids by dietary manipulation. Aquaculture 2006, 261, 337–342. [Google Scholar] [CrossRef]
- Faukner, J.; Rawles, S.D.; Sink, T.D.; Lochmann, R.; Proctor, A.; Chen, R.; Phillips, H. The Effects of Diets Containing Standard Soybean Oil, Soybean Oil Enhanced with Conjugated Linoleic Acids, Menhaden Fish Oil, or an Algal Docosahexaenoic Acid Supplement on Juvenile Channel Catfish Performance, Hematology, Body Composition, and Nonspecific Immune Response. N. Am. J. Aquac. 2015, 77, 217–229. [Google Scholar] [CrossRef]
- McCrorie, A.T.; Keaveney, E.M.; Wallace, J.M.W.; Livingston, M.B.E. Human health effects of conjugated linoleic acid from milk and supplements. Nutr. Res. Rev. 2011, 24, 206–207. [Google Scholar] [CrossRef]
- Moon, H.Y.; Gatlin, D.M. Total sulfur amino acid requirement of juvenile red drum, Sciaenops ocellatus. Aquaculture 1991, 95, 97–106. [Google Scholar] [CrossRef]
- Li, M.H.; Robinson, E.H.; Peterson, B.C.; Bates, T.D. Growth and Feed Efficiency of Juvenile Channel Catfish Reared at Different Water Temperatures and Fed Diets Containing Various Levels of Fish Meal. North Am. J. Aquac. 2008, 70, 347–352. [Google Scholar] [CrossRef]
- Lochmann, S.E.; Perschbacher, P.W.; Merry, G.E.; Fijan, N. Aggressive Biting among Channel Catfish in Pool Studies. Progress. Fish-Culturist 1998, 60, 119–126. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis, 16th ed.; AOAC: Arlington, VA, USA, 1995. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Morrison, R.W.; Smith, L.M. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride-methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Barrows, F.T.; Gaylord, T.G.; Sealey, W.; Rawles, S.D. Database of Nutrient Digestibility’s of Traditional and Novel Feed Ingredients for Trout and Hybrid Striped Bass. 2011. Available online: http://www.ars.usda.gov/Main/docs.htm?docid=21905 (accessed on 1 November 2015).
- Zar, J.H. Biostatistical Analysis; Prentice-Hall International: London, UK, 2009. [Google Scholar]
- Bell, J.G.; Koppe, W. Welfare and health of fish fed vegetable oils as alternative lipid sources to fish oil. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G.M., Ng, W.-K., Tocher, D.R., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 21–59. [Google Scholar]
- Gunasekera, M.R.; Lelarasamee, K.; De Silva, S.S. Lipid and fatty acid digestibility of three oil types in the Australian shortfin eel, Anguilla australis. Aquaculture 2002, 203, 335–347. [Google Scholar] [CrossRef]
- Bahurmiz, O.M.; Ng, W.-K. Effects of dietary palm oil source on growth, tissue fatty acid composition and nutrient digestibility of red hybrid tilapia, Oreochromis sp., raised from stocking to marketable size. Aquaculture 2007, 262, 382–392. [Google Scholar] [CrossRef]
- Yılmaz, H.A.; Eroldoğan, O.T. Effects of fish oil substitution with two different vegetable oil classes on fatty acid digestibility in juvenile European sea bass, Dicentrarchus labrax. Turk. J. Fish. Aquat. Sci. 2015, 15, 1–12. [Google Scholar] [CrossRef]
- Ng, W.-K.; Sigholt, T.; Bell, J.G. The influence of environmental temperature on the apparent nutrient and fatty acid digestibility in Atlantic salmon (Salmo salar L.) fed finishing diets containing different blends of fish oil, rapeseed oil and palm oil. Aquac. Res. 2004, 35, 1228–1237. [Google Scholar] [CrossRef]
- Francis, D.S.; Turchini, G.M.; Jones, P.L.; De Silva, S.S. Effects of fish oil substitution with a mix blend vegetable oil on nutrient sdigestibility in Murray cod, Maccullochella peelii peelii. Aquaculture 2007, 269, 447–455. [Google Scholar] [CrossRef]
- Austreng, E.; Skrede, A.; Eldegard, Å. Effect of Dietary Fat Source on the Digestibility of Fat and Fatty Acids in Rainbow Trout and Mink. Acta Agric. Scand. 1979, 29, 119–126. [Google Scholar] [CrossRef]
- Caballero, J.M.; Obach, A.; Roselund, G.; Montero, D.; Gisvold, M.; Izquierdo, M.S. Impact of different dietary lipid sources on growth lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar] [CrossRef]
- Grisdale-Helland, B.; Ruytera, B.; Rosenland, G.; Obach, A.; Helland, S.J.; Sandberg, M.G.; Standal, H.; Røsjø, C. Influence of high contents of dietary soybean oil on growth, feed utilization, tissue fatty acid composition, heart histology and standard oxygen consumption of Atlantic salmon (Salmo salar) raised at two temperatures. Aquaculture 2002, 207, 311–329. [Google Scholar] [CrossRef]
- Rowan, J.P.; Durrance, K.L.; Combs, G.E.; Fisher, L.Z. The Digestive Tract of the Pig; UF/IFAS Extension Publication No. AS 23; University of Florida: Gainesville, FL, USA, 2015. [Google Scholar]
- Jørgensen, H.; Gabert, V.M.; Hedemann, M.S.; Jensen, S.K. Digestion of Fat Does Not Differ in Growing Pigs Fed Diets Containing Fish Oil, Rapeseed Oil or Coconut Oil. J. Nutr. 2000, 130, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Duran-Montgé, P.; Lizardo, R.; Torrallardona, D.; Esteve-Garcia, E. Fat and fatty acid digestibility of different fat sources in growing pigs. Livest. Sci. 2007, 109, 66–69. [Google Scholar] [CrossRef]
- Jørgensen, H.; Jakobsen, K.; Eggum, B.O. The influence of different protein, fat and mineral levels on the digestibility of fat and fatty acids measure at the terminal ileum and in feces of growing pigs. Acta Agric. Scand. 1992, 42, 177–184. [Google Scholar]
- Gjellesvik, R.D.; Lombardo, D.; Walther, B.T. Pancreatic bile salt dependent lipase from cod (Gadus morhua): Purification and properties. Biochim. et Biophys. Acta 1992, 1124, 123–124. [Google Scholar] [CrossRef]
- Olsen, E.R.; Henderson, R.J.; Ringø, E. The digestion and selective absorption of dietary fatty acids in Arctic charr, Salvelinus alpinus. Aquac. Nutr. 1998, 4, 13–21. [Google Scholar] [CrossRef]
- Koven, M.W.; Henderson, R.J.; Sargent, J.R. Lipid digestion in turbot (Scophthalmus maximus): In-vivo and in-vitro studies of the lipolytic activity in various segments of the digestive tract. Aquaculture 1997, 151, 155–171. [Google Scholar] [CrossRef]
- Twibell, R.G.; Watkins, B.A.; Rogers, L.; Brown, P.B. Effects of dietary conjugated linoleic acids on hepatic and muscle lipids in hybrid striped bass. Lipids 2000, 35, 155–161. [Google Scholar] [CrossRef]
- Kennedy, S.R.; Campbell, P.J.; Porter, A.; Tocher, D.R. Influence of dietary conjugated linoleic acid (CLA) on lipid and fatty acid composition in liver and flesh of Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. B 2005, 141, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.R.; Leaver, M.J.; Campbell, P.J.; Zheng, X.; Dick, J.R.; Tocher, D.R. Influence of dietary oil content and conjugated linoleic acid (CLA) on lipid metabolism enzyme activities and gene expression in tissues of Atlantic salmon (Salmo salar). Lipids 2006, 41, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.R.; Roys, L.L. Behavioral interactions in juvenile channel catfish, Ictaluras punctatus. In Recent Developments in Catfish Aquaculture; Tave, D., Tucker, C.S., Eds.; Haworth Press: New York, NY, USA, 1990; pp. 363–381. [Google Scholar]


{kind=link}
| Ingredients | % |
|---|---|
| Soybean meal 1 | 50.0 |
| Cottonseed meal 1 | 10.0 |
| Wheat shorts 1 | 31.0 |
| Yttrium Oxide 2 | 0.5 |
| Vitamin premix 3 | 1.0 |
| Mineral premix 3 | 1.0 |
| Ethoxyquin | 0.0125 |
| Supplemental lipid 4 | 4.0 |
| Carboxymethylcellulose | 2.5 |
| Diet | Lipid | Protein | Dry Matter | Ash |
|---|---|---|---|---|
| SBO | 8.2 | 32.9 | 93.4 | 7.6 |
| CLA-SBO | 7.6 | 32.2 | 93.6 | 7.8 |
| COO | 7.9 | 32.3 | 95.8 | 7.4 |
| FXO | 7.3 | 32.7 | 93.8 | 7.3 |
| MFO | 7.3 | 32.4 | 93.3 | 7.1 |
| PF | 7.7 | 33.0 | 95.1 | 7.3 |
| Diets | ||||||
|---|---|---|---|---|---|---|
| FATTY ACID | SBO | CLA-SBO | COO | FXO | MFO | PF |
| 14:0 | 0.1 ± 0.1 | ND | 0.8 ± 0.03 | 0.1 ± 0.002 | 5.7 ± 0.1 | 0.7 ± 0.04 |
| 16:0 | 14.7 ± 0.4 | 16.1 ± 0.2 | 19.3 ± 0.02 | 11.1 ± 0.1 | 19.7 ± 0.1 | 22.7 ± 0.1 |
| 18:0 | 4.0 ± 0.004 | 4.7 ± 0.01 | 5.0 ± 0.01 | 4.0 ± 0.02 | 3.8 ± 0.02 | 5.0 ± 0.02 |
| 20:0 | 0.3 ± 0.01 | 0.4 ± 0.02 | ND | ND | ND | ND |
| 22:0 | 0.4 ± 0.01 | 0.4 ± 0.004 | ND | ND | ND | ND |
| ∑ SFA 2 | 19.4 ± 0.3 | 21.6 ± 0.1 | 25.0 ± 0.1 | 15.3 ± 0.1 | 29.2 ± 0.1 | 28.4 ± 0.2 |
| 16:1n-7 | 0.1 ± 0.1 | 0.1 ± 0.1 | 1.8 ± 0.001 | ND | 7.1 ± 0.02 | 4.9 ± 0.02 |
| 18:1n-9 | 20.6 ± 0.1 | 22.7 ± 0.1 | 36.2 ± 0.02 | 19.2 ± 0.2 | 12.0 ± 0.01 | 30.7 ± 0.2 |
| 18:1n-7 | 1.5 ± 0.02 | 1.6 ± 0.05 | 1.7 ± 0.05 | 0.8 ± 0.02 | 2.6 ± 0.02 | 1.8 ± 0.01 |
| 20:1n-9 | 0.1 ± 0.1 | 0.5 ± 0.002 | 0.9 ± 0.004 | 0.1 ± 0.1 | 0.4 ± 0.01 | 0.1 ± 0.12 |
| 22:1n-11 | ND | ND | ND | ND | 1.2 ± 0.002 | ND |
| ∑ MUFA 3 | 22.2 ± 0.1 | 24.9 ± 0.1 | 40.6 ± 0.03 | 20.1 ± 0.3 | 23.3 ± 0.01 | 37.5 ± 0.3 |
| CLA-9c,11t | ND | 1.0 ± 0.03 | ND | ND | ND | ND |
| CLA-10t,12c | ND | 0.9 ± 0.01 | ND | ND | ND | ND |
| ∑CLA isomers 4 | ND | 1.9 ± 0.04 | ND | ND | ND | ND |
| 18:2n-6 | 52.7 ± 0.1 | 47.0 ± 0.1 | 30.6 ± 0.1 | 30.7 ± 0.2 | 24.6 ± 0.2 | 31.2 ± 0.1 |
| 18:3n-3 | 5.6 ± 0.04 | 4.5 ± 0.01 | 3.0 ± 0.03 | 33.9 ± 0.4 | 4.4 ± 0.1 | 2.9 ± 0.04 |
| 20:2n-6 | ND | ND | 0.5 ± 0.003 | ND | ND | ND |
| 20:4n-6 | ND | ND | 0.3 ± 0.02 | ND | 0.7 ± 0.0 | ND |
| 20:5n-3 | ND | ND | ND | ND | 8.2 ± 0.04 | ND |
| 22:5n-3 | ND | ND | ND | ND | 1.3 ± 0.01 | ND |
| 22:6n-3 | ND | ND | ND | ND | 8.3 ± 0.002 | ND |
| ∑ n-3 5 | 5.6 ± 0.04 | 4.5 ± 0.01 | 3.0 ± 0.03 | 33.9 ± 0.4 | 22.3 ± 0.1 | 2.9 ± 0.04 |
| ∑ n-6 6 | 52.7 ± 0.1 | 47.0 ± 0.1 | 31.4 ± 0.1 | 30.7 ± 0.2 | 25.3 ± 0.2 | 31.2 ± 0.1 |
| ∑ n-3 LC-PUFA 7,8 | ND | ND | ND | ND | 17.8 ± 0.03 | ND |
| ∑ n-6 LC-PUFA 7,9 | ND | ND | 0.8 ± 0.04 | ND | 0.7 ± 0.0 | ND |
| n-3/n-6 | 0.1 ± 0.001 | 0.1 ± 0.0001 | 0.1 ± 0.001 | 1.1 ± 0.02 | 0.9 ± 0.007 | 0.1 ± 0.001 |
| Diets | ||||||||
|---|---|---|---|---|---|---|---|---|
| Fatty Acid | SBO | CLA-SBO | COO | FXO | MFO | PF | PSE 2 | Pr > F |
| 14:0 | 0.7 bc | 0.6 c | 1.0 b | 0.9 bc | 3.2 a | 0.9 bc | 0.1 | <0.001 |
| 16:0 | 20.0 d | 20.5 cd | 22.7 bc | 22.3 cd | 25.0 b | 30.0 a | 0.9 | <0.001 |
| 18:0 | 5.7 | 6.2 | 6.4 | 6.9 | 6.2 | 7.6 | 0.6 | 0.388 |
| 20:0 | 0.4 | 0.5 | 0.4 | 0.3 | 0.4 | 0.3 | 0.06 | 0.213 |
| 22:0 | 0.6 a | 0.6 a | 0.4 b | 0.5 b | 0.5 b | 0.3 c | 0.04 | 0.001 |
| ∑ SFA 3 | 27.4 b | 28.5 b | 30.9 b | 30.9 b | 35.3 a | 39.1 a | 1.4 | <0.001 |
| 16:1n-7 | 0.8 d | 1.0 cd | 1.4 c | 0.6 d | 3.2 a | 2.1 b | 0.2 | <0.001 |
| 18:1n-9 | 18.2 b | 18.6 b | 24.4 a | 18.8 b | 15.6 c | 20.3 b | 0.8 | <0.001 |
| 18:1n-7 | 2.1 b | 2.1 b | 2.3 b | 2.2 b | 2.8 a | 2.1 b | 0.1 | 0.019 |
| 20:1n-9 | 0.5 b | 0.5 b | 0.8 a | 0.5 b | 0.5 b | 0.4 b | 0.04 | <0.001 |
| 22:1n-11 | ND | 0.1 | ND | ND | 0.1 | 0.1 | 0.1 | 0.690 |
| ∑ MUFA 4 | 21.5 c | 22.5 c | 29.0 a | 22.1 bc | 22.3 bc | 25.2 b | 1.03 | 0.002 |
| CLA-9c,11t | ND | 0.5 | ND | ND | ND | ND | 0.03 | ----- |
| CLA-10t,12c | ND | 0.3 | ND | ND | ND | ND | 0.03 | ----- |
| ∑CLA isomers 5 | ND | 0.8 | ND | ND | ND | ND | 0.05 | ----- |
| 18:2n-6 | 45.6 a | 43.6 a | 35.6 bc | 37.0 b | 34.1 bc | 32.0 c | 1.2 | <0.001 |
| 18:3n-3 | 4.7 b | 4.3 bc | 3.4 d | 9.9 a | 3.8 cd | 3.1 d | 0.3 | <0.001 |
| 20:2n-6 | 0.2 | 0.1 | 0.4 | 0.1 | 0.1 | 0.1 | 0.1 | 0.056 |
| 20:4n-6 | 0.2 | 0.2 | 0.3 | ND | 0.1 | 0.2 | 0.1 | 0.876 |
| 20:5n-3 | ND | 0.1 b | ND | ND | 1.8 a | ND | 0.1 | <0.001 |
| 22:5n-3 | ND | ND | ND | ND | ND | ND | ----- | ----- |
| 22:6n-3 | ND | 0.2 b | 0.3 b | ND | 2.4 a | ND | 0.2 | <0.001 |
| ∑ n-3 6 | 4.7 c | 4.7 c | 3.7 cd | 9.9 a | 8.0 b | 3.1 d | 0.3 | <0.001 |
| ∑ n-6 7 | 46.0 a | 43.9 a | 36.3 bc | 37.1 b | 34.3 bc | 32.7 c | 1.2 | <0.001 |
| ∑ n-3 LC-PUFA 8,9 | ND | 0.3 b | 0.3 b | ND | 4.2 a | ND | 0.3 | <0.001 |
| ∑ n-6 LC-PUFA 8,10 | 0.4 | 0.2 | 0.9 | 0.1 | 0.2 | 0.3 | 0.2 | 0.081 |
| n-3/n-6 | 0.1 c | 0.1 c | 0.1 c | 0.3 a | 0.2 b | 0.1 c | 0.006 | <0.001 |
| Diets | ||||||||
|---|---|---|---|---|---|---|---|---|
| Fatty Acid | SBO | CLA-SBO | COO | FXO | MFO | PF | PSE 2 | Pr > F |
| lipid | 83.0 | 82.8 | 83.9 | 83.5 | 88.4 | 84.9 | 3.7 | 0.889 |
| 14:0 | 0.0 c | ----- | 53.9 b | 0.0 c | 82.3 a | 55.2 b | 2.8 | <0.001 |
| 16:0 | 50.1 | 57.7 | 53.8 | 33.4 | 59.2 | 55.9 | 7.0 | 0.178 |
| 18:0 | 46.4 | 55.4 | 48.2 | 41.9 | 47.1 | 49.4 | 9.9 | 0.957 |
| 20:0 | 52.7 | 57.7 | ----- | ----- | ----- | ----- | 4.7 | 0.496 |
| 22:0 | 40.6 | 47.5 | ----- | ----- | ----- | ----- | 6.4 | 0.489 |
| ∑SFAs 3 | 48.2 | 56.0 | 51.3 | 33.6 | 60.9 | 54.1 | 7.5 | 0.238 |
| 16:1n-7 | 0.0 c | 0.0 c | 68.5 b | ----- | 85.7 a | 86.0 a | 1.8 | <0.001 |
| 18:1n-9 | 67.8 b | 72.8 ab | 73.9 ab | 68.1 b | 58.4 c | 78.0 a | 2.9 | 0.007 |
| 18:1n-7 | 46.9 b | 57.2 ab | 48.6 b | 0.0 c | 66.0 a | 61.1 ab | 5.6 | <0.001 |
| 20:1n-9 | 0.0 c | 67.0 a | 66.0 a | 0.0 c | 58.2 b | 0.0 c | 2.4 | <0.001 |
| 22:1n-11 | ----- | ----- | ----- | ----- | 97.1 | ----- | 2.8 | ----- |
| ∑MUFAs 4 | 64.7 | 70.3 | 72.3 | 64.5 | 69.3 | 77.8 | 3.0 | 0.073 |
| CLA-9c,11t | ----- | 84.1 | ----- | ----- | ----- | ----- | 1.8 | ----- |
| CLA-10t,12c | ----- | 90.0 | ----- | ----- | ----- | ----- | 0.9 | ----- |
| ∑CLAisomers 5 | ----- | 86.9 | ----- | ----- | ----- | ----- | 1.1 | ----- |
| 18:2n-6 | 68.6 | 69.4 | 55.3 | 60.3 | 55.6 | 65.3 | 3.9 | 0.081 |
| 18:3n-3 | 69.9 bc | 68.3 bc | 56.5 d | 90.3 a | 73.0 b | 64.3 c | 2.5 | <0.001 |
| 20:2n-6 | ----- | ----- | 72.3 | ----- | ----- | ----- | 1.8 | ----- |
| 20:4n-6 | ----- | ----- | 58.5 b | ----- | 94.2 a | ----- | 6.6 | 0.019 |
| 20:5n-3 | ----- | ----- | ----- | ----- | 93.1 | ----- | 0.2 | ----- |
| 22:5n-3 | ----- | ----- | ----- | ----- | 100.0 | ----- | 0.0 | ----- |
| 22:6n-3 | ----- | ----- | ----- | ----- | 90.6 | ----- | 0.9 | ----- |
| ∑n-3 6 | 70.0 b | 70.0 b | 53.5 c | 90.3 a | 88.5 a | 64.3 b | 2.6 | <0.001 |
| ∑n-6 7 | 68.3 | 68.3 | 55.4 | 60.5 | 56.3 | 65.0 | 3.9 | 0.103 |
| ∑n-3 LC-PUFA 8,9 | ----- | ----- | ----- | ----- | 92.3 | ----- | 0.5 | ----- |
| ∑n-6 LC-PUFA 8,10 | ----- | ----- | 66.0 b | ----- | 94.2 a | ----- | 5.0 | 0.016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maina, A.; Lochmann, R.; Rawles, S.D.; Rosentrater, K. Digestibility of Conventional and Novel Dietary Lipids in Channel Catfish Ictalurus punctatus. Animals 2023, 13, 1456. https://doi.org/10.3390/ani13091456
Maina A, Lochmann R, Rawles SD, Rosentrater K. Digestibility of Conventional and Novel Dietary Lipids in Channel Catfish Ictalurus punctatus. Animals. 2023; 13(9):1456. https://doi.org/10.3390/ani13091456
Chicago/Turabian StyleMaina, Andrew, Rebecca Lochmann, Steven D. Rawles, and Kurt Rosentrater. 2023. "Digestibility of Conventional and Novel Dietary Lipids in Channel Catfish Ictalurus punctatus" Animals 13, no. 9: 1456. https://doi.org/10.3390/ani13091456