
Survival, Growth Performance, and Hepatic Antioxidant and Lipid Profiles in Infected Rainbow Trout (Oncorhynchus mykiss) Fed a Diet Supplemented with Dihydroquercetin and Arabinogalactan
 , , and
, , and 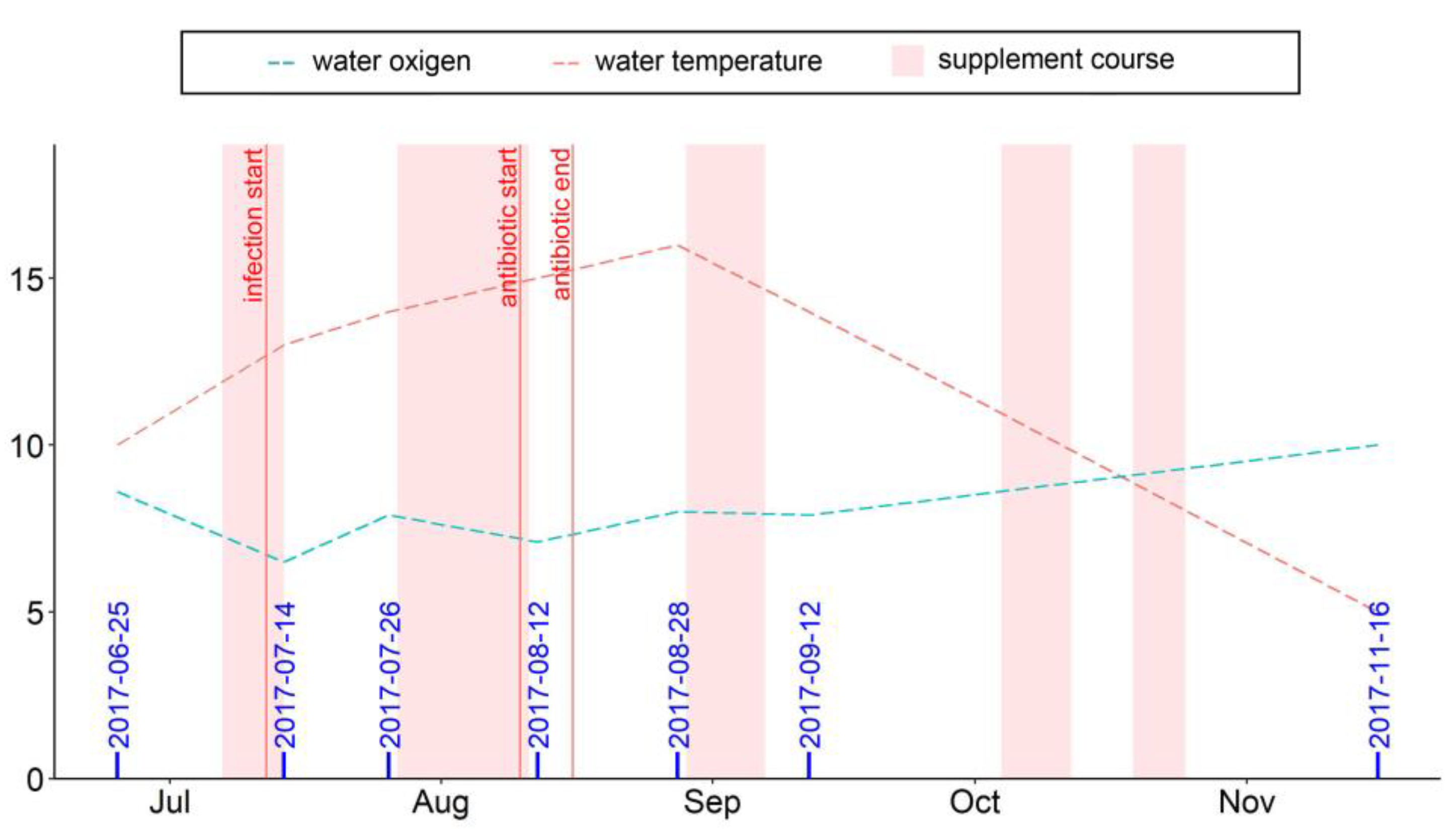{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Maintenance and Feeding
2.2. Fish Sampling
2.3. Biometric Indices and Mortality
2.4. Biochemical Assay
2.4.1. Antioxidant Analyses
2.4.2. Lipid Composition Analysis
2.5. Statistical Analyses
3. Results
3.1. Fish Survival and Growth
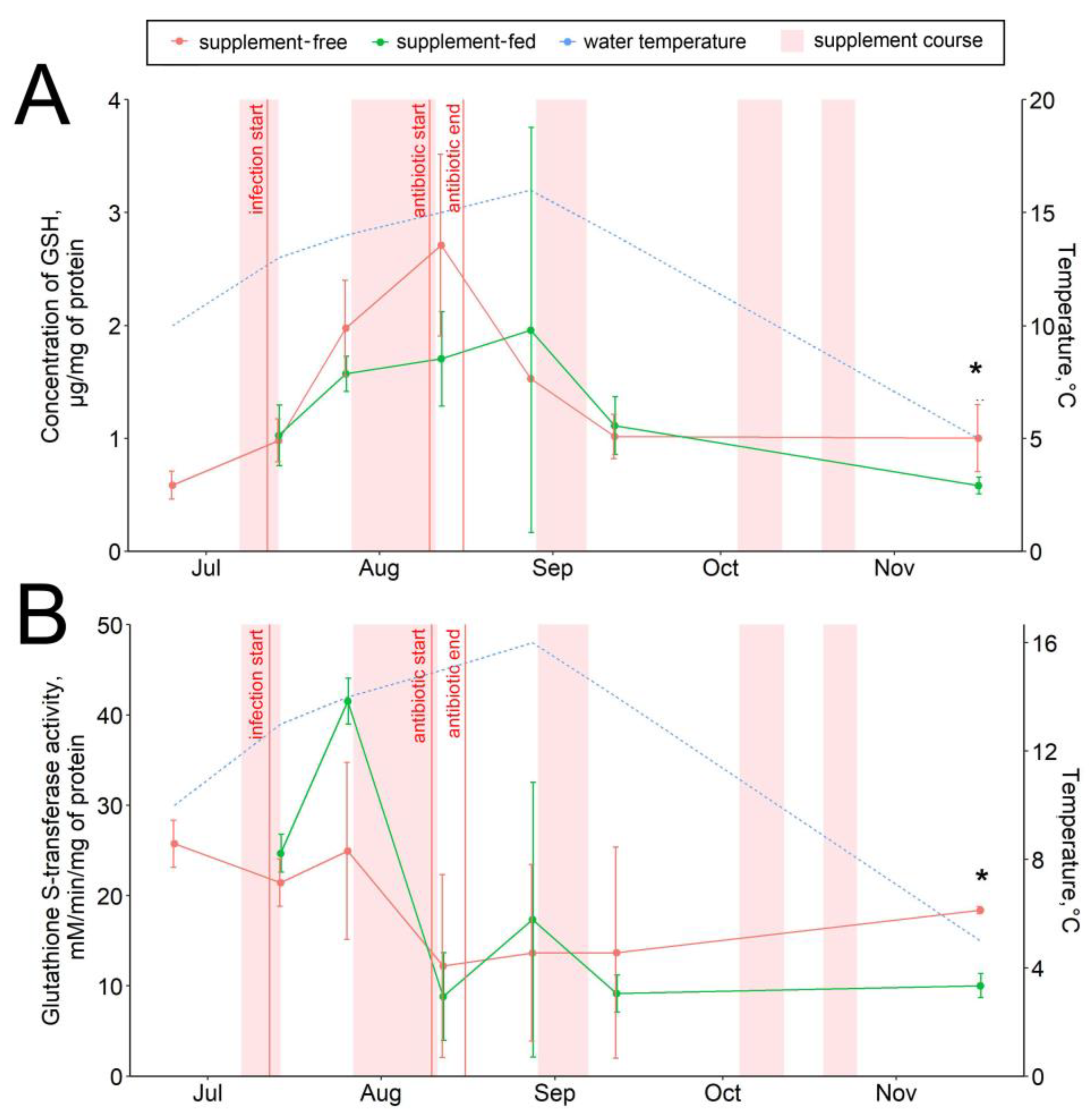
3.2. Hepatic Antioxidant Components
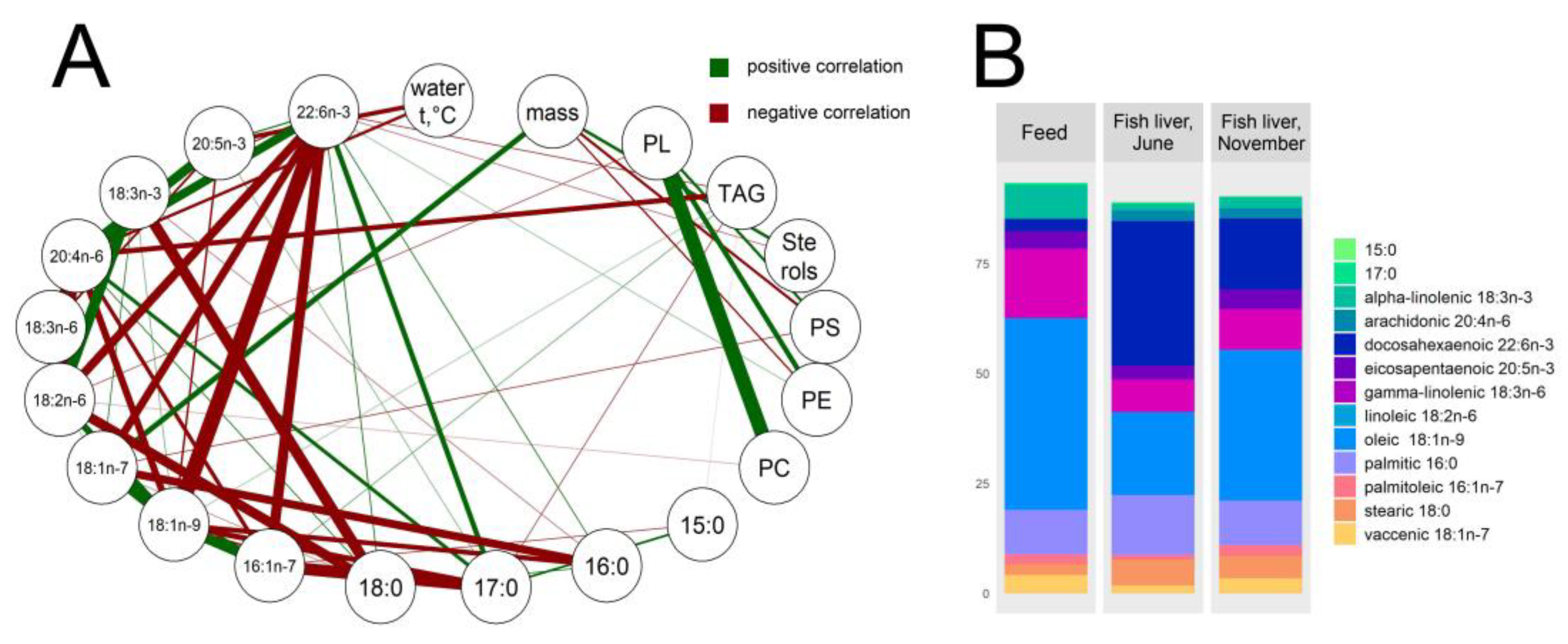
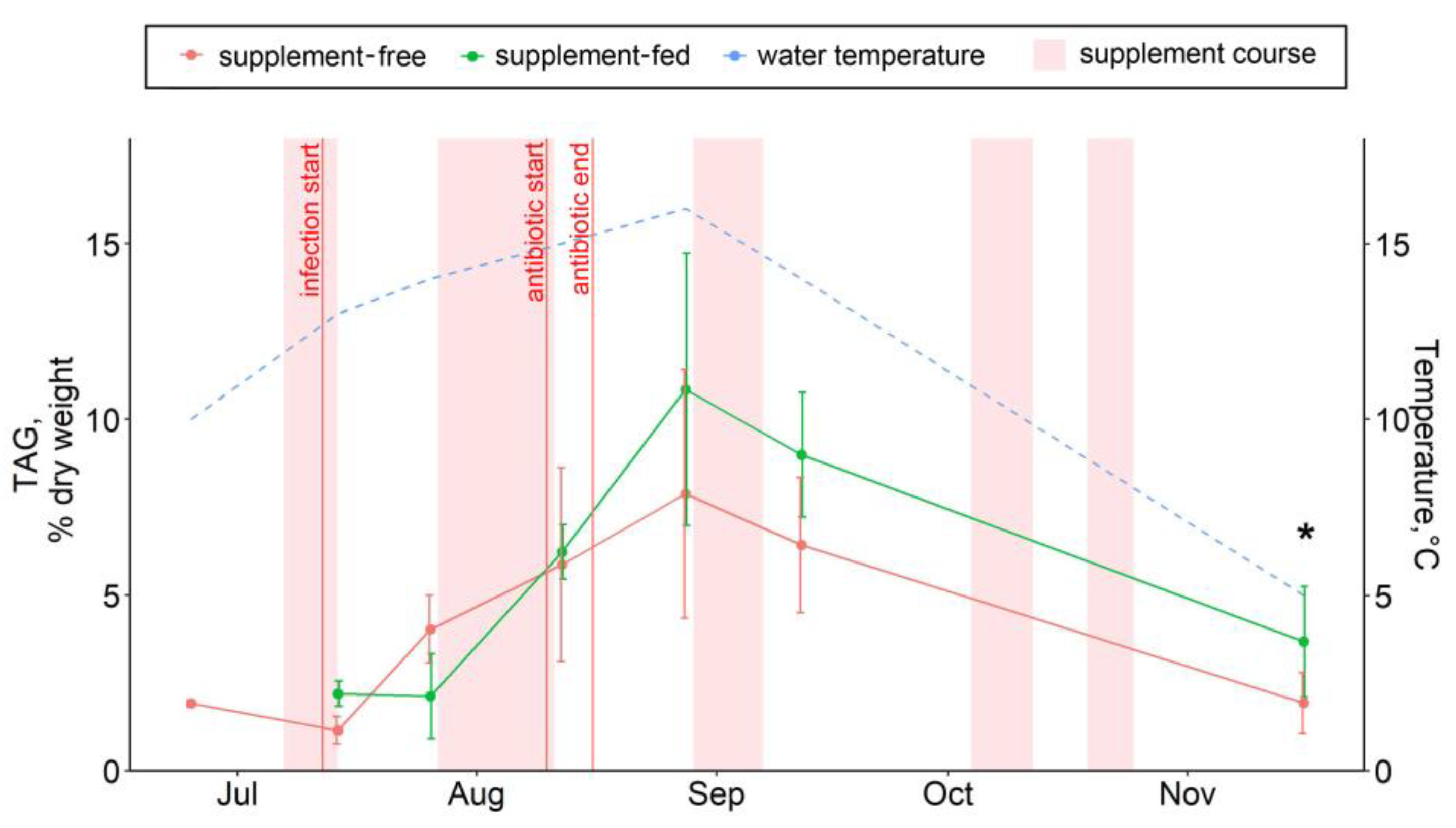
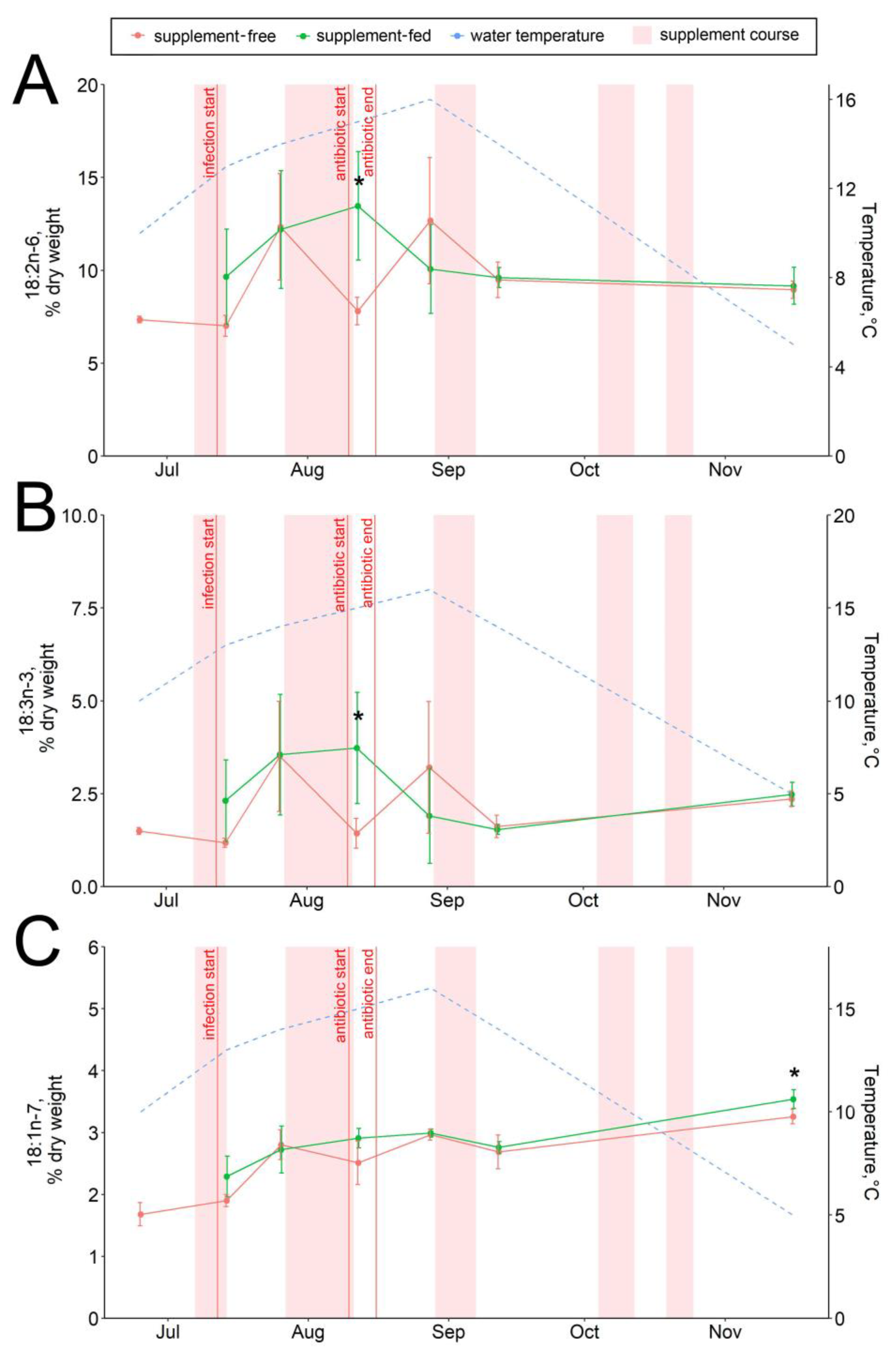
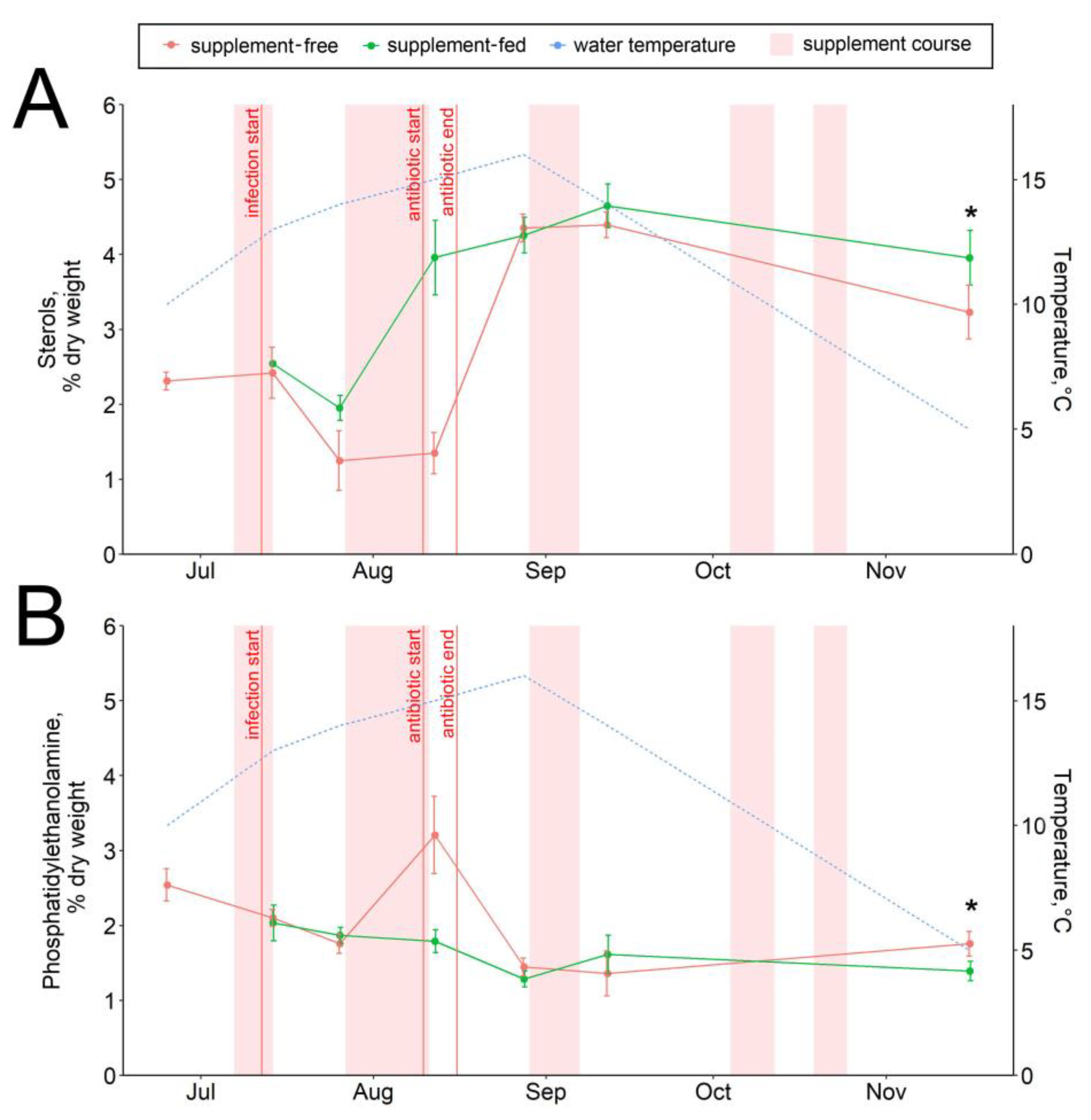
3.3. Hepatic Lipid and Fatty Acid Profile
4. Discussion
4.1. Infections and Mortality
4.2. Antioxidant Defense
4.3. Lipid and Fatty Acid Response to Stressors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action. Rome. 2020. Available online: https://doi.org/10.4060/ca9229en (accessed on 30 November 2020).
- Beers, J.M.; Jayasundara, N. Antarctic notothenioid fish: What are the future consequences of ‘losses’ and ‘gains’ acquired during long-term evolution at cold and stable temperatures? J. Exp. Biol. 2015, 218, 1834–1845. [Google Scholar] [CrossRef] [Green Version]
- Blazer, V.S. Nutrition and disease resistance in fish. Annu. Rev. Fish Dis. 1992, 2, 309–323. [Google Scholar] [CrossRef]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Pourmohammadi Fallah, H.; Yousefi, M.; Dawood, M.A.O.; Hoseinifar, S.H.; Adineh, H.; Yilmaz, S.; Paolucci, M.; Doan, H.V. The Gene Regulatory Roles of Herbal Extracts on the Growth, Immune System, and Reproduction of Fish. Animals 2021, 11, 2167. [Google Scholar] [CrossRef]
- Nik Mohamad Nek Rahimi, N.; Natrah, I.; Loh, J.-Y.; Ervin Ranzil, F.K.; Gina, M.; Lim, S.-H.E.; Lai, K.-S.; Chong, C.-M. Phytocompounds as an Alternative Antimicrobial Approach in Aquaculture. Antibiotics 2022, 11, 469. [Google Scholar] [CrossRef]
- Skaper, S.D.; Fabris, M.; Ferrari, V.; Dalle Carbonare, M.; Leon, A. Quercetin protects cutaneous tissue-associated cell types including sensory neurons from oxidative stress induced by glutathione depletion: Cooperative effects of ascorbic acid. Free Radic. Biol. Med. 1997, 22, 669–678. [Google Scholar] [CrossRef]
- Teselkin, Y.O.; Babenkova, I.V.; Kolhir, V.K.; Baginskaya, A.L.; Tjukavkina, N.A.; Kolesnik, Y.A.; Selivanova, I.A.; Eichholz, A.A. Dihydroquercetin as a means of antioxidative defense in rats with tetrachloromethane hepatitis. Phytother. Res. 2000, 14, 160–162. [Google Scholar] [CrossRef]
- Weidmann, A.E. Dihydroquercetin: More than just an impurity? Eur. J. Pharmacol. 2012, 684, 9–26. [Google Scholar] [CrossRef]
- Hou, J.; Hu, M.; Zhang, L.; Gao, Y.; Ma, L.; Xu, Q. Dietary taxifolin protects against dextran sulfate sodium-induced colitis via NF-kB signaling, enhancing intestinal barrierand modulating gut microbiota. Front. Immunol. 2021, 11, 631809. [Google Scholar] [CrossRef]
- Awad, E.; Awaad, A.S.; Esteban, M.A. Effects of dihydroquercetin obtained from deodar (Cedrus deodara) on immune status of gilthead sea bream (Sparus aurata L.). Fish Shellfish Immunol. 2015, 43, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Manigandan, K.; Jayaraj, R.L.; Jagatheesh, K.; Elangovan, N. Taxifolin mitigates oxidative DNA damage in vitro and protects zebra fish (Danio rerio) embryos against cadmium toxicity. Environ. Toxicol. Pharmacol. 2015, 39, 1252–1261. [Google Scholar] [CrossRef]
- Pirgozliev, V.; Westbrook, C.; Woods, S.; Karagecili, M.R.; Karadas, F.; Rose, S.P.; Mansbridge, S.C. Feeding dihydroquercetin to broiler chickens. Br. Poult. Sci. 2019, 60, 241–245. [Google Scholar] [CrossRef]
- Nantz, M.; Painter, A.; Parker, E.; McGill, C.; Percival, S. Evaluation of arabinogalactan’s effect on human immunity. FASEB J. 2001, 15, 633. [Google Scholar]
- Grieshop, C.M.; Flickinger, E.A.; Fahey, G.C., Jr. Oral administration of arabinogalactan affects immune status and fecal microbial populations in dogs. Nutr. J. 2002, 132, 478–482. [Google Scholar] [CrossRef] [Green Version]
- Riede, L.; Grube, B.; Gruenwald, J. Larch arabinogalactan effects on reducing incidence of upper respiratory infections. Curr. Med. Res. Opin. 2013, 29, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Dion, C.; Chappuis, E.; Ripoll, C. Does larch arabinogalactan enhance immune function? A review of mechanistic and clinical trials. Nutr. Metab. 2016, 13, 28. [Google Scholar] [CrossRef] [Green Version]
- Polyak, S.J.; Morishima, C.; Lohmann, V.; Pal, S.; Lee, D.Y.W.; Liu, Y.; Graf, T.N.; Oberlies, N.H. Identification of hepatoprotective flavonolignans from silymarin. Proc. Natl. Acad. Sci. USA 2010, 107, 5995–5999. [Google Scholar] [CrossRef] [Green Version]
- Wagoner, J.; Negash, A.; Kane, O.J.; Martinez, L.E.; Nahmias, Y.; Bourne, N.; Owen, D.M.; Grove, J.; Brimacombe, C.; McKeating, J.A.; et al. Multiple effects of silymarin on the hepatitis C virus lifecycle. J. Hepatol. 2010, 51, 1912–1921. [Google Scholar] [CrossRef] [Green Version]
- Abid, N.; Hamad, E.; Ibrahim, M.; Abid, H.A. Antibacterial and antibiofilm activities of taxifolin against vancomycin-resistant S. aureus (VRSA). Baghdad J. Biochem. Appl. Biol. Sci. 2022, 3, 262–272. [Google Scholar] [CrossRef]
- Hauer, J.; Anderer, F.A. Mechanism of stimulation of human natural killer cytotoxicity by arabinogalactan from Larix occidentalis. Cancer Immunol. Immunother. 1993, 36, 237–244. [Google Scholar] [CrossRef]
- Choi, E.M.; Kim, A.J.; Kim, Y.O.; Hwang, J.K. Immunomodulating activity of arabinogalactan and fucoidan in vitro. J. Med. Food 2005, 8, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Udani, J.K. Immunomodulatory effects of ResistAid: A randomized, double-blind, placebo-controlled, multidose study. J. Am. Coll. Nutr. 2013, 32, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Udani, J.K.; Singh, B.B.; Barrett, M.L.; Singh, V.J. Proprietary arabinogalactan extract increases antibody response to the pneumonia vaccine: A randomized, double-blind, placebo-controlled, pilot study in healthy volunteers. Nutr. J. 2010, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Awad, E.; Austin, D.; Lyndon, A.R. Effect of black cumin seed oil (Nigella sativa) and nettle extract (Quercetin) on enhancement of immunity in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquaculture 2013, 388, 193–197. [Google Scholar] [CrossRef]
- Churova, M.V.; Lysenko, L.A.; Kantserova, N.P.; Sukhovskaya, I.V.; Rodin, M.A.; Krupnova, M.Y.; Nemova, N.N. Activity of metabolic enzymes in farmed rainbow trout Oncorhynchus mykiss Walb. affected by bacterial septicemia: The effect of food additives. Dokl. Biol. Sci. 2019, 489, 178–180. [Google Scholar] [CrossRef]
- Kantserova, N.; Lysenko, L.; Churova, M.; Tushina, E.; Sukhovskaya, I.; Nemova, N. Dietary supplement with dihydroquercetin and arabinogalactan affects growth performance, intracellular protease activities and muscle-specific gene expression in bacterially infected Oncorhynchus mykiss. Int. Aquat. Res. 2020, 12, 63–73. [Google Scholar] [CrossRef]
- Kantserova, N.P.; Tushina, E.D.; Sukhovskaya, I.V.; Lysenko, L.A. Oxidative modification and proteolysis of hepatic proteins in reared rainbow trout affected by environmental factors. Russ. J. Bioorganic Chem. 2022, 48, 609–616. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Beers, R.F., Jr.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar] [CrossRef]
- Cohn, V.H.; Lyle, J. A fluorometric assay for glutathione. Anal. Biochem. 1966, 14, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Hissin, P.J.; Hilf, R. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Noble, J.E.; Bailey, M.J.A. Quantitation of protein. Meth. Enzymol. 2009, 463, 73–95. [Google Scholar] [CrossRef]
- Skurikhin, V.N.; Dvinskaya, L.M. Determination of α-tocopherol and retinol in the blood plasma of agricultural animals by microcolumn HPLC method. Selskokhoz. Biol. 1989, 4, 127–129. (In Russian) [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Sidorov, V.S.; Lizenko, E.I.; Bolgova, O.M.; Nefedova, Z.A. Fish lipids. Analysis technique. In Salmons (Salmonidae) of Karelia; Potapova, O.I., Smirnov, Y.A., Eds.; Karelian Branch of the USSR Academy of Science: Petrozavodsk, Russia, 1972; pp. 150–162. [Google Scholar]
- Engelbrecht, F.M.; Mari, F.; Anderson, J.T. Cholesterol determination in serum: A rapid direction method. S. Afr. Med. J. 1974, 48, 250–256. [Google Scholar]
- Arduini, A.; Peschechera, A.; Dottori, S.; Sciarroni, A.F.; Serafini, F.; Calvani, M. High performance liquid chromatography of long-chain acylcarnitine and phospholipids in fatty acid turnover studies. J. Lipid Res. 1996, 37, 684–689. [Google Scholar] [CrossRef]
- R Core Team. R Foundation for Statistical Computing. R Development Core Team R: A Language and Environment for Statistical Computing. Vienna, Austria, 2019. Available online: http://www.r-project.org (accessed on 10 January 2020).
- Tiamiyu, A.M.; Olatoye, I.O.; Olayemi, O.A.; Ekundayo, T.C.; Adedeji, O.B.; Okocha, R.C. Medicinal Plant Feed Additives Enhanced Survivability and Growth Performance of Clarias gariepinus (African Catfish) against Bacterial Infection. Microbiol. Res. 2021, 12, 54. [Google Scholar] [CrossRef]
- Immanuel, G.; Uma, R.P.; Iyapparaj, P.; Citarasu, T.; Punitha Peter, S.M.; Michael Babu, M.; Palavesam, A. Dietary medicinal plant extracts improve growth, immune activity and survival of tilapia Oreochromis mossambicus. J. Fish Biol. 2009, 74, 1462–1475. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Tu, H.T.; Silvestre, F.; Bernard, A.; Douny, C.; Phuong, N.T.; Tao, C.T.; Kestemont, P. Oxidative stress response of black tiger shrimp (Penaeus monodon) to enrofloxacin and to culture system. Aquaculture 2008, 285, 244–248. [Google Scholar] [CrossRef]
- Jiang, W.D.; Liu, Y.; Jiang, J.; Wu, P.; Feng, L.; Zhou, X.Q. Copper exposure induces toxicity to the antioxidant system via the destruction of Nrf2/ARE signaling and caspase-3-regulated DNA damage in fish muscle: Amelioration by myo-inositol. Aquat. Toxicol. 2015, 159, 245–255. [Google Scholar] [CrossRef]
- Sukhovskaya, I.V.; Kantserova, N.P.; Lysenko, L.A.; Morozov, A.A. Taxifolin modulates transcriptomic response to heat stress in rainbow trout, Oncorhynchus mykiss. Animals 2022, 12, 1321. [Google Scholar] [CrossRef]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef]
- Mut-Salud, N.; Álvarez, P.J.; Garrido, J.M.; Carrasco, E.; Aránega, A.; Rodríguez-Serrano, F. Antioxidant intake and antitumor therapy: Toward nutritional recommendations for optimal results. Oxid. Med. Cell. Longev. 2016, 2016, 6719534. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Liu, Y.; Hyde, A.S.; Simpson, M.A.; Barycki, J.J. Emerging regulatory paradigms in glutathione metabolism. Adv. Cancer Res. 2014, 122, 69–101. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Cha, K.H.; Selenge, D.; Solongo, A.; Nho, C.W. The chemopreventive effect of taxifolin is exerted through ARE-dependent gene regulation. Biol. Pharm. Bull. 2007, 30, 1074–1079. [Google Scholar] [CrossRef] [Green Version]
- Fodor, E.; Jones, R.H.; Buda, C.; Kitajka, K.; Dey, I.; Farkas, T. Molecular architecture and biophysical properties of phospholipids during thermal adaptation in fish: An experimental and model study. Lipids 1995, 30, 1119–1126. [Google Scholar] [CrossRef]
- Logue, J.A.; de Vries, A.L.; Fodor, E.; Cossins, A.R. Lipid compositional correlates of temperature-adaptive interspecific differences in membrane physical structure. J. Exp. Biol. 2000, 203, 2105–2115. [Google Scholar] [CrossRef]
- Vance, D.E. Biochemistry of Lipids. Lipoproteins and Membranes, 4th ed.; Vance, J.E., Ed.; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Martinez-Rubio, L.; Morais, S.; Evensen, Ø.; Wadsworth, S.; Vecino, J.G.; Ruohonen, K.; Bell, J.G.; Tocher, D.R. Effect of functional feeds on fatty acid and eicosanoid metabolism in liver and head kidney of Atlantic salmon (Salmo salar L.) with experimentally induced heart and skeletal muscle inflammation. Fish Shellfish Immunol. 2013, 34, 1533–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish, C.C. Essential fatty acids in aquatic food webs. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 309–326. [Google Scholar] [CrossRef]
- Rodríguez, C.; Acosta, C.; Badía, P.; Cejas, J.R.; Santamaría, F.J.; Lorenzo, A. Assessment of lipid and essential fatty acids requirements of black seabream (Spondyliosoma cantharus) by comparison of lipid composition in muscle and liver of wild and captive adult fish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Görgün, S.; Akpinar, M.A. Liver and muscle fatty acid composition of mature and immature rainbow trout (Oncorhynchus mykiss) fed two different diets. Biologia 2007, 62, 351–355. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Dessen, J.E.; Weihe, R.; Hatlen, B.; Thomassen, M.S.; Rørvik, K.A. Different growth performance, lipid deposition, and nutrient utilization in in-season (S1) Atlantic salmon post-smolt fed isoenergetic diets differing in protein-to-lipid ratio. Aquaculture 2017, 473, 345–354. [Google Scholar] [CrossRef]
- Parshukov, A.N.; Fokina, N.N.; Sukhovskaya, I.V.; Kantserova, N.P.; Lysenko, L.A. Infection and antibiotic treatment have prolonged effect on gut microbiota, muscle and hepatic fatty acids in rainbow trout (Oncorhynchus mykiss). J. Appl. Microbiol. 2022, 133, 1709–1724. [Google Scholar] [CrossRef]
- Crockett, E.L. Cholesterol function in plasma membranes from ectotherms: Membrane-specific roles in adaptation to temperature. Am. Zool. 1998, 38, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Franschini, F.; Dermatini, G.; Esposti, D. Pharmacology of silymarin. Clin. Drug Investig. 2002, 22, 51–56. [Google Scholar] [CrossRef]
- Blumenthal, M.; Busse, W.R.; Goldberg, A.; Gruenwald, J.; Hall, T.; Riggins, C.W.; Rister, R.S. The Complete German Commission E Monographs: Therapeutic Guide to Herbal Medicines; Blumenthal, M., Busse, W.R., Council, A.B., Rister, R., Hall, T., Riggins, C., Eds.; American Botanical Council: Austin, TX, USA, 1998. [Google Scholar]
- Casaaschi, A.; Rubio, B.K.; Maiyoh, G.K.; Theriault, A.G. Inhibitory activity of diacylglycerol acyltransferase (DGAT) and microsomal triglyceride transfer protein (MTP) by the flavonoid, taxifolin, in HepG2 cells: Potential role in the regulation of apolipoprotein B secretion. Atherosclerosis 2004, 176, 247–253. [Google Scholar] [CrossRef]
- Gebhardt, R. Variable influence of kaempferol and myricetin on in vitro hepatocellular cholesterol biosynthesis. Planta Med. 2003, 69, 1071–1074. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukhovskaya, I.V.; Lysenko, L.A.; Fokina, N.N.; Kantserova, N.P.; Borvinskaya, E.V. Survival, Growth Performance, and Hepatic Antioxidant and Lipid Profiles in Infected Rainbow Trout (Oncorhynchus mykiss) Fed a Diet Supplemented with Dihydroquercetin and Arabinogalactan. Animals 2023, 13, 1345. https://doi.org/10.3390/ani13081345
Sukhovskaya IV, Lysenko LA, Fokina NN, Kantserova NP, Borvinskaya EV. Survival, Growth Performance, and Hepatic Antioxidant and Lipid Profiles in Infected Rainbow Trout (Oncorhynchus mykiss) Fed a Diet Supplemented with Dihydroquercetin and Arabinogalactan. Animals. 2023; 13(8):1345. https://doi.org/10.3390/ani13081345
Chicago/Turabian StyleSukhovskaya, Irina V., Liudmila A. Lysenko, Natalia N. Fokina, Nadezhda P. Kantserova, and Ekaterina V. Borvinskaya. 2023. "Survival, Growth Performance, and Hepatic Antioxidant and Lipid Profiles in Infected Rainbow Trout (Oncorhynchus mykiss) Fed a Diet Supplemented with Dihydroquercetin and Arabinogalactan" Animals 13, no. 8: 1345. https://doi.org/10.3390/ani13081345