
Fish Meal Replacement and Early Mild Stress Improve Stress Responsiveness and Survival of Fish after Acute Stress
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethic, Animal and Experimental Conditions
2.2. Diet Formulation and Experimental Design
2.3. Collecting Samples and Measuring Growth Performance
2.4. Chemical Analysis of Diets and Body
2.4.1. Collection of Blood and Sample Preparation
2.4.2. Hematology Analysis
2.4.3. Blood Biochemistry, Antioxidant Enzymes Activities, Serum Enzymes and Cortisol
2.5. Non-Specific Immune Parameters
2.6. Acute Confinement Stress (AC Stress)
2.7. Statistical Analysis
3. Results
3.1. Growth Performance, Survival Rate and Body Composition
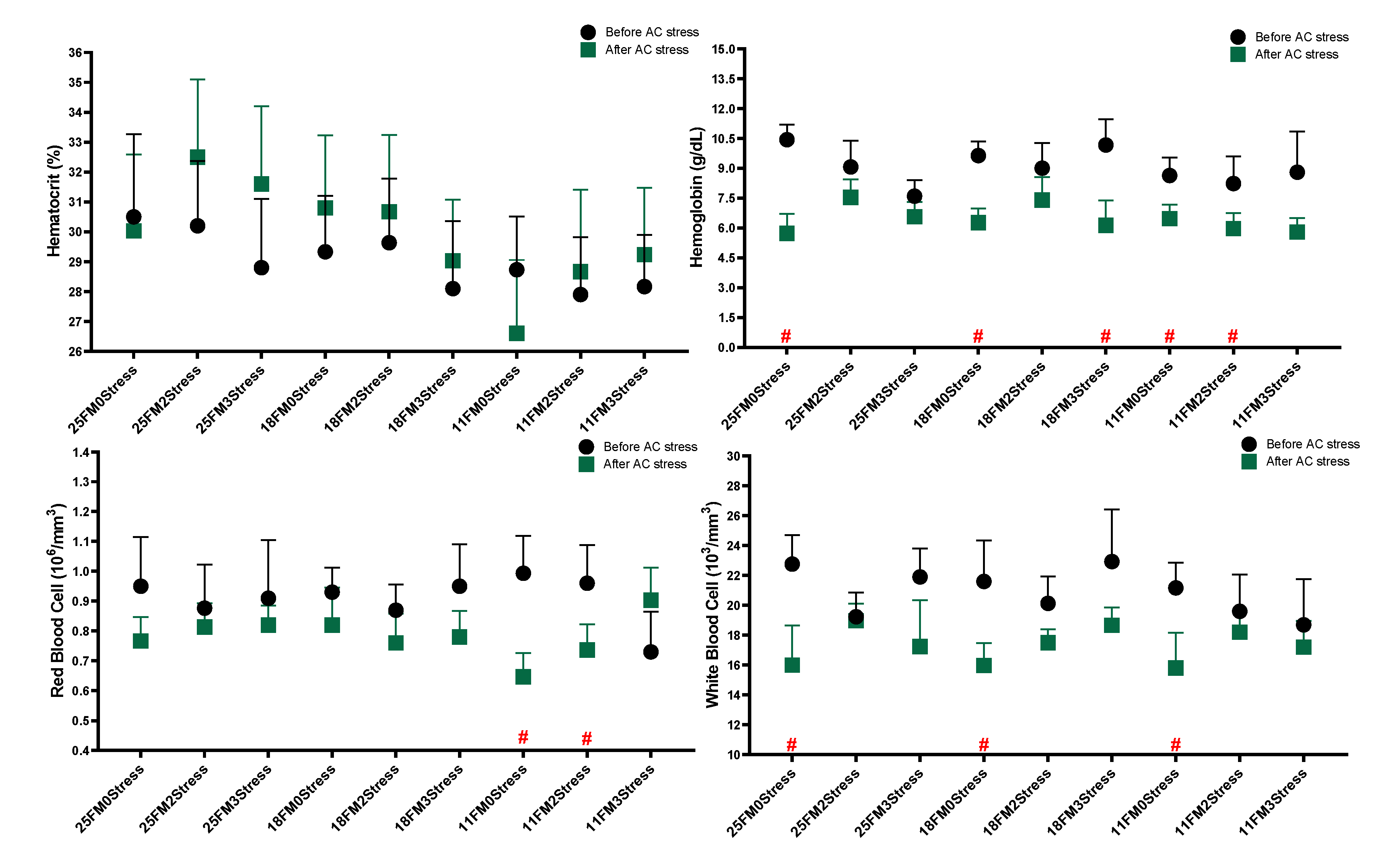
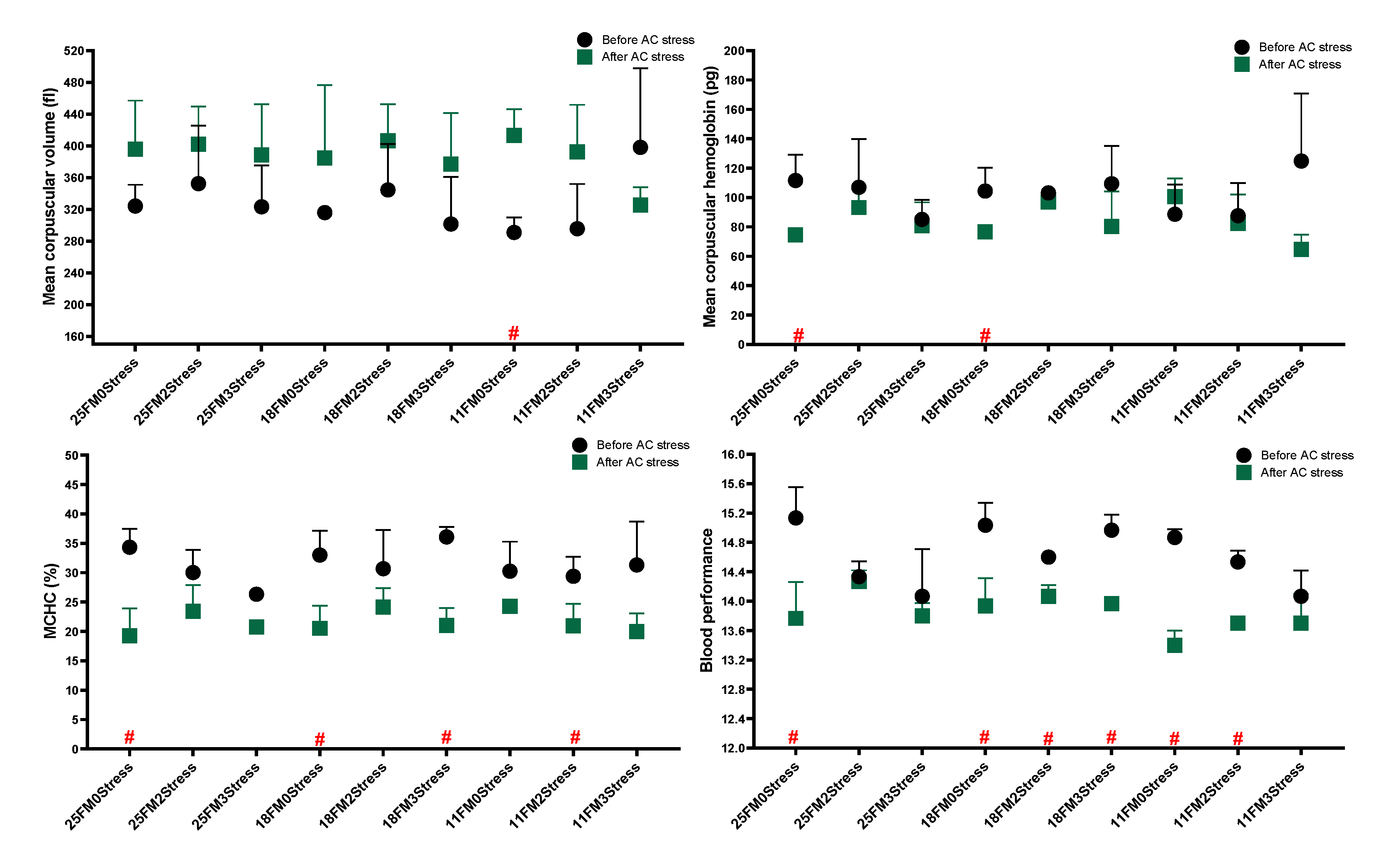
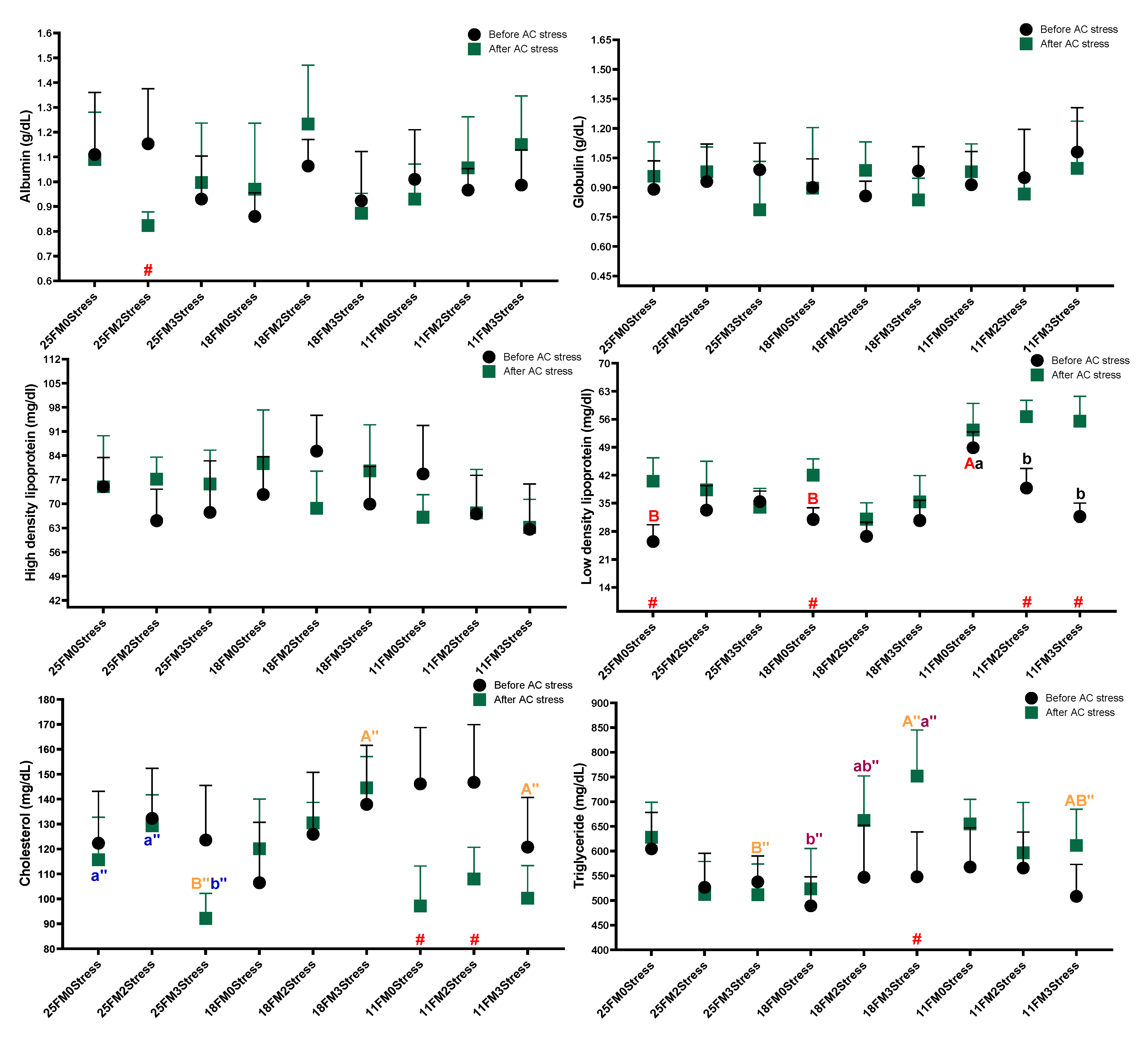
3.2. Haematology and Blood Biochemistry
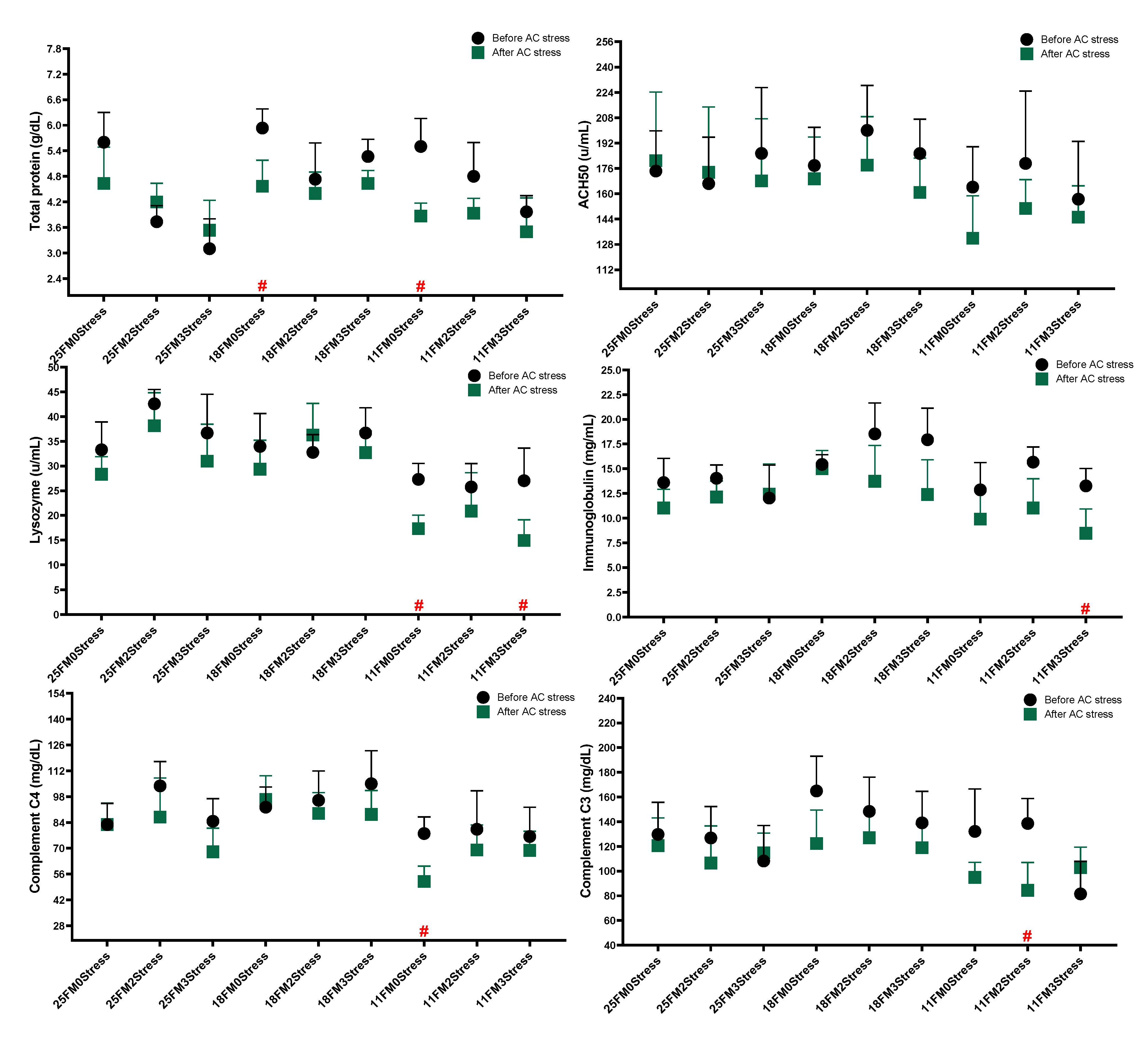
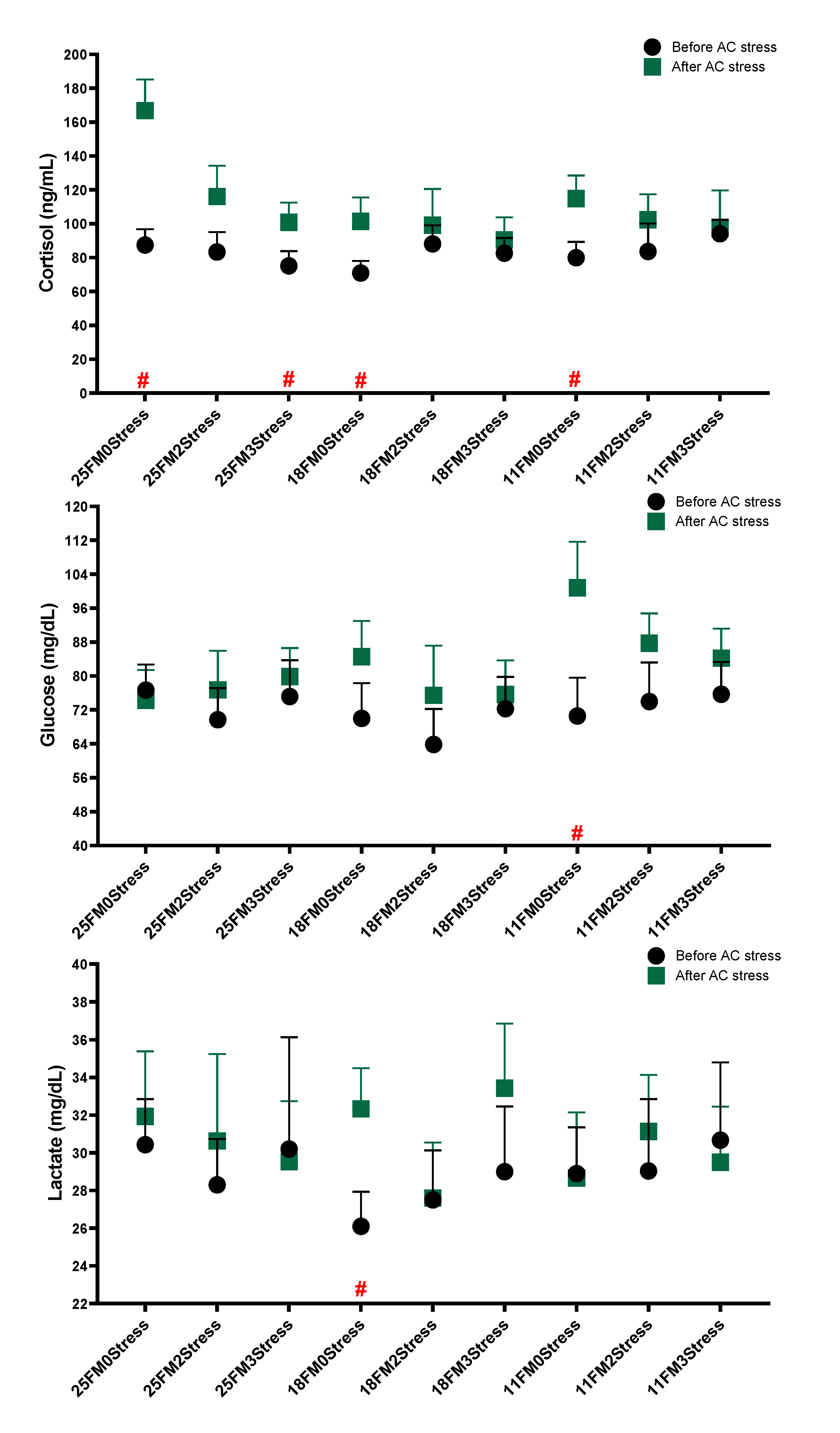
3.3. Immune and Stress Responses
3.4. Antioxidant Enzymes Activities
3.5. Liver Enzymes
4. Discussion
4.1. Growth Performance and Survival Rate
4.2. Body Composition
4.3. Haematology and Blood Biochemistry
4.4. Immune and Stress Response
4.5. Antioxidant Enzymes Activities
4.6. Liver Enzymes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosi Mobaraki, M.R.; Abedian Kenari, A.; Bahrami Gorji, S.; Esmaeili, M. Effect of dietary fish and vegetable oil on the growth performance, body composition, fatty acids profile, reproductive performance and larval resistance in pearl gourami (Trichogaster leeri). Aquac. Nutr. 2020, 26, 894–907. [Google Scholar] [CrossRef]
- Esmaeili, M.; Hosseini, H.; Zare, M.; Sobhan, R.A.; Rombenso, A. Early Mild Stress along with Lipid Improves the Stress Responsiveness of Oscar (Astronotus ocellatus). Aquac. Nutr. 2022, 2022, 8991678. [Google Scholar] [CrossRef]
- Roohani, A.M.; Abedian Kenari, A.; Fallahi Kapoorchali, M.; Borani, M.S.; Zoriezahra, S.J.; Smiley, A.H.; Esmaeili, M.; Rombenso, A.N. Effect of spirulina Spirulina platensis as a complementary ingredient to reduce dietary fish meal on the growth performance, whole-body composition, fatty acid and amino acid profiles, and pigmentation of Caspian brown trout (Salmo trutta caspius) juveniles. Aquac. Nutr. 2019, 25, 633–645. [Google Scholar]
- Zaretabar, A.; Ouraji, H.; Kenari, A.A.; Yeganeh, S.; Esmaeili, M.; Amirkolaee, A.K. One step toward aquaculture sustainability of a carnivorous species: Fish meal replacement with barley protein concentrate plus wheat gluten meal in Caspian brown trout (Salmo trutta caspius). Aquac. Rep. 2021, 20, 100714. [Google Scholar] [CrossRef]
- Hua, K. A meta-analysis of the effects of replacing fish meals with insect meals on growth performance of fish. Aquaculture 2021, 530, 735732. [Google Scholar] [CrossRef]
- Zhang, Q.; Liang, H.; Longshaw, M.; Wang, J.; Ge, X.; Zhu, J.; Li, S.; Ren, M. Effects of replacing fishmeal with methanotroph (Methylococcus capsulatus, Bath) bacteria meal (FeedKind®) on growth and intestinal health status of juvenile largemouth bass (Micropterus salmoides). Fish Shellfish Immunol. 2022, 122, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Luthada-Raswiswi, R.; Mukaratirwa, S.; O’Brien, G. Animal protein sources as a substitute for fishmeal in aquaculture diets: A systematic review and meta-analysis. Appl. Sci. 2021, 11, 3854. [Google Scholar] [CrossRef]
- Daniel, N. A review on replacing fish meal in aqua feeds using plant protein sources. Int. J. Fish. Aquat 2018, 6, 164–179. [Google Scholar]
- Hodar, A.; Vasava, R.; Mahavadiya, D.; Joshi, N. Fish meal and fish oil replacement for aqua feed formulation by using alternative sources: A review. J. Exp. Zool. India 2020, 23, 13–21. [Google Scholar]
- Shukla, A.; Munish, K.; Gyandeep, G.; Neeraj, P.; Varun, M. A review on replacing fish meal in aqua feeds using plant and animal protein sources. Int. J. Chem. Stud. 2019, 7, 4732–4739. [Google Scholar]
- Oliva-Teles, A.; Enes, P.; Couto, A.; Peres, H. Replacing fish meal and fish oil in industrial fish feeds. Feed. Feed. Pract. Aquac. 2022, 1, 231–268. [Google Scholar]
- Mugwanya, M.; Dawood, M.A.; Kimera, F.; Sewilam, H. Replacement of fish meal with fermented plant proteins in the aquafeed industry: A systematic review and meta-analysis. Rev. Aquac. 2022, 15, 62–88. [Google Scholar] [CrossRef]
- Ai, Q.; Mai, K.; Tan, B.; Xu, W.; Duan, Q.; Ma, H.; Zhang, L. Replacement of fish meal by meat and bone meal in diets for large yellow croaker, Pseudosciaena crocea. Aquaculture 2006, 260, 255–263. [Google Scholar] [CrossRef]
- Esmaeili, M.; Abedian Kenari, A.; Rombenso, A. Effects of fish meal replacement with meat and bone meal using garlic (Allium sativum) powder on growth, feeding, digestive enzymes and apparent digestibility of nutrients and fatty acids in juvenile rainbow trout (Oncorhynchus mykiss Walbaum, 1792). Aquac. Nutr. 2017, 23, 1225–1234. [Google Scholar] [CrossRef]
- Tacon, A. Utilization of conventional and unconventional protein sources in practical fish feed. A review. Nutr. Feed. Fish 1985. [Google Scholar]
- Hu, M.; Wang, Y.; Wang, Q.; Zhao, M.; Xiong, B.; Qian, X.; Zhao, Y.; Luo, Z. Replacement of fish meal by rendered animal protein ingredients with lysine and methionine supplementation to practical diets for gibel carp, Carassius auratus gibelio. Aquaculture 2008, 275, 260–265. [Google Scholar] [CrossRef]
- Madaro, A.; Olsen, R.E.; Kristiansen, T.S.; Ebbesson, L.O.; Nilsen, T.O.; Flik, G.; Gorissen, M. Stress in Atlantic salmon: Response to unpredictable chronic stress. J. Exp. Biol. 2015, 218, 2538–2550. [Google Scholar] [CrossRef] [Green Version]
- Vindas, M.A.; Madaro, A.; Fraser, T.W.; Höglund, E.; Olsen, R.E.; Øverli, Ø.; Kristiansen, T.S. Coping with a changing environment: The effects of early life stress. R. Soc. Open Sci. 2016, 3, 160382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auperin, B.; Geslin, M. Plasma cortisol response to stress in juvenile rainbow trout is influenced by their life history during early development and by egg cortisol content. Gen. Comp. Endocrinol. 2008, 158, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Zare, M.; Esmaeili, M.; Hosseini, H.; Akhavan, S.; Rombenso, A. How do optimum dietary protein and early mild stress events prepare fish for a stressful future? Stress responsiveness of oscar (Astronotus ocellatus). Aquac. Nutr. 2022, 2022, 8991678. [Google Scholar]
- Hosseinpour Aghaei, R.; Abedian Kenari, A.; Yazdani Sadati, M.A.; Esmaeili, M. The effect of time-dependent protein restriction on growth factors, nonspecific immunity, body composition, fatty acids and amino acids in the Siberian sturgeon (Acipenser baerii). Aquac. Res. 2018, 49, 3033–3044. [Google Scholar] [CrossRef]
- Esmaeili, M.; Abedian Kenari, A.; Rombenso, A. Immunohematological status under acute ammonia stress of juvenile rainbow trout (Oncorhynchus mykiss Walbaum, 1792) fed garlic (Allium sativum) powder-supplemented meat and bone meal-based feeds. Comp Clin Path. 2017, 26, 853–866. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the AOAC International; The Association: Mission Viejo, CA, USA, 2000; Volume 18. [Google Scholar]
- Aksnes, A.; Opstvedt, J. Content of digestible energy in fish feed ingredients determined by the ingredient-substitution method. Aquaculture 1998, 161, 45–53. [Google Scholar] [CrossRef]
- NRC. National Research Council, Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Kenari, A.A.; Mahmoudi, N.; Soltani, M.; Abediankenari, S. Dietary nucleotide supplements influence the growth, haemato-immunological parameters and stress responses in endangered Caspian brown trout (Salmo trutta caspius Kessler, 1877). Aquac. Nutr. 2013, 19, 54–63. [Google Scholar] [CrossRef]
- Řehulka, J.; Minařík, B.; Řehulková, E. Red blood cell indices of rainbow trout Oncorhynchus mykiss (Walbaum) in aquaculture. Aquac. Res. 2004, 35, 529–546. [Google Scholar] [CrossRef]
- Wintrobe, M. The volume and hemoglobin content of the red blood corpuscle: Simple method of calculation, normal findings, and value of such calculations in the anemias. Am. J. Med. Sci. 1929, 177, 513–522. [Google Scholar] [CrossRef]
- Esmaeili, M. Blood Performance: A New Formula for Fish Growth and Health. Biology 2021, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Clerton, P.; Troutaud, D.; Verlhac, V.; Gabaudan, J.; Deschaux, P. Dietary vitamin E and rainbow trout (Oncorhynchus mykiss) phagocyte functions: Effect on gut and on head kidney leucocytes. Fish Shellfish Immunol. 2001, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Amar, E.C.; Kiron, V.; Satoh, S.; Okamoto, N.; Watanabe, T. Effects of dietary β-carotene on the immune response of rainbow trout Oncorhynchus mykiss. Fish Sci. 2000, 66, 1068–1075. [Google Scholar] [CrossRef]
- Hosseini, H.; Pooyanmehr, M.; Foroughi, A.; Esmaeili, M.; Ghiasi, F.; Lorestany, R. Remarkable positive effects of figwort (Scrophularia striata) on improving growth performance, and immunohematological parameters of fish. Fish Shellfish Immunol. 2022, 120, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Asgari, M.; Abedian Kenari, A.; Esmaeili, M.; Rombenso, A. Effects of hydroalcoholic extract of honeybee pollen on growth performance, flesh quality, and immune and stress response response of rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2020, 26, 1505–1519. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture. In Sustainability in Action; The Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Monica Joicy, C.; Navitha, A.P.; Sivaraj, C. Studies on effect of marine macro algae Enteromorpha intestinalis as skin color enhancer and as an alternative for fish meal in feed supplemented to ornamental fishes. Int. J. Fish. Aquat 2021, 9, 201–205. [Google Scholar]
- Tu, N.P.C.; Ha, N.N.; Linh, N.T.T.; Tri, N.N. Effect of astaxanthin and spirulina levels in black soldier fly larvae meal-based diets on growth performance and skin pigmentation in discus fish, Symphysodon sp. Aquaculture 2022, 553, 738048. [Google Scholar] [CrossRef]
- Sultana, R.; Khatoon, H.; Rahman, M.R.; Haque, M.E.; Nayma, Z.; Mukta, F.A. Potentiality of Nannochloropsis sp. as partial dietary replacement of fishmeal on growth, proximate composition, pigment and breeding performance in guppy (Poecilia reticulata). Bioresour. Technol. Rep. 2022, 18, 101112. [Google Scholar] [CrossRef]
- Nishshanka, K.M.; Radampola, K.; Bulugahapitiya, V. Effects of partial replacement of dietary fishmeal using plant-protein sources on the growth performance, coloration and liver histology of guppy fry (Poecilia reticulata) in outdoor farming conditions. J. Appl. Aquac. 2021, 34, 715–733. [Google Scholar] [CrossRef]
- Chong, A.; Hashim, R.; Ali, A.b. Assessment of soybean meal in diets for discus (Symphysodon aequifasciata HECKEL) farming through a fishmeal replacement study. Aquac. Res. 2003, 34, 913–922. [Google Scholar] [CrossRef]
- Khanzadeh, M.; Esmaeili Fereidouni, A.; Seifi Berenjestanaki, S. Effects of partial replacement of fish meal with Spirulina platensis meal in practical diets on growth, survival, body composition, and reproductive performance of three-spot gourami (Trichopodus trichopterus) (Pallas, 1770). Aquac. Int. 2016, 24, 69–84. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, S.; Gan, L.; Lin, Y.; Shao, J.; Jiang, H.; Li, M. Effects of fishmeal replacement with eight protein sources on growth performance, blood biochemistry and stress resistance in Opsariichthys bidens. Aquac. Nutr. 2021, 27, 2529–2540. [Google Scholar] [CrossRef]
- Hossain, M.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M.; Islam, M. Fishmeal replacement by soya protein concentrate with inosine monophosphate supplementation influences growth, digestibility, immunity, blood health, and stress resistance of red sea bream, Pagrus major. Fish Physiol. Biochem. 2019, 45, 613–629. [Google Scholar] [CrossRef] [PubMed]
- Bu, X.; Chen, A.; Lian, X.; Chen, F.; Zhang, Y.; Muhammad, I.; Ge, X.; Yang, Y. An evaluation of replacing fish meal with cottonseed meal in the diet of juvenile Ussuri catfish Pseudobagrus ussuriensis: Growth, antioxidant capacity, nonspecific immunity and resistance to Aeromonas hydrophila. Aquaculture 2017, 479, 829–837. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, Z.; Cai, J.; Wang, J.; Wang, G.; Zhu, Z.; Cao, F. Effects of dietary fish meal replacement by fermented moringa (Moringa oleifera Lam.) leaves on growth performance, nonspecific immunity and disease resistance against Aeromonas hydrophila in juvenile gibel carp (Carassius auratus gibelio var. CAS III). Fish Shellfish Immunol. 2020, 102, 430–439. [Google Scholar] [CrossRef]
- Shearer, K.D. Factors affecting the proximate composition of cultured fishes with emphasis on salmonids. Aquaculture 1994, 119, 63–88. [Google Scholar] [CrossRef]
- Moutinho, S.; Martínez-Llorens, S.; Tomás-Vidal, A.; Jover-Cerdá, M.; Oliva-Teles, A.; Peres, H. Meat and bone meal as partial replacement for fish meal in diets for gilthead seabream (Sparus aurata) juveniles: Growth, feed efficiency, amino acid utilisation, and economic efficiency. Aquaculture 2017, 468, 271–277. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Q.; Cao, H.; Tong, T.; Huang, G.; Li, W. Replacement of fish meal by meat and bone meal in diets for juvenile snakehead Ophiocephalus argus. Fish Sci. 2015, 81, 723–729. [Google Scholar] [CrossRef]
- Hernández, C.; González-Santos, A.; Valverde-Romero, M.; González-Rodríguez, B.; Domínguez-Jiménez, P. Partial replacement of fishmeal with meat and bone meal and tuna byproducts meal in practical diets for juvenile spotted rose snapper Lutjanus guttatus. Lat. Am. J. Aquat. Res. 2016, 44, 56–64. [Google Scholar] [CrossRef]
- Kim, J.; Cho, S.H.; Kim, T.; Hur, S.W. Substitution effect of fish meal with various sources of animal by-product meals in feed on growth, feed utilisation, body composition, haematology and non-specific immune response of olive flounder (Paralichthys olivaceus, Temminck & Schlegel, 1846). Aquac. Res. 2021, 52, 2802–2817. [Google Scholar]
- Ghanbary, K.; Firouzbakhsh, F.; Arkan, E.; Mojarrab, M. The effect of Thymbra spicata hydroalcoholic extract loaded on chitosan polymeric nanoparticles on some growth performances, hematology, immunity, and response to acute stress in rainbow trout (Oncorhynchus mykiss). Aquaculture 2022, 548, 737568. [Google Scholar] [CrossRef]
- Carson, J.A.S.; Lichtenstein, A.H.; Anderson, C.A.; Appel, L.J.; Kris-Etherton, P.M.; Meyer, K.A.; Petersen, K.; Polonsky, T.; Van Horn, L. Dietary cholesterol and cardiovascular risk: A science advisory from the American Heart Association. Circulation 2020, 141, e39–e53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Liu, X.; Wang, Z.; Wang, K. Effect of partial fish meal replacement by soybean meal on the growth performance and biochemical indices of juvenile Japanese flounder Paralichthys olivaceus. Aquac. Int. 2011, 19, 143–153. [Google Scholar] [CrossRef]
- Wang, Z.; Qian, X.; Xie, S.; Yun, B. Changes of growth performance and plasma biochemical parameters of hybrid grouper (Epinephelus lanceolatus♂ × Epinephelus fuscoguttatus♀) in response to substitution of dietary fishmeal with poultry by-product meal. Aquac. Rep. 2020, 18, 100516. [Google Scholar] [CrossRef]
- Khieokhajonkhet, A.; Aeksiri, N.; Rojtinnakorn, J.; Van Doan, H.; Kaneko, G. Sacha inchi meal as a fish-meal replacer in red hybrid tilapia (Oreochromis niloticus × O. mossambicus) feeds: Effects on dietary digestibility, growth metrics, hematology, and liver and intestinal histology. Aquac. Int. 2022, 30, 677–698. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Wang, X.; Ye, J. Growth performance, plasma components, and intestinal barrier in grouper (Epinephelus coioides) are altered by dietary fish meal replacement with extruded soybean meal. Aquac. Rep. 2021, 21, 100863. [Google Scholar] [CrossRef]
- Montazeri, H.; Abedian Kenari, A.; Esmaeili, M. Soybean-based diets plus probiotics improve the profile of fatty acids, digestibility, intestinal microflora, growth performance, and the innate immunity of beluga (Huso huso). Aquac. Res. 2021, 52, 152–166. [Google Scholar] [CrossRef]
- Ravardshiri, M.; Bahram, S.; Javadian, S.R.; Bahrekazemi, M. Cinnamon Promotes Growth Performance, Digestive Enzyme, Blood Parameters, and Antioxidant Activity of Rainbow Trout (Oncorhynchus mykiss) in Low-Carbohydrate Diets. Turkish J. Fish. Aquat. Sci. 2021, 21, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A. Gut feelings: The emerging biology of gut–brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Liang, D.; Yang, Q.; Tan, B.; Dong, X.; Chi, S.; Liu, H.; Zhang, S. The effect of partial replacement of fish meal by soy protein concentrate on growth performance, immune responses, gut morphology and intestinal inflammation for juvenile hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂). Fish Shellfish Immunol. 2020, 98, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Pottinger, T. Changes in blood cortisol, glucose and lactate in carp retained in anglers’ keepnets. J. Fish Biol. 1998, 53, 728–742. [Google Scholar] [CrossRef]
- Busti, S.; Bonaldo, A.; Dondi, F.; Cavallini, D.; Yúfera, M.; Gilannejad, N.; Moyano, F.J.; Gatta, P.P.; Parma, L. Effects of different feeding frequencies on growth, feed utilisation, digestive enzyme activities and plasma biochemistry of gilthead sea bream (Sparus aurata) fed with different fishmeal and fish oil dietary levels. Aquaculture 2020, 529, 735616. [Google Scholar] [CrossRef]
- Kader, M.A.; Bulbul, M.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Nguyen, B.T.; Komilus, C.F. Effect of complete replacement of fishmeal by dehulled soybean meal with crude attractants supplementation in diets for red sea bream, Pagrus major. Aquaculture 2012, 350, 109–116. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative stress and antioxidant defense in fish: The implications of probiotic, prebiotic, and synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 198–217. [Google Scholar] [CrossRef]
- Song, S.G.; Chi, S.Y.; Tan, B.P.; Liang, G.L.; Lu, B.Q.; Dong, X.H.; Yang, Q.H.; Liu, H.Y.; Zhang, S. Effects of fishmeal replacement by Tenebrio molitor meal on growth performance, antioxidant enzyme activities and disease resistance of the juvenile pearl gentian grouper (Epinephelus lanceolatus♂ × Epinephelus fuscoguttatus♀). Aquac. Res. 2018, 49, 2210–2217. [Google Scholar] [CrossRef]
- Wang, Y.; Tao, S.; Liao, Y.; Lian, X.; Luo, C.; Zhang, Y.; Yang, C.; Cui, C.; Yang, J.; Yang, Y. Partial fishmeal replacement by mussel meal or meat and bone meal in low-fishmeal diets for juvenile Ussuri catfish (Pseudobagrus ussuriensis): Growth, digestibility, antioxidant capacity and IGF-I gene expression. Aquac. Nutr. 2020, 26, 727–736. [Google Scholar] [CrossRef]
- Twahirwa, I.; Wu, C.; Ye, J.; Zhou, Q. The effect of dietary fish meal replacement with blood meal on growth performance, metabolic activities, antioxidant and innate immune responses of fingerlings black carp, Mylopharyngodon piceus. Aquac. Res. 2021, 52, 702–714. [Google Scholar] [CrossRef]
- Ji, H.; Zhang, J.L.; Huang, J.Q.; Cheng, X.F.; Liu, C. Effect of replacement of dietary fish meal with silkworm pupae meal on growth performance, body composition, intestinal protease activity and health status in juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Res. 2015, 46, 1209–1221. [Google Scholar] [CrossRef]
- Hu, Y.; Huang, Y.; Tang, T.; Zhong, L.; Chu, W.; Dai, Z.; Chen, K.; Hu, Y. Effect of partial black soldier fly (Hermetia illucens L.) larvae meal replacement of fish meal in practical diets on the growth, digestive enzyme and related gene expression for rice field eel (Monopterus albus). Aquac. Rep. 2020, 17, 100345. [Google Scholar] [CrossRef]
- Liao, H.; Liu, P.; Deng, Y.; Zhang, W.; Pan, C.; Jia, Y.; Long, F.; Tang, H. Feeding effects of low-level fish meal replacement by algal meals of Schizochytrium limacinum and Nannochloropsis salina on largemouth bass (Micropterus salmoides). Aquaculture 2022, 557, 738311. [Google Scholar] [CrossRef]
- Ramezanzadeh, S.; Abedian Kenari, A.; Esmaeili, M. Immunohematological parameters of rainbow trout (Oncorhynchus mykiss) fed supplemented diet with different forms of barberry root (Berberis vulgaris). Comp Clin Path. 2020, 29, 177–187. [Google Scholar] [CrossRef]
- Ramezanzadeh, s.; Abedian Kenari, A.; Esmaeili, M.; Rombenso, A. Effects of different forms of barberry root (Berberis vulgaris) on growth performance, muscle fatty acids profile, whole-body composition, and digestive enzymes of rainbow trout (Oncorhynchus mykiss). J. World Aquac. Soc. 2021, 52, 284–302. [Google Scholar] [CrossRef]
- Zeilab Sendijani, R.; Abedian Kenari, A.; Smiley, A.H.; Esmaeili, M. The effect of extract from dill (Anethum graveolens) on the growth performance, body composition, immune system and antioxidant system of Rainbow Trout (Oncorhynchus mykiss). N. Am. J. Aquac. 2020, 82, 119–131. [Google Scholar] [CrossRef]
- Zhao, Z.-X.; Song, C.-Y.; Xie, J.; Ge, X.-P.; Liu, B.; Xia, S.-L.; Yang, S.; Wang, Q.; Zhu, S.-H. Effects of fish meal replacement by soybean peptide on growth performance, digestive enzyme activities, and immune responses of yellow catfish Pelteobagrus fulvidraco. Fish Sci. 2016, 82, 665–673. [Google Scholar] [CrossRef]
- Lim, S.-J.; Kim, S.-S.; Ko, G.-Y.; Song, J.-W.; Oh, D.-H.; Kim, J.-D.; Kim, J.-U.; Lee, K.-J. Fish meal replacement by soybean meal in diets for Tiger puffer, Takifugu rubripes. Aquaculture 2011, 313, 165–170. [Google Scholar] [CrossRef]
- Chen, Y.; Sagada, G.; Xu, B.; Chao, W.; Zou, F.; Ng, W.K.; Sun, Y.; Wang, L.; Zhong, Z.; Shao, Q. Partial replacement of fishmeal with Clostridium autoethanogenum single-cell protein in the diet for juvenile black sea bream (Acanthopagrus schlegelii). Aquac. Res. 2020, 51, 1000–1011. [Google Scholar] [CrossRef]
- Yu, M.; Li, Z.; Chen, W.; Rong, T.; Wang, G.; Wang, F.; Ma, X. Evaluation of full-fat Hermetia illucens larvae meal as a fishmeal replacement for weanling piglets: Effects on the growth performance, apparent nutrient digestibility, blood parameters and gut morphology. Anim. Feed Sci. Technol. 2020, 264, 114431. [Google Scholar] [CrossRef]
- Hassaan, M.; El-Sayed, A.; Soltan, M.; Iraqi, M.; Goda, A.; Davies, S.; El-Haroun, E.; Ramadan, H. Partial dietary fish meal replacement with cotton seed meal and supplementation with exogenous protease alters growth, feed performance, hematological indices and associated gene expression markers (GH, IGF-I) for Nile tilapia, Oreochromis niloticus. Aquaculture 2019, 503, 282–292. [Google Scholar] [CrossRef]
- Mikołajczak, Z.; Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Kołodziejski, P.; Pruszyńska-Oszmałek, E.; Józefiak, D. The first insight into black soldier fly meal in brown trout nutrition as an environmentally sustainable fish meal replacement. Animal 2022, 16, 100516. [Google Scholar] [CrossRef] [PubMed]
- Madibana, M.J.; Mwanza, M.; Lewis, B.R.; Fouché, C.H.; Toefy, R.; Mlambo, V. Black soldier fly larvae meal as a fishmeal substitute in juvenile dusky kob diets: Effect on feed utilisation, growth performance, and blood parameters. Sustainability 2020, 12, 9460. [Google Scholar] [CrossRef]
- Tejpal, C.; Pal, A.; Sahu, N.; Kumar, J.A.; Muthappa, N.; Vidya, S.; Rajan, M. Dietary supplementation of L-tryptophan mitigates crowding stress and augments the growth in Cirrhinus mrigala fingerlings. Aquaculture 2009, 293, 272–277. [Google Scholar] [CrossRef]
- Liu, F.; Shi, H.-Z.; Guo, Q.-S.; Yu, Y.-B.; Wang, A.-M.; Lv, F.; Shen, W.-B. Effects of astaxanthin and emodin on the growth, stress resistance and disease resistance of yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish Immunol. 2016, 51, 125–135. [Google Scholar] [CrossRef]
- Dawood, M.A.; Gewaily, M.S.; Monier, M.N.; Younis, E.M.; Van Doan, H.; Sewilam, H. The regulatory roles of yucca extract on the growth rate, hepato-renal function, histopathological alterations, and immune-related genes in common carp exposed with acute ammonia stress. Aquaculture 2021, 534, 736287. [Google Scholar] [CrossRef]
- Sun, Z.; Tan, X.; Liu, Q.; Ye, H.; Zou, C.; Xu, M.; Zhang, Y.; Ye, C. Physiological, immune responses and liver lipid metabolism of orange-spotted grouper (Epinephelus coioides) under cold stress. Aquaculture 2019, 498, 545–555. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Gupta, S.K.; Yousefi, M.; Kulikov, E.V.; Drukovsky, S.G.; Petrov, A.K.; Mirghaed, A.T.; Hoseinifar, S.H.; Van Doan, H. Mitigation of transportation stress in common carp, Cyprinus carpio, by dietary administration of turmeric. Aquaculture 2022, 564, 737380. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | 25FM | 18FM | 11FM | |
|---|---|---|---|---|
| g/kg, as-fed basis | ||||
| Corn meal | 121.6 | 103.6 | 86.6 | |
| Blood meal | 80 | 95 | 110 | |
| Fish meal | 250 | 180 | 110 | |
| Meat and bone meal | 60 | 130 | 200 | |
| Lysine | 0 | 2 | 3 | |
| Methionine | 0 | 1 | 2 | |
| Other ingredients † | 488.4 | 488.4 | 488.4 | |
| Proximate composition (g/kg dry matter) | ||||
| Crude protein | 445.1 | 450.0 | 448.9 | |
| Crude lipid | 182.4 | 175.1 | 171.3 | |
| Ash | 65.0 | 67.5 | 67.1 | |
| Carbohydrate †† | 199.2 | 196.4 | 194.5 | |
| Moisture | 108.3 | 113.0 | 111.2 | |
| Gross energy (kJ/g) ‡‡ | 21.14 | 20.83 | 20.98 | |
| Amino acids (g/kg, as-fed basis) | Requirements # | |||
| Arginine | 23.7 | 20.8 | 24.1 | 15 |
| Histidine | 8.4 | 7.6 | 8.2 | 8 |
| Isoleucine | 17.5 | 19.3 | 18.0 | 11 |
| Leucine | 30.3 | 28.7 | 29.7 | 15 |
| Lysine | 26.4 | 26.0 | 27.8 | 24 |
| Methionine + Cysteine | 11.2 | 11.3 | 11.0 | 10 |
| Phenylalanine | 20.9 | 22.1 | 21.2 | 9 |
| Tyrosine | 13.2 | 12.9 | 13.5 | 9 |
| Threonine | 18.1 | 18.5 | 19.4 | 11 |
| Valine | 21.8 | 22.2 | 22.7 | 12 |
| 25FM, without Stress | 25FM, 2-Time Stress | 25FM, 3-Time Stress | 18FM, without Stress | 18FM, 2-Time Stress | 18FM, 3-Time Stress | 11FM, without Stress | 11FM, 2-Time Stress | 11FM, 3-Time Stress | |
|---|---|---|---|---|---|---|---|---|---|
| Week 1 | WS | WS | WS | WS | WS | WS | WS | WS | WS |
| Week 2 | WS | Stress | Stress | WS | Stress | Stress | WS | Stress | Stress |
| Week 3 | WS | WS | WS | WS | WS | WS | WS | WS | WS |
| Week 4 | WS | WS | WS | WS | WS | WS | WS | WS | WS |
| Week 5 | WS | WS | WS | WS | WS | WS | WS | WS | WS |
| Week 6 | WS | WS | Stress | WS | WS | Stress | WS | WS | Stress |
| Week 7 | WS | WS | WS | WS | WS | WS | WS | WS | WS |
| Week 8 | WS | Stress | Stress | WS | Stress | Stress | WS | Stress | Stress |
| Week 9 | WS | WS | WS | WS | WS | WS | WS | WS | WS |
| Calculation of weight of all fishes, growth, sampling of blood and other factors | |||||||||
| Week 10 | WS | WS | WS | WS | WS | WS | WS | WS | WS |
| 25FM0Stress | 25FM2Stress | 25FM3Stress | 18FM0Stress | 18FM2Stress | 18FM3Stress | 11FM0Stress | 11FM2Stress | 11FM3Stress | |
|---|---|---|---|---|---|---|---|---|---|
| Initial weight (g) | 5.20 ± 1.25 | 5.33 ± 1.11 | 5.77 ± 1.12 | 4.47 ± 0.71 | 5.40 ± 1.35 | 4.72 ± 0.44 | 5.25 ± 1.29 | 5.07 ± 0.40 | 5.57 ± 0.65 |
| Final weight (g) | 47.17 ± 7.47 | 40.57 ± 6.84 | 36.23 ± 6.57 | 43.08 ± 4.40 | 50.64 ± 9.35 | 35.47 ± 7.40 | 44.03 ± 5.78 | 41.36 ± 6.95 | 37.46 ± 6.06 |
| SGR (%/day) | 3.52 ± 0.20 | 3.23 ± 0.20 | 2.92 ± 0.25 | 3.61 ± 0.38 | 3.57 ± 0.18 | 3.18 ± 0.35 | 3.40 ± 0.56 | 3.32 ± 0.09 | 3.02 ± 0.18 |
| FCR | 2.06 ± 0.18 | 2.24 ± 0.21 | 2.28 ± 0.31 | 2.20 ± 0.37 | 2.20 ± 0.29 | 2.44 ± 0.44 | 2.18 ± 0.25 | 2.20 ± 0.27 | 2.34 ± 0.35 |
| DFI (%/day) | 5.27 ± 0.52 | 5.46 ± 0.92 | 5.25 ± 0.78 | 5.66 ± 1.02 | 5.66 ± 0.76 | 5.88 ± 1.26 | 5.41 ± 0.62 | 5.43 ± 0.65 | 5.51 ± 0.99 |
| HSI (%) | 2.30 ± 0.37 | 2.59 ± 0.52 | 2.52 ± 0.36 | 2.35 ± 0.54 | 2.32 ± 0.27 | 2.48 ± 0.16 | 2.98 ± 0.43 | 2.89 ± 0.34 | 3.04 ± 0.81 |
| VSI (%) | 6.17 ± 0.40 | 6.11 ± 0.51 | 6.20 ± 0.56 | 5.97 ± 0.32 | 6.20 ± 1.05 | 6.14 ± 0.66 | 6.30 ± 0.46 | 6.03 ± 0.57 | 6.20 ± 0.56 |
| Condition factor | 2.39 ± 0.17 | 2.06 ± 0.19 | 1.92 ± 0.12 | 2.21 ± 0.19 | 2.51 ± 0.13 | 1.89 ± 0.34 | 2.29 ± 0.57 | 2.09 ± 0.23 | 2.01 ± 0.32 |
| Survival rate (%) # | 94.44 ± 6.36 | 94.44 ± 4.81 | 91.67 ± 4.17 | 93.06 ± 2.41 | 95.83 ± 7.22 | 91.67 ± 8.33 | 93.06 ± 6.36 | 91.67 ± 0.00 | 91.67 ± 4.17 |
| Survival rate after AC (%) | 56.67 ± 5.77 | 83.33 ± 11.55 | 63.33 ± 20.82 | 56.67 ± 5.77 | 73.33 ± 20.82 | 56.67 ± 20.82 | 43.33 ± 11.55 | 53.33 ± 15.28 | 46.67 ± 5.77 |
| p-Value | Main Effects (Mean) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Diet Effect | Stress–Number Effect | Interactions | 25FM Diets | 18FM Diets | 11FM Diets | 0stress | 2stress | 3stress | |
| Final weight | 0.790 | 0.033 | 0.409 | 44.75 a | 44.19 a | 36.39 b | |||
| Weight gain | 0.666 | 0.019 | 0.429 | 39.78 a | 38.92 a | 31.03 b | |||
| SGR | 0.260 | 0.031 | 0.974 | 3.51 a | 3.37 a | 3.03 b | |||
| Survival rate after AC | 0.025 | 0.042 | 0.869 | 67.7 a | 62.2 a | 47.7 b | 52.2 b | 70.0 a | 55.5 b |
| Total protein | 0.109 | 0.001 | 0.347 | 5.68 a | 4.42 b | 3.44 c | |||
| BP | 0.253 | 0.001 | 0.633 | 15.01 a | 14.55 ab | 14.17 b | |||
| Ht-stress | 0.041 | 0.469 | 0.608 | 31.38 a | 30.07 b | 28.16 b | |||
| WBC-stress | 0.920 | 0.049 | 0.762 | 15.92 b | 18.22 a | 17.70 a | |||
| BP-stress | 0.025 | 0.127 | 0.658 | 13.93 a | 13.99 a | 13.62 b | |||
| TP-stress | 0.036 | 0.247 | 0.457 | 4.53 a | 4.12 ab | 3.77 b | |||
| Glucose-stress | 0.006 | 0.202 | 0.288 | 76.94 b | 78.53 b | 90.91 a | |||
| Cortisol-stress | 0.032 | 0.028 | 0.609 | 119.1 a | 97.2 b | 104.8 ab | 118.8 a | 105.8 b | 96.2 b |
| LDL-stress | 0.001 | 0.336 | 0.232 | 37.59 b | 36.12 b | 55.17 a | |||
| Lysozyme | 0.001 | 0.646 | 0.360 | 37.53 a | 34.48 a | 26.70 b | |||
| Immunoglobulin | 0.005 | 0.179 | 0.661 | 13.22 b | 17.30 a | 13.93 b | |||
| Complement C4 | 0.034 | 0.440 | 0.510 | 90.44 ab | 97.78 a | 78.23 b | |||
| Complement C3 | 0.036 | 0.041 | 0.608 | 121.6 b | 150.8 a | 117.4 b | 142.3 a | 137.9 a | 109.6 b |
| Lysozyme-stress | 0.001 | 0.048 | 0.837 | 32.50 a | 32.80 a | 17.72 b | 25.01 b | 31.79 a | 26.22 ab |
| Immunoglobulin-stress | 0.025 | 0.638 | 0.705 | 11.87 ab | 13.71 a | 9.80 b | |||
| C4-stress | 0.001 | 0.571 | 0.230 | 79.29 a | 91.20 a | 63.33 b | |||
| C3-stress | 0.032 | 0.770 | 0.835 | 113.9 ab | 122.8 a | 94.1 b | |||
| SOD | 0.020 | 0.218 | 0.321 | 63.1 a | 63.4 a | 51.0 b | |||
| Catalase | 0.001 | 0.090 | 0.145 | 47.48 a | 51.22 a | 29.49 b | |||
| GPX | 0.039 | 0.997 | 0.066 | 195.2 ab | 215.1 a | 179.5 b | |||
| ALP | 0.035 | 0.764 | 0.094 | 231.5 ab | 242.0 a | 199.9 b | |||
| LDH | 0.002 | 0.074 | 0.286 | 1564 a | 1190 b | 1579 a | |||
| ALP-stress | 0.198 | 0.047 | 0.496 | 199.2 b | 205.8 ab | 237.8 a | |||
| LDH-stress | 0.001 | 0.843 | 0.423 | 1540 b | 1927 a | 2017 a | |||
| AST-stress | 0.009 | 0.016 | 0.059 | 219.3 b | 218.4 b | 267.8 a | 212.7 b | 229.1 b | 263.7 a |
| 25FM0Stress | 25FM2Stress | 25FM3Stress | 18FM0Stress | 18FM2Stress | 18FM3Stress | 11FM0Stress | 11FM2Stress | 11FM3Stress | |
|---|---|---|---|---|---|---|---|---|---|
| Protein | 180.2 ± 18.5 | 186.9 ± 9.4 | 184.8 ± 6.5 | 180.0 ± 15.7 | 178.8 ± 11.8 | 181.9 ± 9.2 | 183.6 ± 14.4 | 175.1 ± 6.7 | 182.1 ± 2.3 |
| Lipid | 60.2 ± 5.8 | 63.1 ± 6.2 | 59.7 ± 7.2 | 60.2 ± 7.3 | 61.7 ± 6.4 | 61.1 ± 5.2 | 58.1 ± 4.5 | 61.6 ± 5.6 | 62.1 ± 3.9 |
| Ash | 30.6 ± 3.5 | 31.0 ± 5.5 | 30.0 ± 2.6 | 27.6 ± 3.8 | 31.6 ± 4.0 | 29.7 ± 2.1 | 30.7 ± 3.5 | 30.5 ± 2.5 | 28.7 ± 2.6 |
| Moisture | 686.7 ± 51.4 | 689.4 ± 38.5 | 672.2 ± 34.0 | 690.8 ± 55.4 | 678.6 ± 35.6 | 672.8 ± 47.8 | 683.9 ± 38.1 | 692.9 ± 44.4 | 697.8 ± 50.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zare, M.; Kazempour, M.; Hosseini, H.; Hosseini Choupani, S.M.; Akhavan, S.R.; Rombenso, A.; Esmaeili, N. Fish Meal Replacement and Early Mild Stress Improve Stress Responsiveness and Survival of Fish after Acute Stress. Animals 2023, 13, 1314. https://doi.org/10.3390/ani13081314
Zare M, Kazempour M, Hosseini H, Hosseini Choupani SM, Akhavan SR, Rombenso A, Esmaeili N. Fish Meal Replacement and Early Mild Stress Improve Stress Responsiveness and Survival of Fish after Acute Stress. Animals. 2023; 13(8):1314. https://doi.org/10.3390/ani13081314
Chicago/Turabian StyleZare, Mahyar, Mohammad Kazempour, Hossein Hosseini, Seyedeh Mahsa Hosseini Choupani, Sobhan R. Akhavan, Artur Rombenso, and Noah Esmaeili. 2023. "Fish Meal Replacement and Early Mild Stress Improve Stress Responsiveness and Survival of Fish after Acute Stress" Animals 13, no. 8: 1314. https://doi.org/10.3390/ani13081314