

Net Portal Appearance of Amino Acids in Iberian and Landrace Pigs Fed Different Protein Content in the Diet
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Experimental Schedule and Measurements
2.3. Statistical Analyses
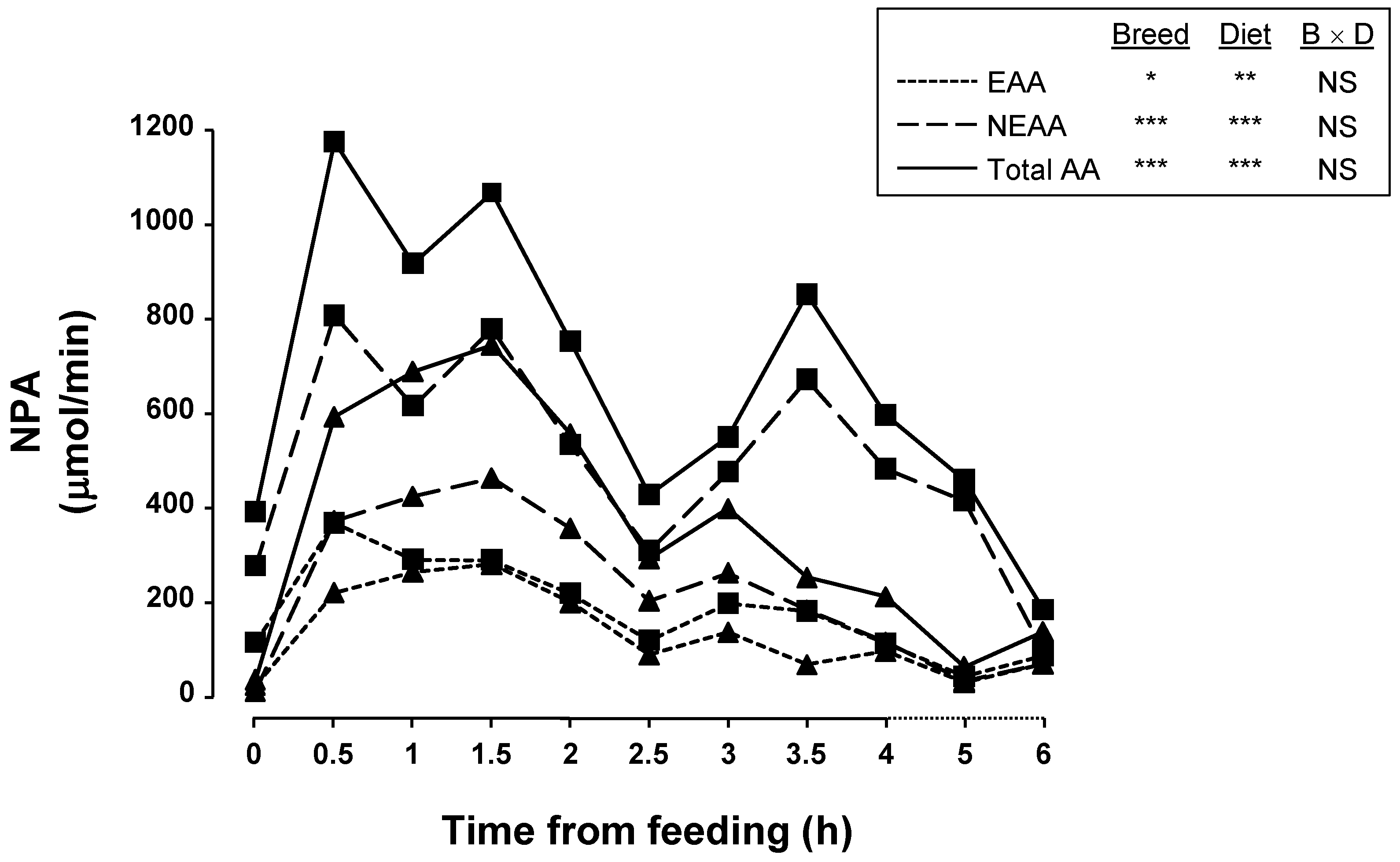
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Estévez, V.; García, A.; Peña, F.; Gómez, A.G. Foraging of Iberian fattening pigs grazing natural pasture in the dehesa. Livest. Sci. 2009, 120, 135–143. [Google Scholar] [CrossRef]
- López-Bote, C.J. Sustained utilization of the Iberian pig breed. Meat Sci. 1998, 49 (Suppl. 1), S17–S27. [Google Scholar] [CrossRef]
- Fernández-Fígares, I.; Lachica, M.; Nieto, R.; Rivera-Ferre, M.G.; Aguilera, J.F. Serum profile of metabolites and hormones in obese (Iberian) and lean (Landrace) growing gilts fed balanced or lysine deficient diets. Livest. Sci. 2007, 110, 73–81. [Google Scholar] [CrossRef]
- Nieto, R.; Lara, L.; Barea, R.; García-Valverde, R.; Aguinaga, M.A.; Conde-Aguilera, J.A.; Aguilera, J.F. Response analysis of the Iberian pig growing from birth to 150 kg body weight to changes in protein and energy supply. J. Anim. Sci. 2012, 90, 3809–3820. [Google Scholar] [CrossRef]
- Rivera-Ferre, M.G.; Aguilera, J.F.; Nieto, R. Muscle fractional protein synthesis is higher in Iberian than in Landrace growing pigs fed adequate or lysine-deficient diets. J. Nutr. 2005, 135, 469–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-López, J.M.; Lachica, M.; González-Valero, L.; Fernández-Fígares, I. Energy expenditure of splanchnic tissues in Iberian and Landrace growing gilts. Livest. Sci. 2010, 133, 61–63. [Google Scholar] [CrossRef]
- Nieto, R.; Miranda, A.; García, M.A.; Aguilera, J.F. The effect of dietary protein content and feeding level on the rate of protein deposition and energy utilization in growing Iberian pigs from 15 to 50 kg body weight. Br. J. Nutr. 2002, 88, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Ferre, M.G.; Aguilera, J.F.; Nieto, R. Differences in whole-body protein turnover between Iberian and Landrace pigs fed adequate or lysine-defi cient diets. J. Anim. Sci. 2006, 84, 3346–3355. [Google Scholar] [CrossRef] [Green Version]
- Freire, J.P.B.; Dias, R.I.M.; Cunha, L.F.; Aumaitre, A. The effect of genotype and dietary fi bre level on the caecal bacterial enzyme activity of young piglets: Digestive consequences. Anim. Feed Sci. Technol. 2003, 106, 119–130. [Google Scholar] [CrossRef]
- Nieto, R.; Rivera, M.; García, M.A.; Aguilera, J.F. Amino acid availability and energy value of acorns in the Iberian pig. Livest. Prod. Sci. 2002, 77, 227–239. [Google Scholar] [CrossRef]
- García-Valverde, R.; Nieto, R.; Lachica, M.; Aguilera, J.F. Effects of herbage ingestion on the digestion site and nitrogen balance in heavy Iberian pigs fed on an acorn-based diet. Livest. Sci. 2007, 112, 63–77. [Google Scholar] [CrossRef]
- Lachica, M.; Rodríguez-López, J.M.; González-Valero, L.; Fernández-Fígares, I. Iberian pig adaptation to acorn consumption: II. Net portal appearance of amino acids. PeerJ 2018, 6, e6137. [Google Scholar] [PubMed] [Green Version]
- British Society of Animal Science (BSAS). Nutrient Requirement Standards for Pigs; British Society of Animal Science (BSAS): Penicuik, UK, 2003. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th ed; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 17th ed; Association of Official Analytical Chemists: Arlington, VA, USA, 2000. [Google Scholar]
- Cohen, S.A.; Meys, M.; Tarvin, T.L. The Pico-Tag Method. A Manual of Advanced Techniques for Amino Acid Analysis; Millipore Corporation: Bedford, MA, USA, 1989. [Google Scholar]
- Moore, S. On the determination of cystine as cysteic acid. J. Biol. Chem. 1963, 238, 235–237. [Google Scholar] [CrossRef]
- Rodríguez-López, J.M.; Lachica, M.; González-Valero, L.; Fernández-Fígares, I. Approaches for quantifying gastrointestinal nutrient absorption and metabolism in a native and a modern pig breed. J. Agric. Sci. 2013, 151, 434–443. [Google Scholar] [CrossRef]
- Fernández-Fígares, I.; Lachica, M.; Rojas-Cano, M.; González-Casado, A. Influence of para-aminohippuric acid analysis on net portal-drained viscera flux of nutrients in pigs. Animal 2018, 12, 232–238. [Google Scholar] [CrossRef]
- Katz, M.L.; Bergman, E.N. Simultaneous measurements of hepatic and portal venous blood flow in the sheep and dog. Am. J. Physiol. 1969, 216, 946–952. [Google Scholar] [CrossRef] [Green Version]
- Zierler, K.L. Theory of the use of arteriovenous concentration differences for measuring metabolism in steady and non-steady states. J. Clin. Investig. 1961, 40, 2111–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenis, N.P.; Bikker, P.; van der Meulen, J.; van Diepen, J.T.M.; Bakker, J.G.M.; Jongbloed, A.W. Effect of dietary neutral detergent fibre on ileal digestibility and portal flux of nitrogen and amino acids and on nitrogen utilization in growing pigs. J. Anim. Sci. 1996, 74, 2687–2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Meulen, J.; Bakker, J.G.M.; Smits, B.; de Visser, H. Effect of source of starch on net portal flux of glucose, lactate, volatile fatty acids and amino acids in the pig. Br. J. Nutr. 1997, 78, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agyekum, A.K.; Kiarie, E.; Walsh, M.C.; Nyachoti, C.M. Postprandial portal fluxes of essential amino acids, volatile fatty acids, and urea-nitrogen in growing pigs fed a high-fiber diet supplemented with a multi-enzyme cocktail. J. Anim. Sci. 2016, 94, 3771–3785. [Google Scholar] [CrossRef] [PubMed]
- González-Valero, L.; Rodríguez-López, J.M.; Lachica, M.; Fernández-Fígares, I. Contribution of portal-drained viscera to heat production in Iberian gilts fed a low protein diet: Comparison to Landrace. J. Sci. Food Agric. 2016, 96, 1202–1208. [Google Scholar] [CrossRef]
- Fara, J.W. Postprandial mesenteric hyperemia. In Physiology of the Intestinal Circulation; Shepherd, A.P., Granger, D.N., Eds.; Raven Press: New York, NY, USA, 1984; pp. 99–106. [Google Scholar]
- Lomax, M.A.; Baird, G.D. Blood flow and nutrient exchange across the liver and gut of the dairy cow. Effects of lactation and fasting. Br. J. Nutr. 1983, 49, 481–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huntington, G. Relationship of portal blood flow to metabolizable energy intake of cattle. Can. J. Anim. Sci. 1984, 64, 16–17. [Google Scholar] [CrossRef]
- Rérat, A. Nutritional supply of proteins and absorption of their hydrolysis products: Consequences on metabolism. Proc. Nutr. Soc. 1993, 52, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simoes Nunes, C.; Galibois, I.; Rérat, A.; Savoie, L.; Vaugelade, P. Hepatic and portal-drained viscera balances of amino acids, insulin, glucagon and gastrin in the pig after ingestion of casein or rapeseed proteins. Reprod. Nutr. Dev. 1991, 31, 217–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, T.A.; Nguyen, H.V.; Garcia-Bravo, R.; Fiorotto, M.L.; Jackson, E.M.; Lewis, D.S.; Lee, D.R.; Reeds, P.J. Amino acid composition of human milk is not unique. J. Nutr. 1994, 124, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.; Henry, J.; Reeds, P.J.; Yu, H.; Jahoor, F.; Burrin, D.G. Catabolism dominates the first-pass intestinal metabolism of dietary essential amino acids in milk protein-fed piglets. J. Nutr. 1998, 128, 606–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Huang, R.; Li, T.; Ruan, Z.; Xie, M.; Deng, Z.; Hou, Y.; Wu, G. Amino acid metabolism in the portal-drained viscera of young pigs: Effects of dietary supplementation with chitosan and pea hull. Amino Acids 2010, 39, 1581–1587. [Google Scholar] [CrossRef]
- Wu, Z.L.; Hou, Y.; Hu, S.; Bazer, F.W.; Meininger, C.J.; McNeal, C.J.; Wu, G. Catabolism and safety of supplemental L-arginine in animals. Amino Acids 2016, 48, 1541–1552. [Google Scholar] [CrossRef] [PubMed]
- Blachier, F.; Darcy-Vrillon, B.; Sener, A.; Duke, P.H.; Malaise, W.J. Arginine metabolism in rat enterocytes. Biochim. Biophys. Acta-Mol. Cell Res. 1991, 1092, 304–310. [Google Scholar] [CrossRef]
- Reverter, M.; Lundh, T.; Gonda, H.L.; Lindberg, J.E. Portal net appearance of amino acids in growing pigs fed a barley-based diet with inclusion of three different forage meals. Br. J. Nutr. 2000, 84, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Pion, R.; Fauconneau, C.; Rérat, A. Variation de la composition en acides aminés du sang porte au cours de la digestion chex le porc (Variations in the amino acid composition of the portal blood during digestion in the pig). Ann. Biol. Anim. Biochim. Biophys. 1964, 4, 383–402. [Google Scholar] [CrossRef] [Green Version]
- Stoll, B.; Burrin, D.G. Measuring splanchnic amino acid metabolism in vivo using stable isotopic tracers. J. Anim. Sci. 2006, 84, E60–E72. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Kristensen, N.B.; Che, L.; Wu, D.; Theil, P.K. Net absorption and liver metabolism of amino acids and heat production of portal-drained viscera and liver in multiparous sows during transition and lactation. J. Anim. Sci. Biotechnol. 2020, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoll, B.; Burrin, D.G.; Henry, J.F.; Jahoor, F.; Reeds, P.J. Dietary and systemic phenylalanine utilization for mucosal and hepatic constitutive protein synthesis in pigs. Am. J. Physiol. 1999, 276, G49–G57. [Google Scholar] [CrossRef]
- Fernández-Fígares, I.; Rodríguez-López, J.M.; González-Valero, L.; Lachica, M. Iberian pig adaptation to acorn consumption: I. Net portal appearance of metabolites. PeerJ 2018, 6, e5861. [Google Scholar] [CrossRef]
- Watford, M. Glutamine metabolism and function in relation to proline synthesis and the safety of glutamine and proline supplementation. J. Nutr. 2008, 138, 2003S–2007S. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Borbolla, A.G.; Knabe, D.A. The uptake of glutamine and release of arginine, citrulline and proline by the small intestine of developing pigs. J. Nutr. 1994, 124, 2437–2444. [Google Scholar] [CrossRef] [PubMed]
- Marini, J.C.; Agarwal, U.; Didelija, I.C. Dietary citrulline supplementation is an efficient strategy to increase arginine availability. In Proceedings of the 5th EAAP International Symposium on Energy and Protein Metabolism and Nutrition, Krakow, Poland, 12–15 September 2016; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; pp. 159–160. [Google Scholar]
- Windmueller, H.G.; Spaeth, A.E. Respiratory fuels and nitrogen metabolism in vivo in small intestine of fed rats. Quantitative importance of glutamine, glutamate and aspartate. J. Biol. Chem. 1980, 225, 107–122. [Google Scholar] [CrossRef]
- Bos, C.; Stoll, B.; Fouillet, H.; Gaudichon, C.; Guan, X.; Grusak, M.A.; Reeds, P.J.; Tome, D.; Burrin, D.G. Intestinal lysine metabolism is driven by the enteral availability of dietary lysine in piglets fed a bolus meal. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1246–E1257. [Google Scholar] [CrossRef] [Green Version]
- Nyachoti, C.M.; de Lange, C.F.M.; McBride, B.W.; Schulze, H. Significance of endogenous gut nitrogen losses in the nutrition of growing pigs: A review. Can. J. Anim. Sci. 1997, 77, 149–163. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Meziere, N.; Seve, B. Whole blood and plasma amino acid transfers across the portal drained viscera and liver of the pig. Reprod. Nutr. Dev. 1999, 39, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Fígares, I.; Lara, L.; Lachica, M. Betaine increases portal appearance of amino acids and peptides in Iberian pigs. In Proceedings of the 6th EAAP International Symposium on Energy and Protein Metabolism and Nutrition, Belo Horizonte, Brazil, 9–12 September 2019; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 293–294. [Google Scholar]
- Pearce, S.C.; Mani, V.; Weber, T.E.; Rhoads, R.P.; Patience, J.F.; Baumgard, L.H.; Gabler, N.K. Heat stress and reduced plane of nutrition decreases intestinal integrity and function in pigs. J. Anim. Sci. 2013, 91, 5183–5193. [Google Scholar] [CrossRef] [PubMed]
- Barea, R.; Nieto, R.; Vitari, F.; Domeneghini, C.; Aguilera, J.F. Effects of pig genotype (Iberian v. Landrace × Large White) on nutrient digestibility, relative organ weight and small intestine structure at two stages of growth. Animal 2011, 5, 547–557. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| LCP | HCP | |
|---|---|---|
| Barley grain | 866 | 802 |
| Soybean meal | 96.2 | 125 |
| Fish meal | 4 | 40 |
| Calcium carbonate | 8 | 8 |
| Calcium acid phosphate | 12 | 12 |
| Sodium chloride | 5 | 5 |
| L-Lys | 4.32 | 3.92 |
| Thr | 1.05 | 0.96 |
| Met | 0.47 | 0.21 |
| Vitamin-mineral premix 1 | 3 | 3 |
| Chemical analysis | ||
| DM, g/kg | 870.12 | 875.14 |
| Ash, g/kg | 58.41 | 59.96 |
| CP, g/kg | 145.43 | 187.12 |
| Gross energy, MJ/kg | 17.83 | 18.08 |
| Amino acids | ||
| EAA 2 | 58.0 | 73.7 |
| Arg | 9.86 | 13.3 |
| His | 3.93 | 5.01 |
| Ile | 4.38 | 5.74 |
| Leu | 8.31 | 11.0 |
| Lys | 11.5 | 13.6 |
| Met | 2.51 | 3.47 |
| Phe | 5.52 | 7.04 |
| Thr | 6.51 | 7.50 |
| Val | 5.52 | 7.10 |
| NEAA 2 | 60.1 | 77.2 |
| Ala | 4.95 | 7.12 |
| Asp | 7.78 | 11.4 |
| Cys | 2.01 | 2.51 |
| Glu | 21.9 | 26.6 |
| Gly | 5.03 | 7.00 |
| Pro | 9.13 | 10.7 |
| Ser | 5.00 | 6.55 |
| Tyr | 4.27 | 5.22 |
| Total AA 2 | 118.1 | 150.8 |
| Breed | Diet | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Iberian | Landrace | LCP | HCP | SEM 1 | Breed | Diet | Breed × Diet | |
| EAA | 146.8 | 186.3 | 132.5 | 200.8 | 10.71 | 0.0388 | 0.0015 | 0.1244 |
| Arg | 25.7 | 30.4 | 20.2 | 35.9 | 3.03 | 0.4469 | 0.0119 | 0.5154 |
| His | 9.06 | 11.8 | 8.82 | 12.1 | 0.584 | 0.0203 | 0.0067 | 0.3054 |
| Ile | 13.6 | 12.0 | 12.3 | 13.3 | 1.03 | 0.4525 | 0.6259 | <0.0001 |
| Leu | 23.7 | 37.4 | 25.1 | 36.0 | 1.60 | <0.0001 | 0.0009 | 0.1877 |
| Lys | 36.1 | 40.5 | 29.6 | 46.9 | 2.48 | 0.3850 | 0.0008 | 0.3674 |
| Met | 7.53 | 7.96 | 6.55 | 8.94 | 0.449 | 0.6419 | 0.0094 | 0.0001 |
| Phe | 15.3 | 21.5 | 16.0 | 20.9 | 0.82 | 0.0003 | 0.0039 | 0.0401 |
| Trp 2 | 1.65 | 5.07 | 2.03 | 4.69 | 0.491 | 0.0008 | 0.0084 | 0.8918 |
| Val | 14.2 | 19.7 | 11.9 | 22.1 | 2.14 | 0.2050 | 0.0200 | 0.6487 |
| NEAA | 275.3 | 519.3 | 309.9 | 484.7 | 24.37 | <0.0001 | 0.0009 | 0.2948 |
| Ala | 93.9 | 161.1 | 105.1 | 149.9 | 5.88 | <0.0001 | 0.0002 | 0.5512 |
| Asn 2 | 43.1 | 65.3 | 47.0 | 61.3 | 2.72 | 0.0001 | 0.0100 | 0.4796 |
| Asp | 5.60 | 7.40 | 5.79 | 7.20 | 0.606 | 0.1452 | 0.2528 | 0.4883 |
| Cit 2 | 13.7 | 11.0 | 12.9 | 11.9 | 0.90 | 0.1343 | 0.5803 | 0.2916 |
| Cys | 3.13 | 15.1 | 8.92 | 9.34 | 1.973 | 0.0031 | 0.9178 | 0.0289 |
| Gln 2 | −18.6 | −20.1 | −22.4 | −16.3 | 0.43 | 0.8666 | 0.5078 | 0.8226 |
| Glu | −29.2 | −7.29 | −26.3 | −10.2 | 4.284 | 0.0149 | 0.0738 | 0.8166 |
| Gly | 73.8 | 144.0 | 80.4 | 137.4 | 6.25 | <0.0001 | <0.0001 | 0.0695 |
| Hyp 2 | 1.47 | 6.83 | 3.55 | 4.75 | 0.461 | <0.0001 | 0.2011 | 0.0299 |
| Orn 2 | 11.6 | 7.21 | 8.15 | 10.7 | 0.908 | 0.0180 | 0.1706 | 0.3447 |
| Pro | 46.9 | 77.4 | 55.9 | 68.3 | 3.72 | 0.0001 | 0.1015 | 0.1528 |
| Ser | 18.2 | 30.6 | 18.7 | 30.1 | 2.00 | 0.0026 | 0.0055 | 0.1236 |
| Tau 2 | 1.79 | 5.63 | 1.84 | 5.58 | 0.651 | 0.0041 | 0.0052 | 0.6649 |
| Tyr | 9.87 | 15.1 | 10.3 | 14.7 | 0.739 | 0.0006 | 0.0042 | 0.6566 |
| TAA | 422.1 | 705.6 | 442.4 | 685.5 | 32.21 | <0.0001 | 0.0008 | 0.1326 |
| AA Intake | Fractional Absorption | |||||||
|---|---|---|---|---|---|---|---|---|
| LCP | HCP | Ib-LCP | Ib-HCP | Ld-LCP | Ld-HCP | SEM | p-Value | |
| EAA 1 | 76.7 | 99.4 | 0.449 a | 0.705 b | 0.774 b | 0.715 b | 0.0426 | 0.0108 |
| Arg | 12.3 | 16.7 | 0.461 | 0.764 | 0.716 | 0.779 | 0.0842 | 0.3553 |
| His | 5.52 | 7.07 | 0.445 a | 0.575 ab | 0.705 b | 0.652 b | 0.0338 | 0.0283 |
| Ile | 7.27 | 9.59 | 0.400 a | 0.717 b | 0.817 b | 0.282 a | 0.0465 | <0.0001 |
| Leu | 13.8 | 18.4 | 0.420 a | 0.613 ab | 0.890 c | 0.798 bc | 0.0357 | <0.0001 |
| Lys | 17.1 | 20.4 | 0.529 a | 0.830 b | 0.716 ab | 0.827 b | 0.0472 | 0.0778 |
| Met | 3.66 | 5.10 | 0.440 a | 0.749 b | 0.847 b | 0.515 a | 0.0375 | 0.0002 |
| Phe | 7.28 | 9.33 | 0.549 a | 0.752 b | 1.029 c | 0.856 bc | 0.0349 | <0.0001 |
| Val | 10.3 | 13.3 | 0.286 | 0.550 | 0.547 | 0.645 | 0.0678 | 0.2694 |
| NEAA 1 | 108.3 | 139.7 | 0.419 a | 0.819 b | 1.301 c | 1.277 c | 0.0477 | <0.0001 |
| Ala | 12.1 | 17.5 | 2.232 a | 2.318 a | 4.020 b | 3.848 b | 0.1385 | <0.0001 |
| Asp | 12.7 | 18.8 | 0.151 | 0.113 | 0.177 | 0.164 | 0.0147 | 0.4503 |
| Cys | 3.61 | 4.54 | −0.147 a | 0.615 a | 1.923 b | 0.865 ab | 0.1812 | 0.0006 |
| Glu | 32.4 | 39.6 | −0.425 a | −0.184 b | −0.159 b | −0.003 b | 0.0451 | 0.0460 |
| Gly | 14.6 | 20.4 | 0.832 a | 2.006 b | 3.136 c | 2.838 bc | 0.1372 | <0.0001 |
| Pro | 17.3 | 20.4 | 0.734 a | 1.035 ab | 1.597 c | 1.383 bc | 0.0712 | 0.0001 |
| Ser | 10.4 | 13.6 | 0.324 a | 0.712 b | 0.973 b | 0.873 b | 0.0619 | 0.0008 |
| Tyr | 5.13 | 6.31 | 0.516 a | 0.707 ab | 0.933 b | 0.970 b | 0.0482 | 0.0033 |
| Total AA 1 | 182.0 | 235.2 | 0.439 a | 0.784 b | 1.100 c | 1.060 c | 0.0784 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lachica, M.; Rodríguez-López, J.M.; González-Valero, L.; Fernández-Fígares, I. Net Portal Appearance of Amino Acids in Iberian and Landrace Pigs Fed Different Protein Content in the Diet. Animals 2023, 13, 1263. https://doi.org/10.3390/ani13071263
Lachica M, Rodríguez-López JM, González-Valero L, Fernández-Fígares I. Net Portal Appearance of Amino Acids in Iberian and Landrace Pigs Fed Different Protein Content in the Diet. Animals. 2023; 13(7):1263. https://doi.org/10.3390/ani13071263
Chicago/Turabian StyleLachica, Manuel, José Miguel Rodríguez-López, Lucrecia González-Valero, and Ignacio Fernández-Fígares. 2023. "Net Portal Appearance of Amino Acids in Iberian and Landrace Pigs Fed Different Protein Content in the Diet" Animals 13, no. 7: 1263. https://doi.org/10.3390/ani13071263