1. Introduction
Fishmeal (FM) is increasingly in demand due to its optimal nutritional qualities for fish diets, as a result of the rapid rise of worldwide aquaculture production [
1]. However, fishmeal resources are limited and prices are rising steadily. Finding alternative sources of protein has become a necessity for sustainable aquaculture [
2]. Plant protein (PP) has been used as an alternative protein source due to its relatively abundant availability and low price.
Although all-PP diets have been successfully applied to some omnivorous and herbivorous fish [
3,
4,
5], and can even be accepted by several carnivorous species [
6,
7,
8,
9], most carnivorous species are more sensitive to the negative aspects of PP, including poor palatability, essential nutrient deficiencies, and antinutritional factors [
10,
11].
With a body length of up to 30 cm, the silver sillago (
Sillago sihama Forskál,
S. sihama) belongs to the Perciformes, Sillago family, and
Sillago genus, and is mostly found in the coastal waters of the Indian Ocean and the eastern coast of the Pacific Ocean [
12]. It is highly loved for its outstanding flesh quality, high economic and nutritional value, and plays a major part in fishing activities in China’s coastal waters as an important economic fish in China’s coastal districts [
13]. However, due to overfishing, the natural resources of
S. sihama have been dropping in recent years, and market prices have increased, resulting in younger individuals, smaller groups, and decreased yields [
14]. Since 2007, research on
S. sihama artificial breeding has been conducted, and in 2012, Guangdong Ocean University effectively broke through the key technology of
S. sihama artificial breeding [
15]. At the moment, studies on
S. sihama are mostly focused on morphology [
16], genetics [
17,
18], muscle nutrition [
19], and peripheral blood cells [
20], among other factors. However, there have only been a few studies on
S. sihama diet, focusing on the appropriate protein and lipid levels [
21], vitamin A, B, and C requirements [
22,
23,
24], and the replacement of FM with dehulled soybean meal [
25].
The liver is a frontline organ involved in regulating metabolic processes, hormone production, detoxification, and immune responses [
26,
27]. Liver disease and fatty liver have become stubborn problems plaguing intensive fish farming, and are often associated with an unbalanced nutrient mix in commercial diets, or the use of poor-quality diet ingredients [
27]. In contrast to FM, plant protein limiting factors can cause metabolic disturbances, chronic inflammation, apoptosis, and immunosuppression in fish, ultimately leading to liver damage [
28,
29,
30].
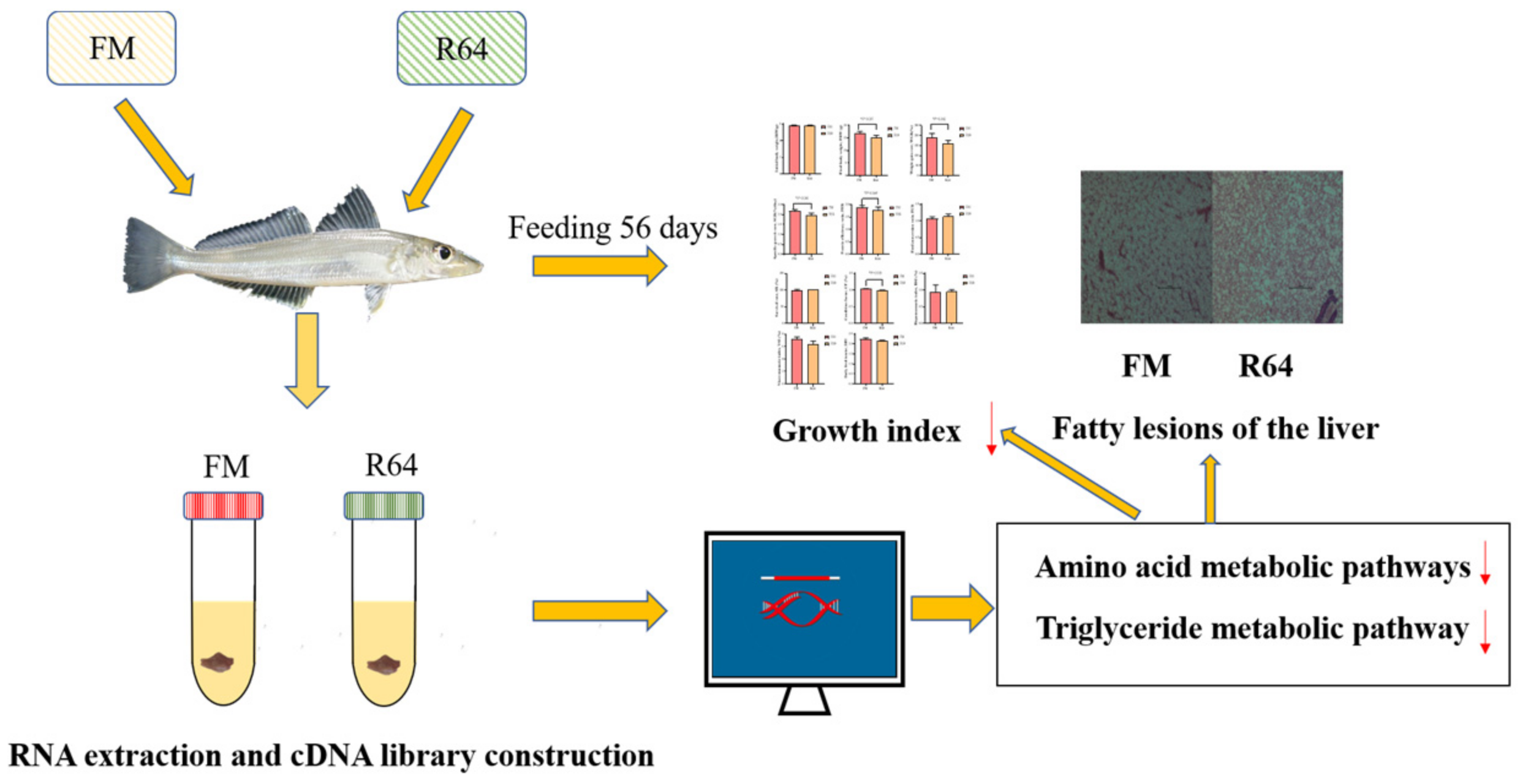
To date, there has been little work to map the regulatory mechanisms associated with the hepatic metabolic response of
S. sihama fed a high-level low-gossypol cottonseed meal (LCSM) diet. The aim of this study was to investigate the potential mechanisms of metabolic regulation of growth performance, liver health, and the liver transcriptome in
S. sihama fed a normal FM or high-level PP diet (
Figure 1).
4. Discussion
Low-gossypol cottonseed meal has been employed as a high-quality protein source in grass carp (
Ctenopharyngodon idellus) [
39], crucian carp (
Carassius auratus) [
40], black sea bass (
Centropristis striata) [
41], and blunt snout bream (
Megalobrama amblycephala) [
42]. In the current investigation, we discovered that high amounts of LCSM in the diet had a negative effect on
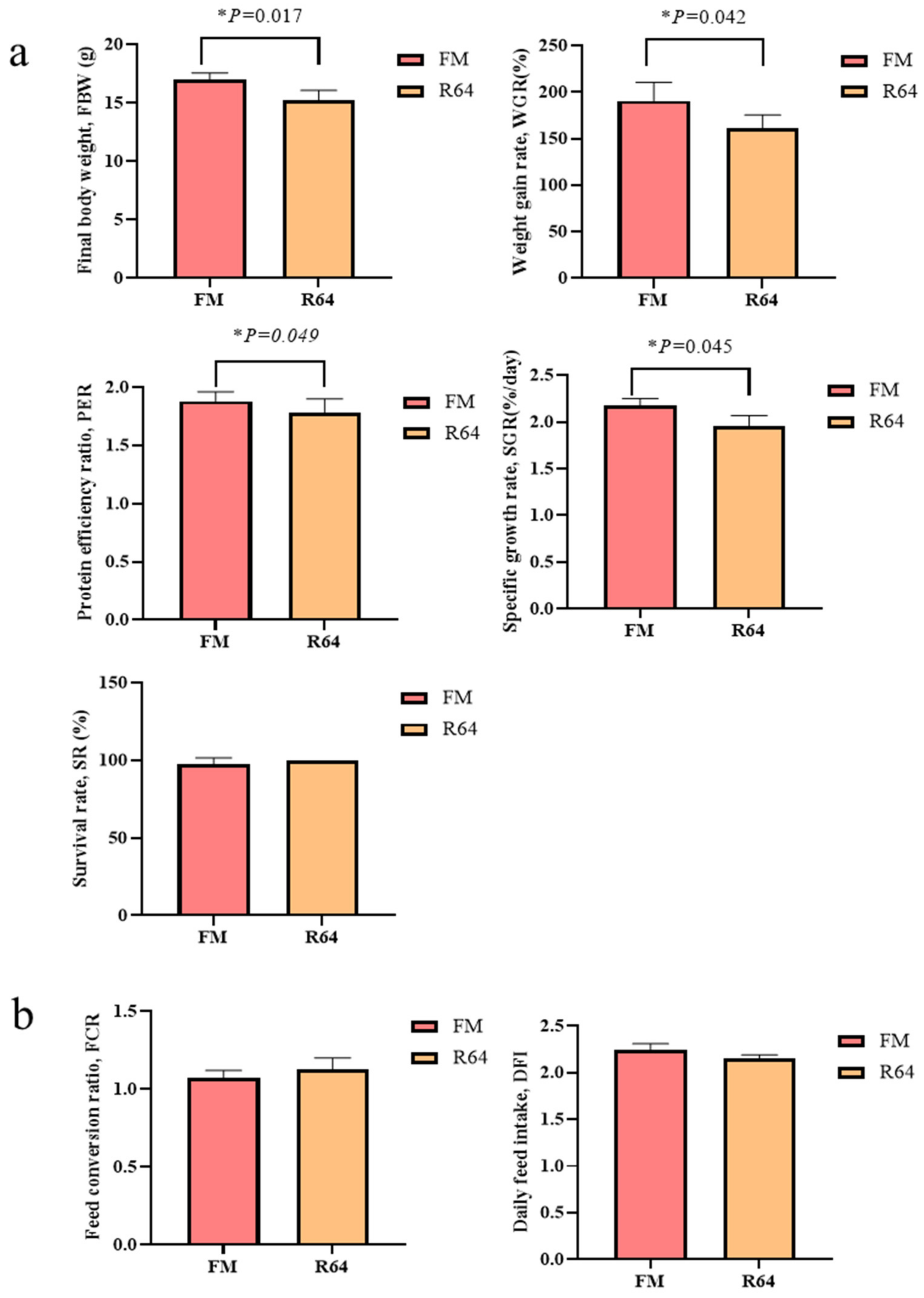
S. sihama growth indicators. The decrease in growth indicators was most likely caused by a reduction in dietary intake. In the studies of blunt snout bream [
42] and black sea bass [
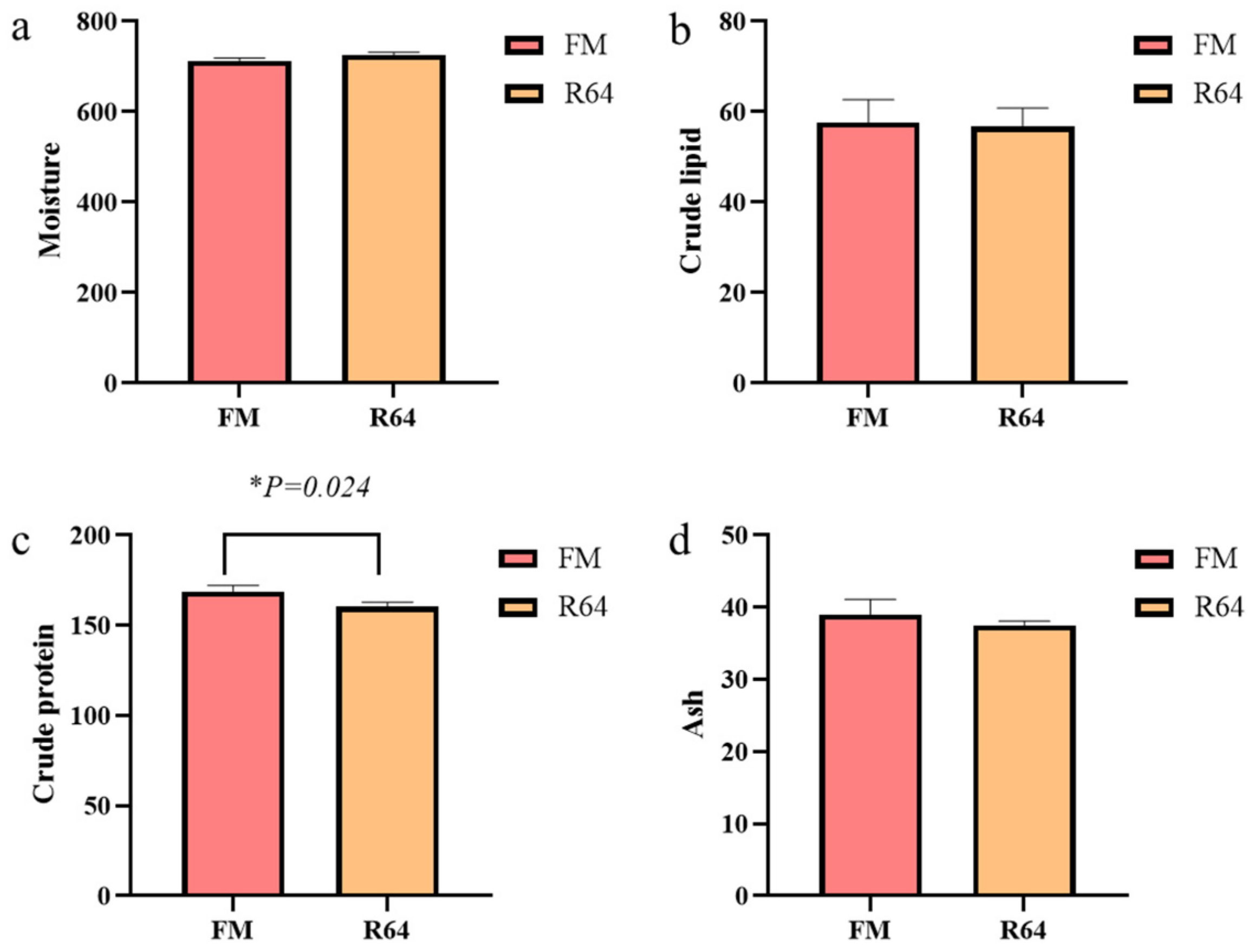
41], it was shown that high levels of PP in the diet reduced the feeding of fish. Previous research suggested that various levels of PP substituting fishmeal in the diet may have induced changes in body composition on a wet matter basis [
39,
40,
43,
44]. In this study, the whole-body crude protein content of fish given the FM diet was significantly higher than that of fish fed the R64 diet. The LCSM replacement fishmeal had no effect on whole-body crude lipid levels. Because this meal was less appealing and was consumed at a much lower rate than the FM diet, the comparatively low whole-body crude protein levels in fish fed the high level of LCSM may indicate greater protein catabolism with less protein storage. Fish fed the R64 diet had 72.58% moisture, while fish fed the FM diet had 71.32% moisture (
Figure 3a). These moisture content changes were also likely attributable to decreased palatability and consumption of the high LCSM diet with less protein storage, resulting in a proportional increase in moisture content while ash levels remained the same. Experiments with PP to substitute fishmeal yielded comparable effects in hybrid snakehead (
Channa maculata ♀ ×
Channa argus ♂) [
45], cobia (
Rachycentron canadum) [
46], red sea bream (
Pagrus major) [
47], and silver crucian carp (
Carassius auratus gibelio ♀ ×
Cyprinus carpio ♂) [
48].
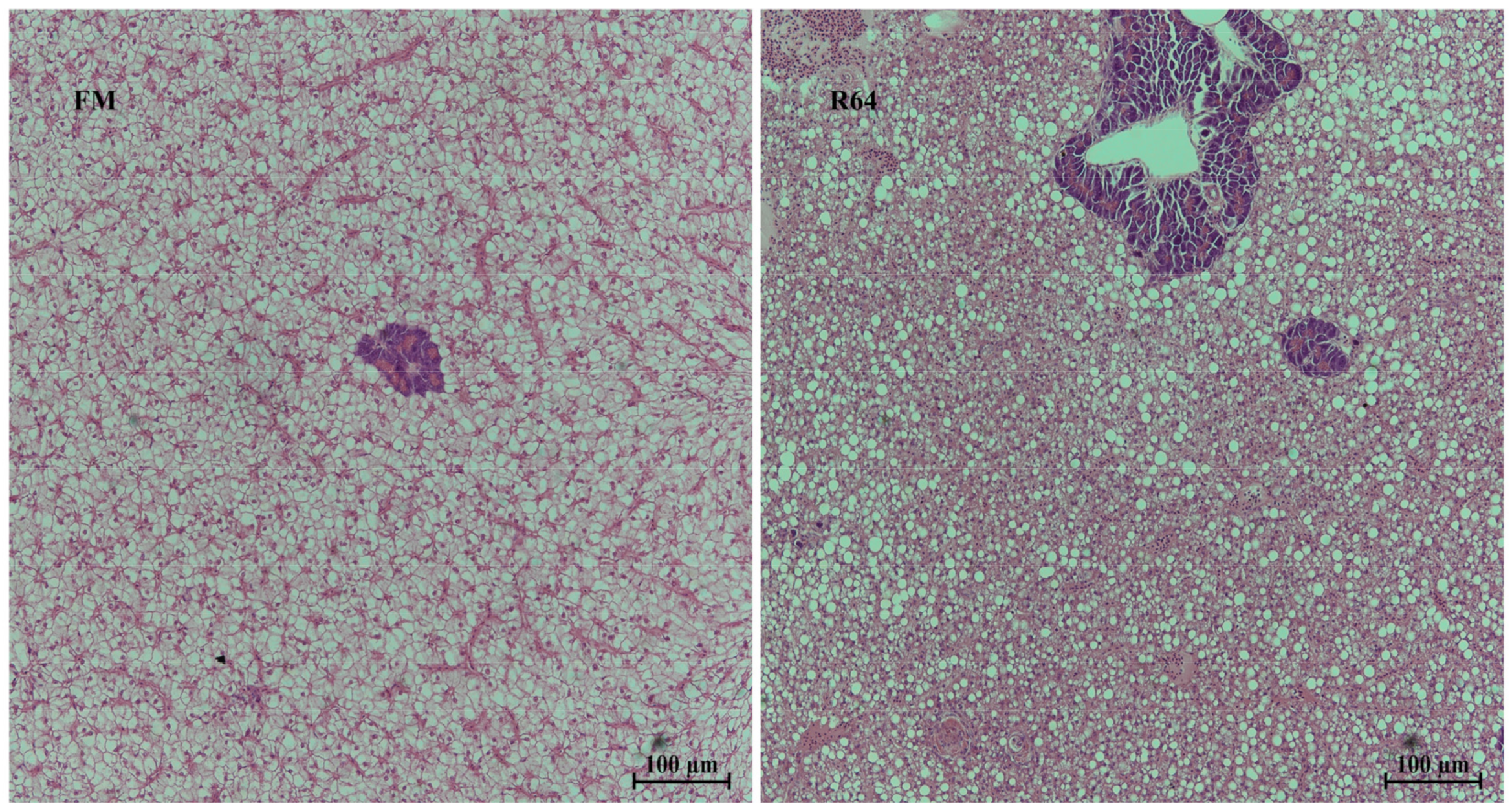
Histological changes are an important aspect in understanding the pathological alterations associated with the nutritional origin of the fish. The replacement of fishmeal by LCSM in the diet led to a number of histopathological changes in
S. sihama’s liver, including diffuse lipid vacuolation, hepatocyte hypertrophy, nuclear consolidation, irregular arrangement of hepatocytes and central venous endothelial cells, obscure hepatic cords, and sinusoidal stenosis. This means that, although some of the antinutritional factors have been removed after cottonseed protein has been degossypolized, the PP itself contains high levels of fiber, polysaccharides, and nonfat soluble antinutritional factors, which may still affect the health of the fish liver. Studies in grass carp [
49], Asian seabass (
Lates calcarifer) [
50], and hybrid sturgeon (
Acipenser schrenckii ♀ ×
A. baeri ♂) [
51] have also found that high levels of PP in the diet can cause negative effects on the liver.
The effects of nutrition on the liver transcriptome of tilapia (
Oreochromis niloticus) [
52], Atlantic salmon (
Salmo salar) [
53], yellow perch (
Perca flavescens) [
54], and pearl gentian grouper (
Epinephelus fuscoguttatus ♀
× Epinephelus lanceolatus ♂) [
55] have been well studied. However, the effects of nutrition on the liver transcriptome of
S. sihama have not been reported. The dietary transcriptional profile of nutritional stress (induced by very high LCSM) includes downregulation of hepatic metabolic pathways, including genes related to amino acid metabolism, glycerolipid metabolism, and the metabolism of xenobiotics. Upregulation includes pathways related to lipid metabolism and immune response. Interestingly, this profile is similar to that observed in pearl gentian grouper fed high levels of soybean meal [
55].
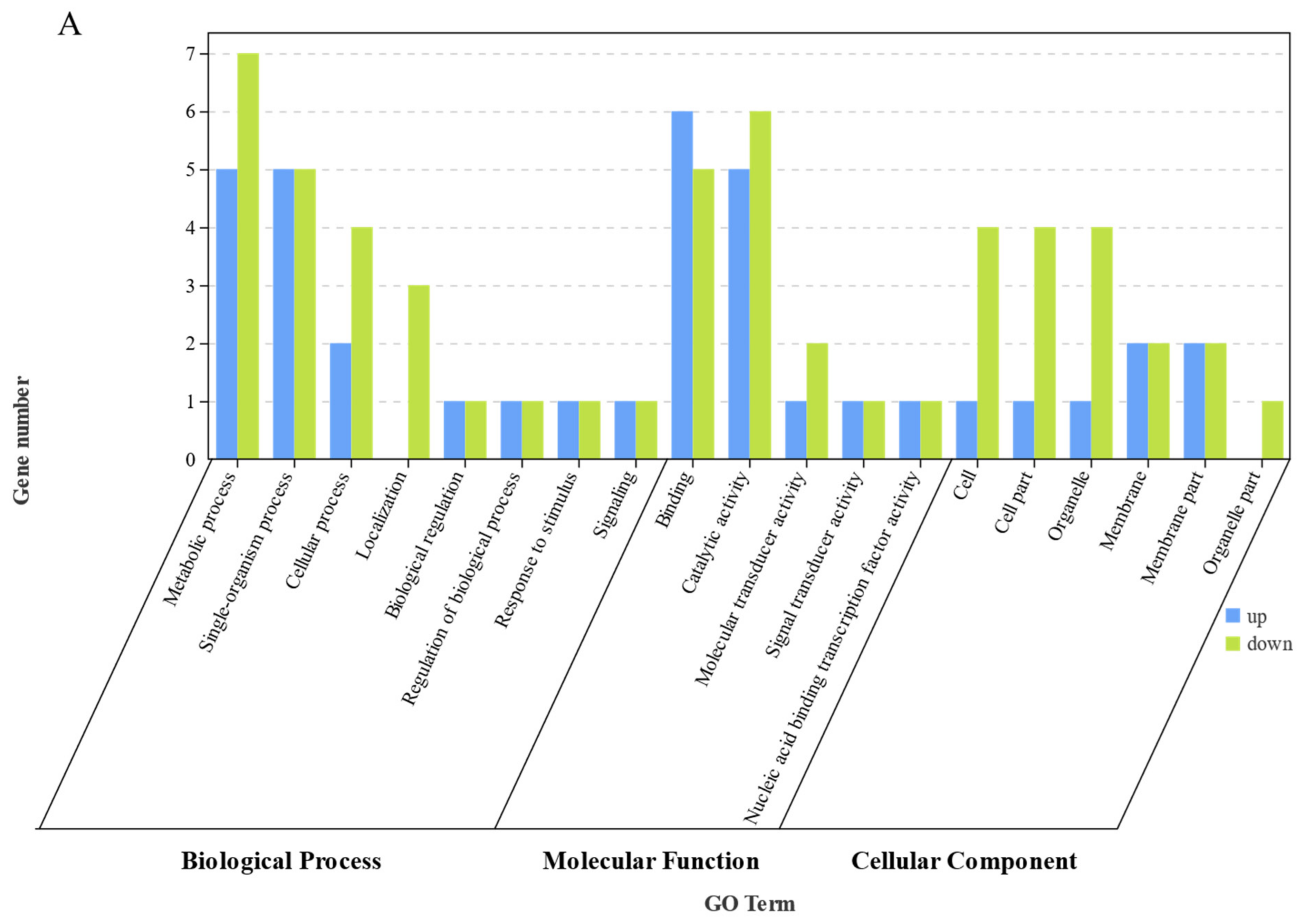
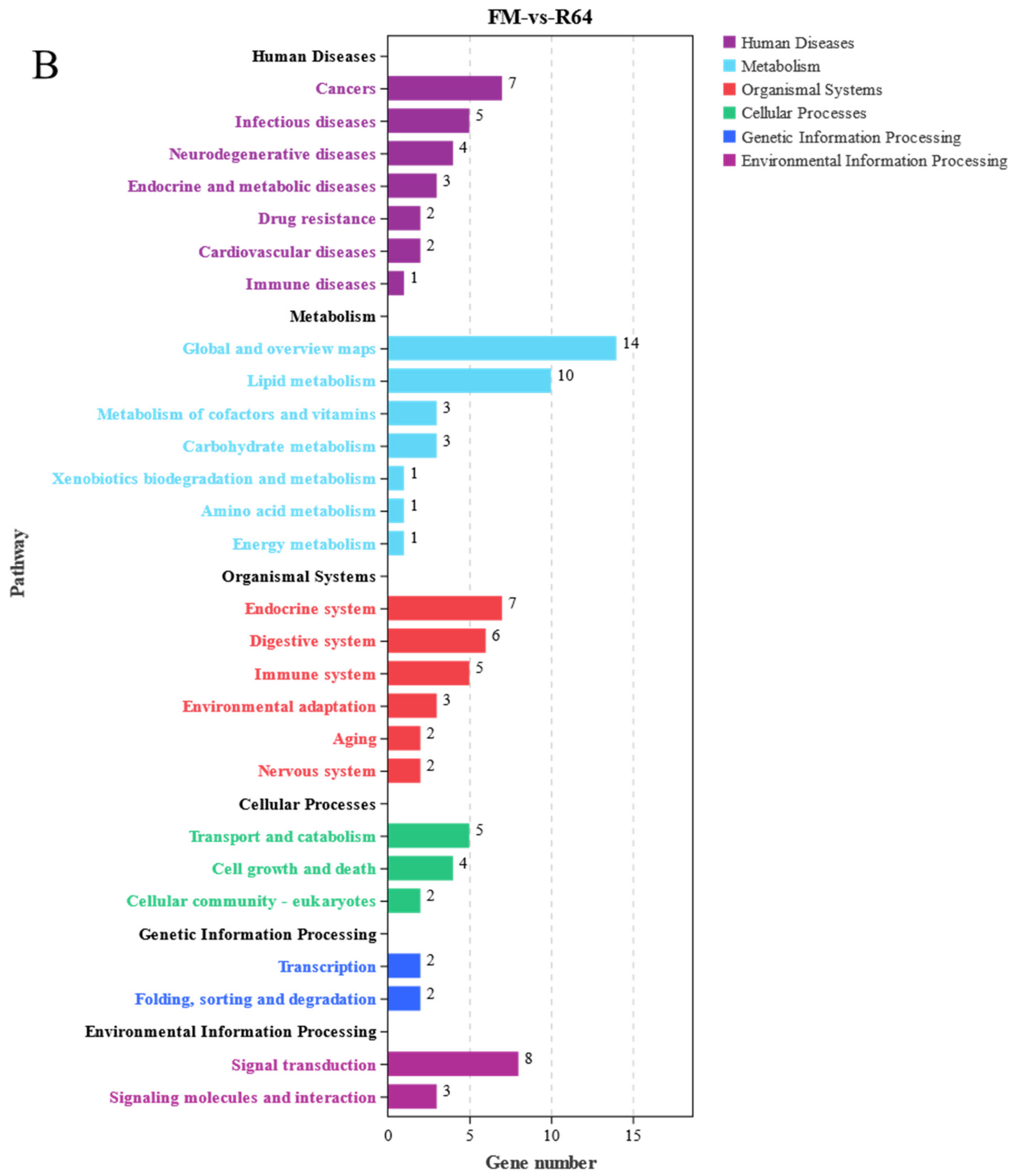
Bioinformatic analysis of the liver transcriptome of S. sihama fed FM or R64 diets showed that LCSM had the greatest effect on metabolic and immune-related genes in the liver. In the GO term enrichment differential analysis, 29 metabolic process differential genes were downregulated and 9 genes upregulated. In the KEGG enrichment analysis, regarding metabolism pathways, 73 differential genes were found, most of which were concentrated in global and overview maps (30), lipid metabolism (12), amino acid metabolism (10), and carbohydrate metabolism (7). In the GSEA, the R64 group showed downregulation of some pathways of amino acid metabolism, glucose metabolism, and lipid metabolism compared to the FM group.
A high proportion of PP in the diet in place of fishmeal is thought to be detrimental to protein synthesis. This has been demonstrated in studies with starry flounder (
Platichthys stellatus) [
56], turbot (
Scophthalmus maximus L.) [
57], and rainbow trout (
Oncorhynchus mykiss) [
58]. In the present study, the substitution of FM by LCSM in the diet by GSEA was found to have resulted in downregulation of arginine biosynthesis, tyrosine metabolism, alanine, aspartate, and glutamate metabolism, glutathione metabolism, phenylalanine metabolism, cysteine and methionine metabolism, and tryptophan metabolic pathways. The decrease in amino acid metabolic pathways may be one of the reasons for the lower growth indicators in the R64 group compared to the FM group.
Combining GSEA and KEGG enrichment analysis, we looked for the following candidate genes for amino acid metabolism: arg2, glud1, ddc, fah, gstz1, adsl, cbs, and cth.
Arginase 2 (
arg2) is generally thought to produce ornithine as a precursor for polyamine, glutamate, and proline biosynthesis [
59], synthesize urea for osmoregulation [
60,
61], and regulate arginine levels required for nitric oxide (NO) synthesis [
62]. In the present study, LCSM substituted for FM had a significant inhibitory effect on
arg2 expression, which may have been caused by the greater content of arginine in LCSM than in FM. There are a number of reports indicating that arginine, in addition to its immune-enhancing effects, can also mediate immunosuppressive effects when added in excess to the diet [
63,
64,
65]. In a recent study, it was also found that supplementation of arginine in the diet may impair the cell-mediated immune response of sea bass to some extent, reducing the number of circulating neutrophils and monocytes [
65]. DOPA decarboxylase (
ddc) is a 5′-phosphate pyridoxal (PLP)-dependent enzyme, known as aromatic 1-amino acid decarboxylase, catalyzes L-3, 4-dihydroxyphenylalanine (L-DOPA), and 5-hydroxytryptophan (5-HTP), respectively. It plays a key role in biological behavior, development, and parasite defense [
66]. In this study, the expression level of
dcc was significantly downregulated in the R64 diet, which may affect the behavior, development and parasite defense of
S. sihama.
Ddc has been found to be responsible for controlling dopamine synthesis in shrimp (
Litopenaeus vannamei), and then regulating physiological and immune responses in shrimp [
67]. In addition, studies on
Drosophila melanogaster also showed that
dcc was closely related to immunity [
68].
Glutamate dehydrogenase 1 (GLUD) is a crucial enzyme in glutamine decomposition, converting glutamate to alpha-ketoglutaric acid (α-kg) to enter the TCA cycle, where NAD(P)
+ is reduced to NAD(P)H.
Glud is triggered by the direct binding of the essential amino acid leucine, which increases glutamate deamination and the formation of α-kg. The GLUD isoenzymes GLUD1 and GLUD2 are both increased in human malignancies, allowing cancer cells to employ this pathway for growth and proliferation [
69]. GLUD1 is not only required for cancer cells to maintain the TCA cycle in order to accomplish rapid proliferation and development, but it can also activate mTORC1 [
70]. Adenylate succinate lyase (ADSL) is an essential enzyme involved in the de novo biosynthesis of purine. ADSL participates in the purine nucleotide cycle, together with adenylate succinate synthase and AMP deaminase. This cycle helps to rigidly regulate cellular metabolism by providing the number of available Krebs cycle intermediates, and pulling the adenosine kinase reaction in the direction of AMP formation. Therefore, ADSL plays an important role in both cell replication and metabolism [
71]. In this study, the expression levels of
glud and
adls in the livers of the R64 diet group were significantly decreased, indicating that LCSM may affect cell proliferation, purine metabolism, and cell replication in
S. sihama liver, thus affecting the growth and development of
S. sihama, which was also consistent with the fact that the growth indices in the R64 diet group were significantly lower than those in the FM diet group.
Cystathionine beta-synthase (CBS)-catalyzed transsulfuration converts homocysteine to cystathionine, which is converted to cysteine by cystathionine γ lyase. Additionally, CBS converts homocysteine to methionine (remethylation) or to taurine (desulfuration). This catabolic pathway alleviates the toxicity of methionine by removing excess homocysteine [
72]. In the present experiment, the R64 group
cbs expression levels were significantly lower than the FM group, suggesting that methylation was downregulated in the livers of fish fed the FM diet, and that the conversion pathway from methionine to taurine may be attenuated in the LCSM treated group. Taurine addition to the diet has been shown to improve growth in rainbow trout [
73], red sea bream [
74], Japanese flounder (
Paralichthys olivaceus), and common carp [
75]. In some fish, taurine can be synthesized from methionine via cystine. However, at high levels of PP in the diet, the rate of taurine synthesis may not be sufficient to meet the requirements of the species, thus, taurine is conditionally essential [
74,
75]. In addition, the cystathionine gamma-lyase (CTH) gene was also significantly downregulated in the R64 group. CTH catalyzes the final step in the transsulfuration pathway from methionine to cysteine, converting cystathionine to cysteine, ammonia, and 2-oxobutyric acid; glutathione synthesis in the liver is dependent on the availability of cysteine. However, the results of this experiment suggest that LCSM may affect hepatic glutathione synthesis by downregulating
cbs and
cth. Heat shock protein family 70 (
hsp70) is involved in a variety of cellular processes as a molecular chaperone, maintaining protein homeostasis during cellular stress through two opposing mechanisms: protein refolding and degradation.
Hsp70 is strongly upregulated by heat stress and toxic chemicals [
76]. In this experiment, substitution of fishmeal by LCSM caused the high expression of
hsp70 in the liver of
S. sihama, indicating that although the degossyphenols of cottonseed protein had been carried out, some other antinutritional or toxic substances in LCSM would still cause metabolic stress and damage to the liver of
S. sihama. Fumaryl acetoacetic acid hydrolase (FAH) is responsible for the final step of tyrosine metabolism. Reduced expression of
fah leads to accumulation of the toxic metabolite fumaracetic acid (FAA) in hepatocytes and proximal renal tubules, leading to mutation and apoptosis [
77]. In this experiment,
fah expression levels were significantly reduced in the R64 diet group, suggesting that LCSM substitution for FM may cause apoptosis in
S. sihama hepatocytes, as corroborated by H&E sections of the liver. Studies in pigs have found that FAH deficiency leads to diffuse and severe hepatocyte damage [
78]. The glutathione S-transferase zeta 1 (GSTZ1) gene belongs to the glutathione S-transferase (GST) superfamily, which encodes multifunctional enzymes that detoxify electrophilic compounds, such as carcinogens, mutagens, and various medicinal drugs, by binding to glutathione. One of the steps in the phenylalanine/tyrosine degradation pathway is catalyzed by this enzyme, which converts maleyl acetoacetate to fumaryl acetoacetate. Oxidative stress is caused by the absence of a comparable gene in mice [
79].
Gstz1 expression levels were significantly lower in the R64 group than in the FM group in this study, suggesting that LCSM may have an oxidative stress effect on the liver. In summary, high levels of LCSM in the diet in place of fishmeal can cause oxidative stress in the liver of
S. sihama. Studies on silvery-black porgy (
Sparidentex hasta) [
80], turbot [
76], and Ussuri catfish (
Pseudobagrus ussuriensis) [
43] have also reached a similar conclusion, that adding high levels of plant proteins to the diet can cause metabolic stress and damage of the liver.
PP replacement for fishmeal has been reported to affect lipid metabolism in trout [
81], Atlantic salmon (
Salmo salar) [
82], and hybrid grouper (
Epinephelus fuscoguttatus ♀ ×
E. lanceolatus ♂) [
83]. In this experiment, we found that the triglyceride metabolic pathway appeared to be significantly downregulated in the R64 diet by GSEA, whereas the steroid biosynthetic pathway appeared to be significantly upregulated in the R64 group compared with the FM group. Comprehensive GSEA and KEGG enrichment analysis identified the following candidate genes:
pnpla2,
mogat2-a,
lipg,
cyp51,
sqle,
sc5d,
lss, and
soat1.
Patatin-like phospholipase domain-containing protein 2 (PNPLA2), which was designated adipose triglyceride lipase (ATGL) 17 years ago [
84], is assumed to be the central rate-limiting enzyme for fatty acid mobilization in mammals [
85]. PNPLA2 specifically hydrolyses triglycerides (TG) to diacylglycerols (DG) and releases free fatty acids (FFA). In this experiment, we found a significant downregulation of
pnpla2 in the R64 group, suggesting that LCSM substitution for fishmeal may lead to abnormal hepatic lipolysis in
S. sihama. There is growing evidence that dysregulated lipolysis is associated with a number of metabolic disorders, such as fatty liver and type two diabetes [
86]. LCSM replacement of fishmeal resulted in abnormal accumulation of liver lipid in
S. sihama, as shown in
Figure 4. Studies in zebrafish and mice have also found that similar lipid deposition is observed with inhibition of
pnpla2 [
85,
87]. In the present study, there was a significant downregulation of
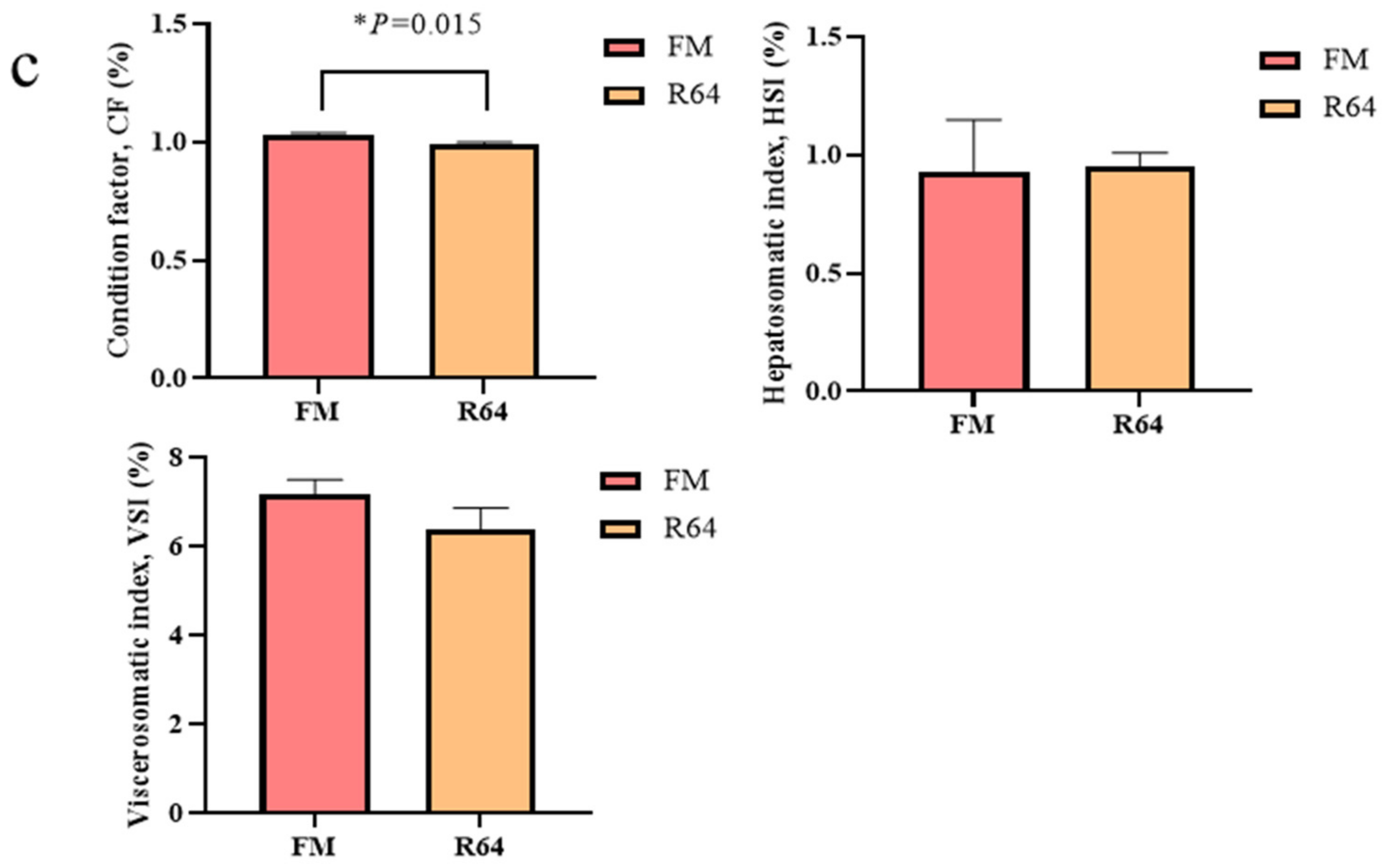
mogat2 expression levels in the R64 diet, suggesting that LCSM affects lipid synthesis in addition to lipolysis. This may also explain the small difference in whole-body crude lipid content between the FM and R64 diets, and the lack of significant differences in VSI and HSI between the two groups. In fact, in a starry flounder study, it was also found that the substitution of PP for fishmeal resulted in a decrease in lipid synthesis and catabolism [
12].
In addition, we found that
cyp51,
sqle,
sc5d,
lss, and
soat1 were all upregulated in this experiment, all of which are key genes in the cholesterol biosynthesis pathway, which means that the LCSM substitution for fishmeal actually promoted the cholesterol synthesis ability of
S. sihama. In numerous studies in the past, PP substitution for fishmeal has been found to reduce serum cholesterol levels [
88,
89]. There is also an abundance of evidence that phytosterols in plant proteins can inhibit cholesterol absorption [
90,
91,
92]. LCSM replaced fishmeal and reduced the content of cholesterol in the diet. In addition, dietary phytosterol contained in PP inhibited the absorption of cholesterol in fish, which led to the possibility that the cholesterol-satisfied growth requirement could not be obtained in the diet. Therefore, genes related to de novo cholesterol synthesis in the liver were upregulated. A study found that the growth of rainbow trout fed with added cholesterol in a PP-based diet was significantly higher than that in the nonsupplemented diet [
93], and similar results were found in channel catfish (
Ictalurus punctatus) [
94], Japanese flounder [
95], and turbot [
96], which also supported our view.
The effects of fishmeal replacement by PP on glucose metabolism is rarely studied. In this study, GSEA showed that the mutual conversion pathway of pentose and glucuronate interconversions was significantly downregulated in the R64 group compared with the FM group. Based on GSEA and KEGG enrichment analysis, we found the following candidate differential genes: eno1, fbp1, g6pd, and pgam2.
ENO1 and PGAM2 are both important enzymes in glycolysis. ENO1 catalyzes the conversion of 2-phosphoglycerate to phosphoenolpyruvate, and PGAM2 catalyzes the reversible reaction from 3-phosphoglycerate (3-PGA) to 2-phosphoglycerate (2-PGA). In this study, LCSM replacement of fishmeal resulted in significant downregulation of
eno1 and
pgam2 in the liver, indicating that LCSM replacement of fishmeal could inhibit glycolysis. It may be that the residual gossypol in LCSM exerted an inhibitory effect on liver glycolysis. In vitro studies found that decreased sperm motility of rabbit could result from a reduced energy supply after inhibition of glycolysis by gossypol [
97]. In addition, gossypol is thought to inhibit glycolysis by inhibiting lactate dehydrogenase isozyme 5 [
98].
Fructose-bisphosphatase 1(FBP1) catalyzes the hydrolysis of fructose 1,6-bisphosphate to fructose 6-phosphate, acting as a rate-limiting enzyme in the process of gluconeogenesis. Glucose-6-phosphate dehydrogenase (G6PD) is the final step in gluconeogenesis. In this experiment, both
fbp1 and
g6pd were significantly downregulated, indicating that fishmeal replacement by LCSM can cause inhibitory effects on
S. sihama gluconeogenesis. This may be because the level of glycogenic amino acids in LCSM is less than that in the fishmeal diet. The total content of glycogenic amino acids in fishmeal is 472 g/kg, while the total content of glycogenic amino acids in LCSM is 441.2 g/kg (
Table 1). The reduction of glycogenic substrates may be one reason for the inhibition of gluconeogenesis. In addition, the availability of amino acids in LCSM may be low. In silvery-black porgy, liver dysfunction is caused by loss of hepatocyte integrity, disrupted protein synthesis, and low amino acid availability [
99]. In this experiment, we only supplemented methionine and lysine, while some glycogenic amino acids, such as alanine and aspartic acid, in the LCSM diet were less than the FM diet. In a Totoaba (
Totoaba macdonaldi) study [
100], it was found that more than 45% soybean meal in the diet affected the FBPase activity in the liver of Totoaba. Similar results were also reported in rainbow trout, where lower FBPase activity was associated with lower alanine content in the diet [
101]. Therefore, we believe that FBPase activity is limited by gluconeogenesis, which can also be attributed to low protein digestibility [
102], which may affect alanine availability [
103]. In addition, a turbot study also found that adding high levels of soybean meal and cottonseed meal to the diet also inhibited G6PD activity, and thus gluconeogenesis [
104]. These effects may be caused by the inefficient use of PP by carnivorous fish.

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}