
Non-Invasive Sex Determination of Nestlings and Adult Bonelli’s Eagles Using Morphometrics



Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Sample Collection
2.3. Molecular Sexing
2.4. Morphometric Measurements
2.5. Statistical Analyses
3. Results
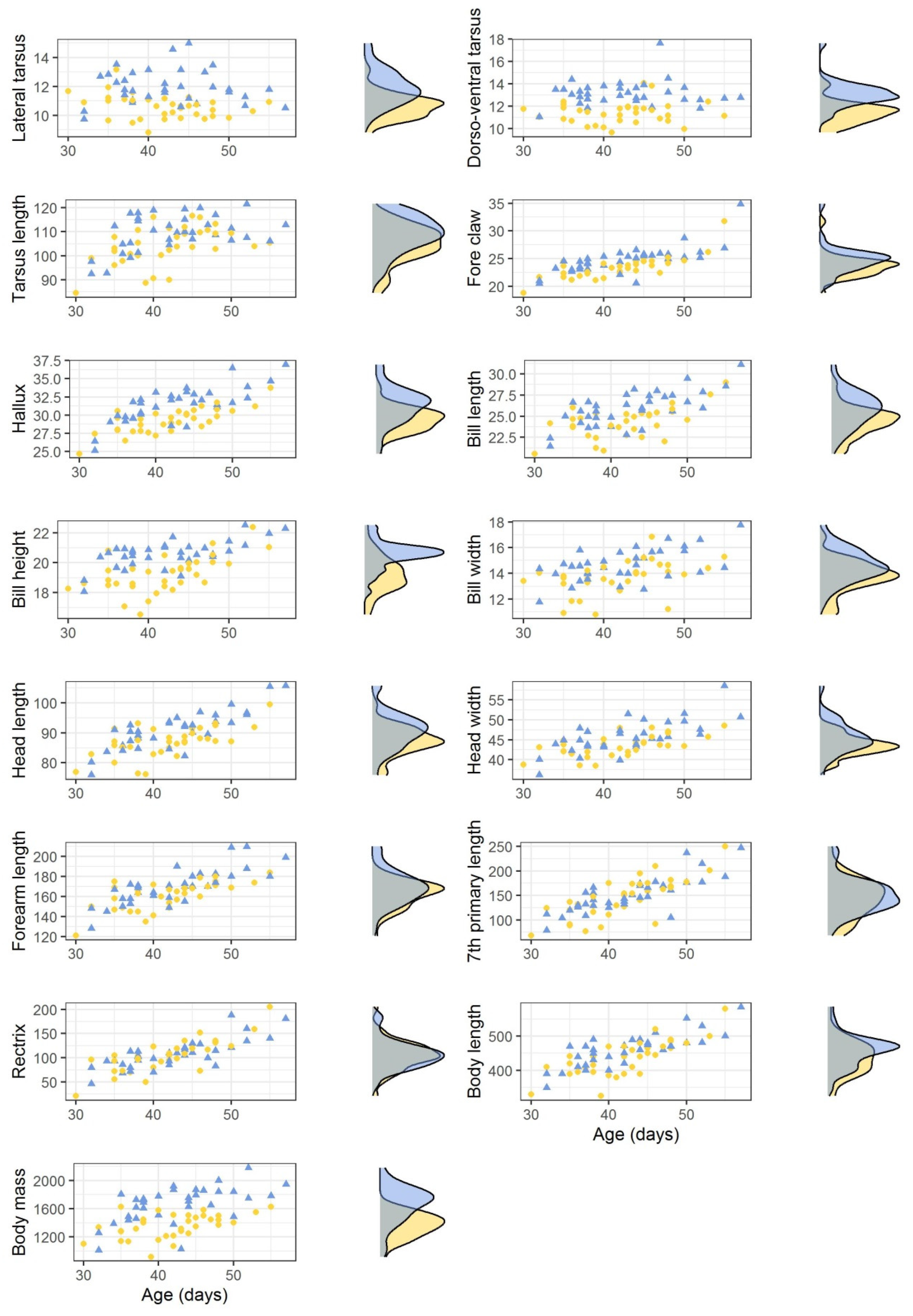
3.1. Bonelli’s Eagle Nestlings
(tarsus length) − 0.323 × (fore claw) + 0.644 × (hallux) − 0.097 × (bill length) +
0.706 × (bill height) + 0.383 × (bill width) − 0.003 × (head length) − 0.057 ×
(head width) − 0.011 × (forearm length) + 0.008 × (7th primary) − 0.036 ×
(rectrix) + 0.004 × (body length)
(fore claw) + 0.443 × (hallux) − 0.093 × (bill length) + 0.662 × (bill height) +
0.378 × (bill width) + 0.016 × (head length) − 0.082 × (head width) − 0.011 ×
(body length)
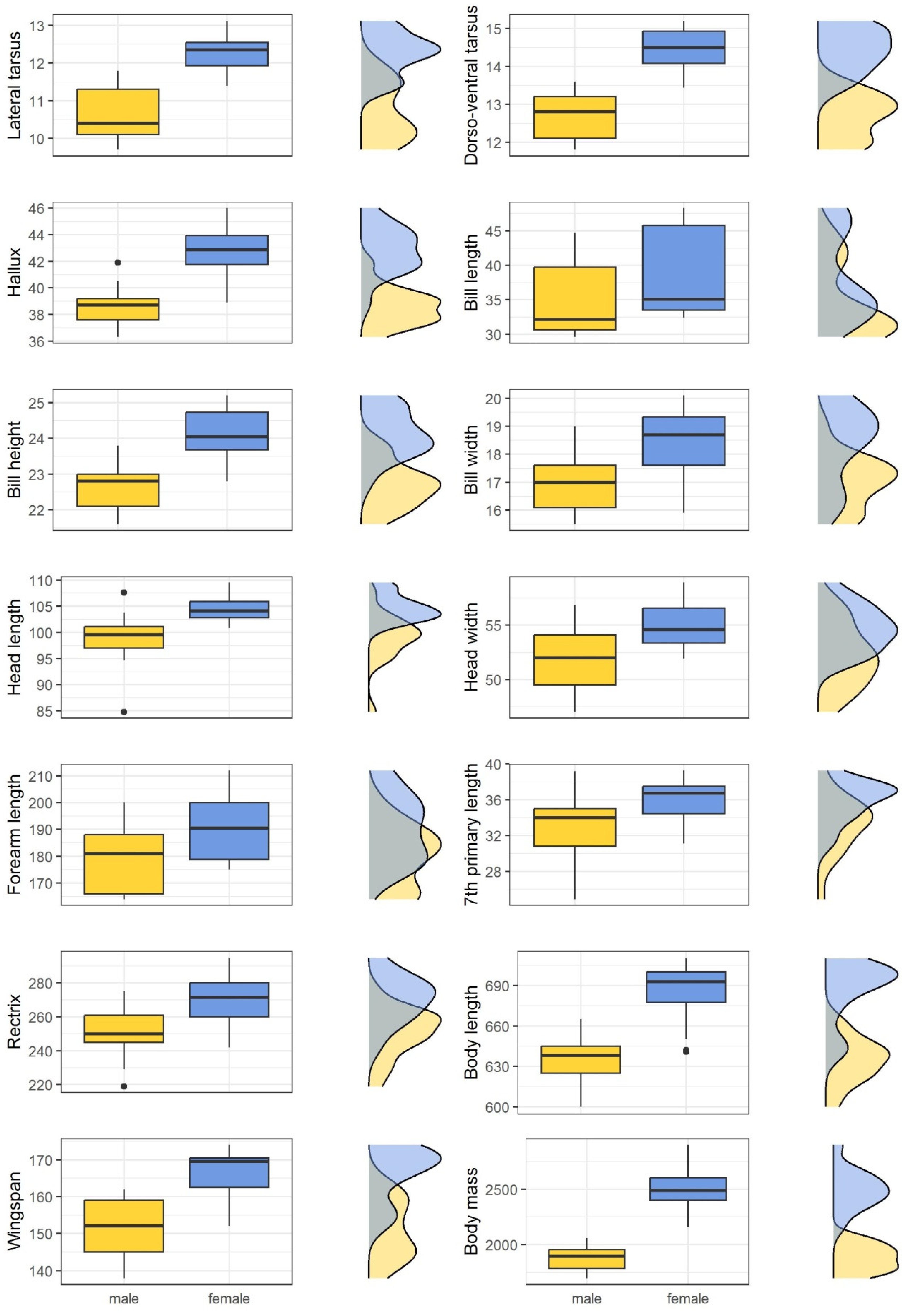
3.2. Bonelli’s Eagle Adults
(hallux) + 0.186 × (bill length) + 0.380 × (bill height) − 0.450 × (bill width) −
0.119 × (head length) − 0.102 × (head width) + 0.044 × (forearm length) − 0.064
× (7th primary) + 0.026 × (rectrix) + 0.030 × (body length) − 0.008 × (wingspan)
(hallux) + 0.067 × (bill length) + 0.537 × (bill height) − 0.450 × (bill width) −
0.096 × (head length) − 0.072 × (head width) + 0.037 × (body length) − 0.017 ×
(wingspan)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cadahía, L.; López-López, P.; Urios, V.; Soutullo, Á.; Negro, J.J. Natal dispersal and recruitment of two Bonelli’s Eagles Aquila fasciata: A four-year satellite tracking study. Acta Ornithol. 2009, 44, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Cadahía, L.; López-López, P.; Urios, V.; Negro, J.J. Satellite telemetry reveals individual variation in juvenile Bonelli’s eagle dispersal areas. Eur. J. Wildl. Res. 2010, 56, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Newton, I.; Marquiss, M.; Rothery, P. Age structure and survival in a sparrowhawk population. J. Anim. Ecol. 1983, 52, 591–602. [Google Scholar] [CrossRef]
- Clutton-Brock, T. Sex ratio variation in birds. Ibis 1986, 128, 317–329. [Google Scholar] [CrossRef]
- Griffiths, R.; Tiwari, B. Sex of the last wild Spix’s macaw. Nature 1995, 375, 454. [Google Scholar] [CrossRef] [PubMed]
- Muriel, R.; Casado, E.; Schmidt, D.; Calabuig, C.P.; Ferrer, M. Morphometric sex determination of young Ospreys Pandion haliaetus using discriminant analysis. Bird Study 2010, 57, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.; Delecourt, C. Sex identification in the Spanish imperial eagle. J. Field Ornithol. 1992, 63, 359–364. [Google Scholar]
- Wheeler, P.; Greenwood, P.J. The evolution of reversed sexual dimorphism in birds of prey. Oikos 1983, 40, 145–149. [Google Scholar] [CrossRef]
- Montgomerie, R.; Lundberg, A. Reversed sexual dimorphism in raptors: Which sex changed size? Oikos 1989, 56, 283–286. [Google Scholar] [CrossRef]
- Schoenjahn, J.; Pavey, C.R.; Walter, G.H. Why female birds of prey are larger than males. Biol. J. Linn. Soc. 2020, 129, 532–542. [Google Scholar] [CrossRef]
- Krüger, O. The evolution of reversed sexual size dimorphism in hawks, falcons and owls: A comparative study. Evol. Ecol. 2005, 19, 467–486. [Google Scholar] [CrossRef]
- Morrison, J.; Maltbie, M. Methods for gender determination of Crested Caracaras. J. Raptor Res. 1999, 33, 128–133. [Google Scholar]
- López-López, P.; Gil, J.A.; Alcántara, M. Morphometrics and sex determination in the endangered Bearded Vulture (Gypaetus barbatus). J. Raptor Res. 2011, 45, 361–366. [Google Scholar] [CrossRef] [Green Version]
- García, V.; Moreno-Opo, R.; Tintó, A. Sex differentiation of Bonelli’s eagle Aquila fasciata in western europe using morphometrics and plumage colour patterns. Ardeola 2013, 60, 261–277. [Google Scholar] [CrossRef]
- Harmata, A.; Montopoli, G. Morphometric sex determination of North American Golden Eagles. J. Raptor Res. 2013, 47, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Boucheker, A.; Nedjah, R.; Prodon, R.; Gillingham, M.; Dechaume-Moncharmont, F.-X.; Béchet, A.; Samraoui, B. Cohort effect on discriminant rate: The case of greater flamingo (Phoenicopterus roseus) chicks sexed with morphological characters. Web Ecol. 2020, 20, 153–159. [Google Scholar] [CrossRef]
- Griffiths, R.; Daan, S.; Dijkstra, C. Sex identification in birds using two CHD genes. Proc. R. Soc. London. Ser. B Biol. Sci. 1996, 263, 1251–1256. [Google Scholar]
- Fridolfsson, A.-K.; Ellegren, H. A simple and universal method for molecular sexing of non-ratite birds. J. Avian Biol. 1999, 30, 116–121. [Google Scholar] [CrossRef]
- Dubiec, A.; Zagalska-Neubauer, M. Molecular techniques for sex identification in birds. Biol. Lett. 2006, 43, 3–12. [Google Scholar]
- Balbontín, J.; Ferrer, M.; Casado, E. Sex determination in booted eagles (Hieraaetus pennatus) using molecular procedures and discrimiant function analysis. J. Raptor Res. 2001, 35, 20–23. [Google Scholar]
- Dykstra, C.R.; Mays, H.L.; Hays, J.L.; Simon, M.M.; Wegman, A.R. Sexing Adult and Nestling Red-Shouldered Hawks Using Morphometrics and Molecular Techniques1. J. Raptor Res. 2012, 46, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Cramp, S.; Ferguson-Lees, I.J.; Gillmor, R.; Hollom, P.; Hudson, R.; Nicholson, E.M.; Ogilvie, M.; Olney, P.; Voous, K.; Wattel, J. The Birds of the Western Palearctic; Oxford University Press: Oxford, UK, 1983; p. 41. [Google Scholar]
- Ferguson-Lees, J.; Christie, D.A. Raptors of the World; Houghton Mifflin Harcourt: Boston, MA, USA, 2001. [Google Scholar]
- Mañosa, S.; Codina, J. Age estimation and growth patterns in nestling Bonelli’s Eagles. J. Raptor Res. 1995, 29, 273–275. [Google Scholar]
- Palma, L.; Mira, S.; Cardia, P.; Beja, P.; Guillemaud, T.; Ferrand, N.; Cancela, M.; da Fonseca, L. Sexing Bonelli’s Eagle nestlings: Morphometrics versus molecular techniques. J. Raptor Res. 2001, 35, 187–193. [Google Scholar]
- Redondo-Gómez, D.; Bautista, J.; Gil-Sánchez, J.; Pares, F.; Hernandez-Matias, A.; Resano-Mayor, J.; Real, J.; Pacteau, C.; Madero, A.; Moleón, M. Towards accurate and simple morphometric sex differentiation in Bonelli’s Eagle Aquila fasciata nestlings: Interpopulation variations and influence of growth conditions. Avian Biol. Res. 2022, 15, 3–12. [Google Scholar] [CrossRef]
- Del Moral, J.C.; Molina, B. El águila Perdicera en España, Población Reproductora en 2018 y Método de Censo; SEO/BirdLife: Madrid, Spain, 2018. [Google Scholar]
- López-López, P.; Perona, A.; Egea-Casas, O.; Morant, J.; Urios, V. Tri-axial accelerometry shows differences in energy expenditure and parental effort throughout the breeding season in long-lived raptors. Curr. Zool. 2022, 68, 57–67. [Google Scholar] [CrossRef]
- Morollón, S.; Urios, V.; López-López, P. Home-Range Size and Space Use of Territorial Bonelli’s Eagles (Aquila fasciata) Tracked by High-Resolution GPS/GSM Telemetry. Diversity 2022, 14, 1082. [Google Scholar] [CrossRef]
- Morollón, S.; Urios, V.; López-López, P. Fifteen days are enough to estimate home-range size in some long-lived resident eagles. J. Ornithol. 2022, 163, 849–854. [Google Scholar] [CrossRef]
- Perona, A.M.; Urios, V.; López-López, P. Holidays? Not for all. Eagles have larger home ranges on holidays as a consequence of human disturbance. Biol. Conserv. 2019, 231, 59–66. [Google Scholar] [CrossRef]
- López-López, P.; García-Ripollés, C.; Urios, V. Population size, breeding performance and territory quality of Bonelli’s eagle Hieraaetus fasciatus in eastern Spain. Bird Study 2007, 54, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Gil-Sánchez, J.M. Efecto de la altitud y de la disponibilidad de presas en la fecha de puesta del águila-azor perdicera (Hieraaetus fasciatus) en la provincia de Granada (SE de España). Ardeola 2000, 47, 1–8. [Google Scholar]
- López-López, P. Potential negative effects of the installation of video surveillance cameras in raptors’ nests. Sci. Rep. 2022, 12, 21969. [Google Scholar] [CrossRef] [PubMed]
- Sansano-Maestre, J.; Garijo-Toledo, M.; Gomez-Munoz, M. Prevalence and genotyping of Trichomonas gallinae in pigeons and birds of prey. Avian Pathol. 2009, 38, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Sudo-Yamaji, A.; Abe, M.; Murase, T.; Tsubota, T. Sex identification by alternative polymerase chain reaction methods in Falconiformes. Zool. Sci. 2003, 20, 339–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zar, J. Biostatistical Analysis, 5th ed.; Pearson: New York, NY, USA, 2014. [Google Scholar]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. 2023. Version 0.7.2. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 28 March 2023).
- Dillon, W.R.; Goldstein, M. Multivariate Analysis: Methods and Applications; Wiley: New York, NY, USA, 1984; ISBN 0-471-08317-8. [Google Scholar]
- Dechaume-Moncharmont, F.-X.; Monceau, K.; Cezilly, F. Sexing birds using discriminant function analysis: A critical appraisal. Auk 2011, 128, 78–86. [Google Scholar] [CrossRef]
- McLachlan, G.J. Discriminant Analysis and Statistical Pattern Recognition; John Wiley & Sons: Hoboken, NJ, USA, 2005; ISBN 978-0-471-72528-2. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing 2022. Available online: http://www.r-project.org/index.html (accessed on 28 March 2023).
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. Posit PBC Dplyr: A Grammar of Data Manipulation 2023. Version 1.1.1. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 28 March 2023).
- Kuhnaut, M.; Wing, J.; Weston, S.; Williams, A.; Keefer, C.; Engelhardt, A.; Cooper, T.; Mayer, Z.; Kenkel, B.; R Core Team; et al. Caret: Classification and Regression Training 2022; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Makowskiaut, D.; Najberg, H.; Simko, V.; Epskamp, S. psycho: Efficient and Publishing-Oriented Workflow for Psychological Science 2021. Version 0.6.1. Available online: https://CRAN.R-project.org/package=psycho (accessed on 28 March 2023).
- Müller, K.; Wickham, H.; Francois, R.; Bryan, J. Tibble: Simple Data Frames 2022. Version 3.2.1. Available online: https://CRAN.R-project.org/package=tibble (accessed on 28 March 2023).
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J.; Freidank, M.; Cai, J.; Protivinsky, T. Corrplot: Visualization of a Correlation Matrix 2021. Version 0.92. Available online: https://CRAN.R-project.org/package=corrplot (accessed on 28 March 2023).
- Pay, J.M.; Katzner, T.E.; Wiersma, J.M.; Brown, W.E.; Hawkins, C.E.; Proft, K.M.; Cameron, E.Z. Morphometric Sex Identification of Nestling and Free-Flying Tasmanian Wedge-Tailed Eagles (Aquila audax fleayi). J. Raptor Res. 2021, 55, 539–551. [Google Scholar] [CrossRef]
- Bortolotti, G.R. Physical development of nestling bald eagles with emphasis on the timing of growth events. Wilson Bull. 1984, 96, 524–542. [Google Scholar]
- Donohue, K.C.; Dufty, A.M., Jr. Sex determination of Red-tailed Hawks (Buteo jamaicensis calurus) using DNA analysis and morphometrics. J. Field Ornithol. 2006, 77, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Bortolotti, G.R. Influence of sibling competition on nestling sex ratios of sexually dimorphic birds. Am. Nat. 1986, 127, 495–507. [Google Scholar] [CrossRef]
- McPherson, S.C.; Brown, M.; Downs, C.T. Gender-related morphometric differences in mature and nestling Crowned Eagles, with comments on ringing of eagle nestlings in KwaZulu-Natal, South Africa. Ostrich 2017, 88, 195–200. [Google Scholar] [CrossRef]
- Poole, K. Determining age and sex of nestling Gyrfalcons. J. Raptor Res. 1989, 23, 45–47. [Google Scholar]
- Sodhi, N. Growth of nestling Merlins, Falco columbarius. Can. Field-Nat. Ott. ON 1992, 106, 387–389. [Google Scholar]
- Choi, C.-Y.; Nam, H.-Y.; Park, J.-G.; Bing, G.-C.; Park, C.; Cho, S.-Y. Morphometrics and sexual dimorphism of Chinese Goshawks (Accipiter soloensis). J. Raptor Res. 2013, 47, 385–391. [Google Scholar] [CrossRef]
- Smith, N.R.; Afton, A.D.; Hess, T.J., Jr. Morphometric sex determination of after-hatch-year Bald Eagles in Louisiana. J. Raptor Res. 2016, 50, 301–304. [Google Scholar] [CrossRef]
- Barton, N.W.; Houston, D. A comparison of digestive efficiency in birds of prey. Ibis 1993, 135, 363–371. [Google Scholar] [CrossRef]
- Bordner, Z.E.; McCabe, R.A.; Brinker, D.; Rosenfield, R.N.; Jacobs, E.A.; England, C.; Wilson, M.; Goodrich, L.J. Broad-Winged Hawk Size Varies by Sex and Latitude in North America. J. Raptor Res. 2022, 56, 412–422. [Google Scholar] [CrossRef]
- Soutullo, A.; López-López, P.; Urios, V. Incorporating spatial structure and stochasticity in endangered Bonelli’s eagle’s population models: Implications for conservation and management. Biol. Conserv. 2008, 141, 1013–1020. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Matias, A.; Real, J.; Moleon, M.; Palma, L.; Sanchez-Zapata, J.; Pradel, R.; Carrete, M.; Gil-Sanchez, J.; Beja, P.; Balbontin, J.; et al. From local monitoring to a broad-scale viability assessment: A case study for the Bonelli’s Eagle in western Europe. Ecol. Monogr. 2013, 83, 239–261. [Google Scholar] [CrossRef] [Green Version]
- Real, J.; Manosa, S. Dispersal of juvenile and immature Bonelli’s Eagles in northeastern Spain. J. Raptor Res. 2001, 35, 9–14. [Google Scholar]
- Hernandez-Matias, A.; Real, J.; Pradel, R.; Ravayrol, A.; Vincent-Martin, N.; Bosca, F.; Cheylan, G. Determinants of territorial recruitment in Bonelli’s eagle (Aquila fasciata) populations. Auk 2010, 127, 173–184. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Variable | Description | Instrument | Units |
|---|---|---|---|
| Lateral tarsus | Lateral width of tarsus-metatarsus in the narrowing part situated above the beginning of the metatarsal angle. | Calipers | mm |
| Dorso-ventral tarsus | Dorso-ventral width of tarsus-metatarsus in the narrowing part situated above the beginning of the metatarsal angle. | Calipers | mm |
| Tarsus length | Distance from the tarsometatarsal joint to the articulation of the middle toe. | Calipers | mm |
| Fore claw length | Length of the claw of the central toe. | Calipers | mm |
| Hallux length | Length of the claw of the hind toe. | Calipers | mm |
| Bill length | Length from the distal end of the bill to the distal dorsal edge of the cere. | Calipers | mm |
| Bill height | Length from the distal edge of the cere to the base of the bill, placing the caliper perpendicular to the bill. | Calipers | mm |
| Bill width | Width of the bill taken by the calipers placed against the corners of the bill at the level of the nostrils. | Calipers | mm |
| Head width | Width of the skull measured behind the eyes in the auricular region. | Calipers | mm |
| Head length | Maximum length from the back of the skull to the distal end of the bill. | Calipers | mm |
| Seventh primary length | Length of the seventh primary feather (counting from the outer tip of the wing inwards) to its insertion. | Metal ruler | mm |
| Forearm length | Length of the forearm ventral to the shoulder, positioning the ruler at the elbow to the distal end of the ulna. | Metal ruler | mm |
| Rectrix | Length of the central feather of the tail to its insertion. | Metal ruler | mm |
| Wingspan | Distance between the tips of the extended wings. | Tape measure | mm |
| Body length | Length from the end of the bill to the end of the central feather of the tail with the bird stretched out on a table. | Tape measure | mm |
| Body mass | Body mass weighted on a digital balance. | Digital balance | g |
| Variable | Sex | Mean | sd | Median | Min | Max | p-Value | Effect Size | % Dimorphism |
|---|---|---|---|---|---|---|---|---|---|
| Lateral tarsus | male | 10.60 | 0.83 | 10.65 | 8.81 | 13.16 | <0.001 | 0.577 | −11.34 |
| female | 11.96 | 1.18 | 11.77 | 9.72 | 14.98 | ||||
| Dorso-ventral tarsus | male | 11.40 | 0.96 | 11.43 | 9.64 | 14.08 | <0.001 | 0.676 | −12.87 |
| female | 13.09 | 1.15 | 12.98 | 11.03 | 17.62 | ||||
| Tarsus length | male | 104.43 | 7.92 | 104.71 | 84.53 | 116.55 | 0.006 | 0.309 | −4.55 |
| female | 109.41 | 7.44 | 109.76 | 92.46 | 121.55 | ||||
| Fore claw | male | 23.48 | 2.08 | 23.64 | 18.78 | 31.74 | 0.002 | 0.344 | −5.19 |
| female | 24.77 | 2.54 | 24.96 | 20.47 | 34.88 | ||||
| Hallux | male | 29.27 | 1.72 | 29.52 | 24.70 | 33.73 | <0.001 | 0.476 | −6.77 |
| female | 31.40 | 2.48 | 31.67 | 25.12 | 36.96 | ||||
| Bill length | male | 24.32 | 1.79 | 24.50 | 20.58 | 28.99 | 0.001 | 0.397 | −6.21 |
| female | 25.93 | 2.08 | 25.96 | 21.42 | 31.12 | ||||
| Bill height | male | 19.23 | 1.22 | 19.30 | 16.53 | 22.40 | <0.001 | 0.543 | −6.53 |
| female | 20.58 | 0.93 | 20.68 | 18.06 | 22.53 | ||||
| Bill width | male | 13.68 | 1.32 | 13.86 | 10.79 | 16.84 | 0.002 | 0.356 | −6.73 |
| female | 14.66 | 1.26 | 14.55 | 11.75 | 17.76 | ||||
| Head length | male | 87.02 | 5.04 | 87.15 | 76.14 | 99.47 | 0.003 | 0.335 | −4.24 |
| female | 90.87 | 6.38 | 90.76 | 75.93 | 105.71 | ||||
| Head width | male | 43.48 | 2.56 | 43.46 | 38.42 | 48.54 | 0.008 | 0.295 | −4.72 |
| female | 45.64 | 4.16 | 45.23 | 36.14 | 58.54 | ||||
| Forearm length | male | 161.41 | 13.92 | 165.50 | 121.00 | 184.00 | 0.032 | 0.225 | −4.80 |
| female | 169.56 | 17.57 | 170.00 | 128.00 | 210.00 | ||||
| Seventh primary | male | 145.47 | 41.06 | 149.50 | 68.00 | 250.00 | 0.352 | 0.047 | −3.77 |
| female | 151.18 | 36.22 | 146.50 | 79.00 | 247.00 | ||||
| Rectrix | male | 105.41 | 33.99 | 105.00 | 21.00 | 205.00 | 0.527 | 0.001 | −1.73 |
| female | 107.27 | 30.13 | 104.50 | 46.00 | 188.00 | ||||
| Body length | male | 436.87 | 52.08 | 440.00 | 325.00 | 580.00 | 0.024 | 0.240 | −5.03 |
| female | 460.03 | 47.68 | 465.00 | 349.00 | 585.00 | ||||
| Body mass | male | 1359.0 | 171.0 | 1387.50 | 920 | 1625 | <0.001 | 0.597 | −18.27 |
| female | 1662.9 | 254.1 | 1725.00 | 1015 | 2180 |
| Variable | Sex | Mean | sd | Median | Min | Max | p-Value | Effect Size | % Dimorphism |
|---|---|---|---|---|---|---|---|---|---|
| Lateral tarsus | male | 10.70 | 0.75 | 10.40 | 9.70 | 11.80 | <0.001 | 0.769 | −12.70 |
| female | 12.26 | 0.57 | 12.35 | 11.40 | 13.12 | ||||
| Dorso-ventral tarsus | male | 12.68 | 0.58 | 12.80 | 11.80 | 13.60 | <0.001 | 0.844 | −12.17 |
| female | 14.44 | 0.57 | 14.50 | 13.44 | 15.20 | ||||
| Hallux | male | 38.59 | 1.39 | 38.70 | 36.30 | 41.90 | <0.001 | 0.771 | −9.90 |
| female | 42.82 | 1.79 | 42.85 | 38.90 | 46.00 | ||||
| Bill length | male | 34.41 | 5.21 | 32.12 | 29.60 | 44.70 | 0.002 | 0.514 | −10.97 |
| female | 38.65 | 6.29 | 35.05 | 32.45 | 48.30 | ||||
| Bill height | male | 22.69 | 0.63 | 22.80 | 21.60 | 23.80 | <0.001 | 0.738 | −6.01 |
| female | 24.15 | 0.68 | 24.05 | 22.80 | 25.20 | ||||
| Bill width | male | 16.97 | 0.96 | 17.00 | 15.50 | 19.00 | 0.002 | 0.505 | −7.68 |
| female | 18.38 | 1.36 | 18.70 | 15.90 | 20.10 | ||||
| Head length | male | 98.93 | 4.89 | 99.50 | 84.80 | 107.60 | <0.001 | 0.677 | −5.54 |
| female | 104.72 | 2.51 | 104.15 | 100.80 | 109.50 | ||||
| Head width | male | 52.07 | 2.90 | 52.00 | 47.00 | 56.80 | 0.004 | 0.464 | −4.95 |
| female | 54.79 | 2.13 | 54.55 | 51.90 | 58.90 | ||||
| Forearm length | male | 179.47 | 11.34 | 181.00 | 164.00 | 200.00 | 0.017 | 0.373 | −5.94 |
| female | 190.81 | 12.37 | 190.50 | 175.00 | 212.00 | ||||
| Seventh primary | male | 33.15 | 3.34 | 34.00 | 24.90 | 39.20 | 0.003 | 0.484 | −7.64 |
| female | 35.89 | 2.23 | 36.75 | 31.10 | 39.30 | ||||
| Rectrix | male | 250.53 | 14.37 | 250.00 | 219.00 | 275.00 | <0.001 | 0.587 | −7.43 |
| female | 270.63 | 13.73 | 271.50 | 242.00 | 295.00 | ||||
| Body length | male | 634.94 | 17.06 | 638.00 | 600.00 | 665.00 | <0.001 | 0.773 | −7.38 |
| female | 685.56 | 22.75 | 693.00 | 641.00 | 710.00 | ||||
| Wingspan | male | 151.18 | 7.88 | 152.00 | 138.00 | 162.00 | <0.001 | 0.694 | −9.03 |
| female | 166.19 | 7.34 | 169.50 | 152.00 | 174.00 | ||||
| Body mass | male | 1882.2 | 115.2 | 1894.00 | 1700 | 2060 | <0.001 | 0.851 | −24.65 |
| female | 2498.1 | 182.9 | 2490.00 | 2160 | 2900 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estellés-Domingo, I.; López-López, P. Non-Invasive Sex Determination of Nestlings and Adult Bonelli’s Eagles Using Morphometrics. Animals 2023, 13, 1201. https://doi.org/10.3390/ani13071201
Estellés-Domingo I, López-López P. Non-Invasive Sex Determination of Nestlings and Adult Bonelli’s Eagles Using Morphometrics. Animals. 2023; 13(7):1201. https://doi.org/10.3390/ani13071201
Chicago/Turabian StyleEstellés-Domingo, Irene, and Pascual López-López. 2023. "Non-Invasive Sex Determination of Nestlings and Adult Bonelli’s Eagles Using Morphometrics" Animals 13, no. 7: 1201. https://doi.org/10.3390/ani13071201