


Glycerol-Free Equilibration with the Addition of Glycerol Shortly before the Freezing Procedure: A Perspective Strategy for Cryopreservation of Wallachian Ram Sperm
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Solutions
2.2. Ethical Statements
2.3. Animal Management
2.4. Semen Collection
2.5. Semen Processing and Equilibration
2.6. Sperm Freezing and Thawing
2.7. Sperm Motility Evaluation by iSperm® mCASA
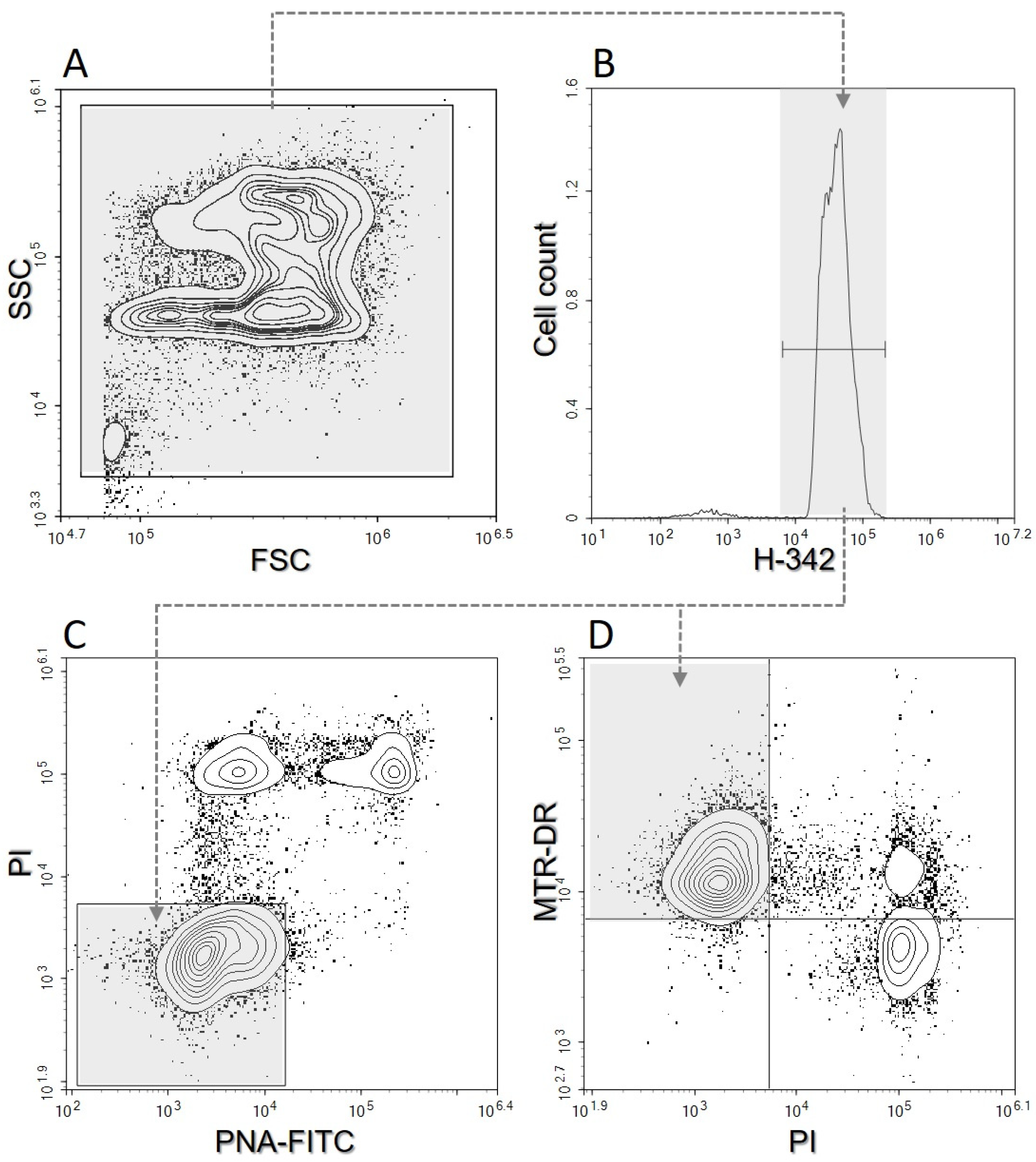
2.8. Flow Cytometry Assessment
2.9. Statistical Analysis
- Yijk = µ + DAYi + VARj + eijk (for equilibrated sperm cells only);
- Yijk = µ + DAYi + VARj + b1−7 × EQ+ eijk (for cryopreserved sperm cells only).
- Yijk = µ + DAYi + VARj + DAY × VARij + eijk (for equilibrated sperm cells only);
- Yijk = µ + DAYi + VARj + DAY × VARij + b1−2×EQ + eijk (for cryopreserved sperm cells only).
3. Results
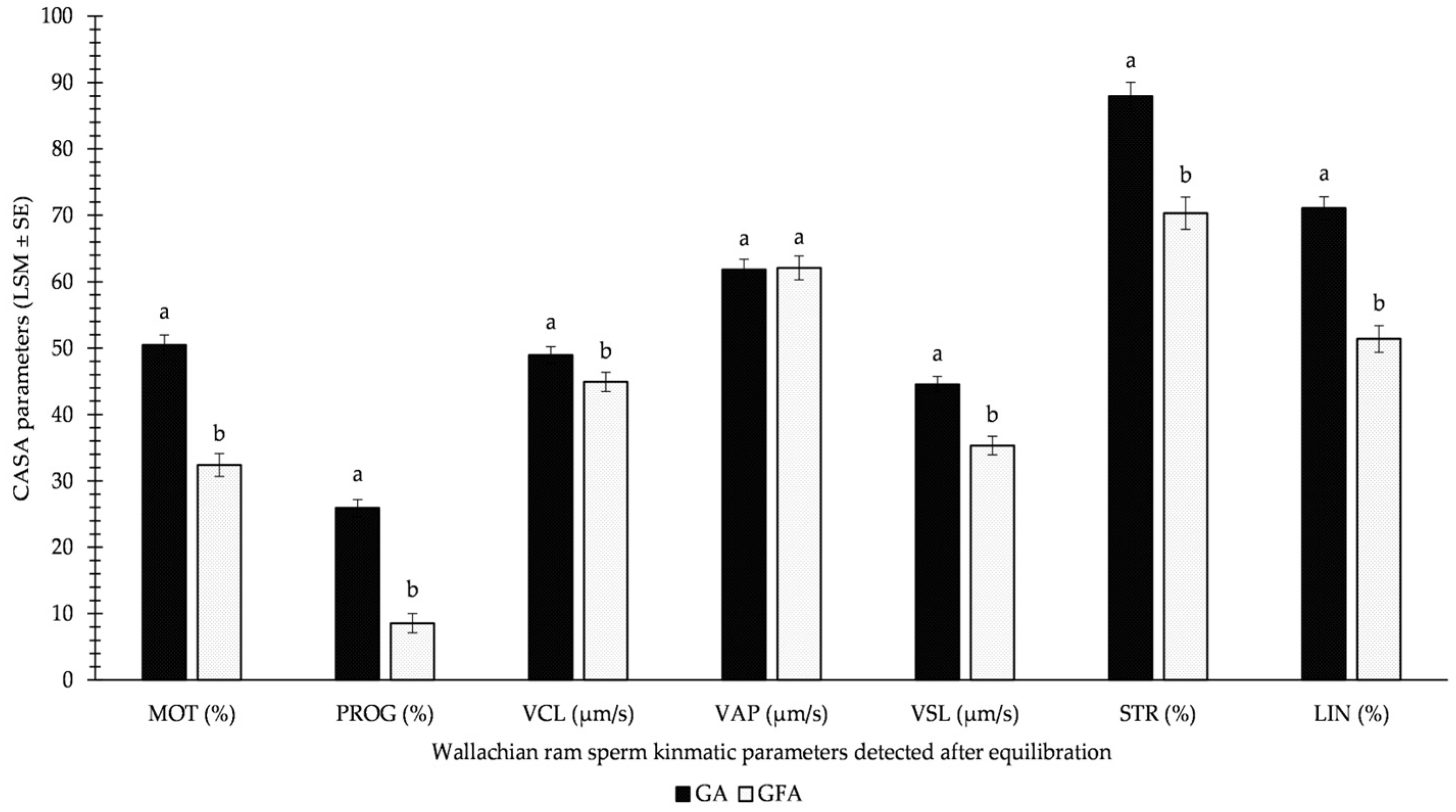
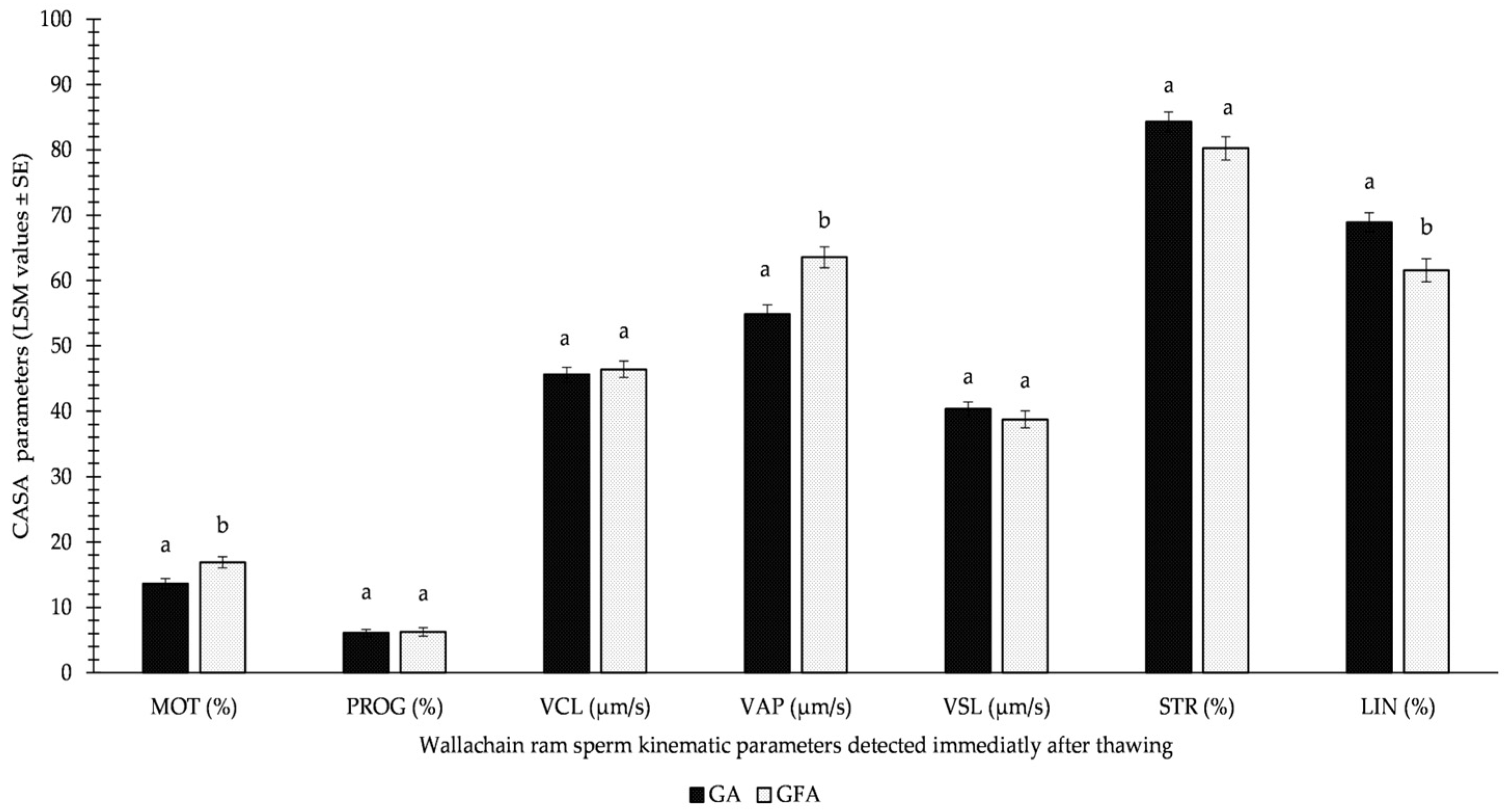
3.1. CASA Parameters
3.2. Flow Cytometry Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Machová, K.; Milerski, M.; Rychtářová, J.; Hofmanová, B.; Vostrá-Vydrová, H.; Moravčíková, N.; Kasarda, R.; Vostrý, L. Assessment of the genetic diversity of Two Czech autochthonous sheep breeds. Small Rumin. Res. 2021, 195, 106301. [Google Scholar] [CrossRef]
- Sonesson, A.K.; Goddard, M.E.; Meuwissen, T.H.E. The use of frozen semen to minimize inbreeding in small populations. Genet. Res. 2002, 80, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Savvulidi, F.G.; Ptacek, M.; Malkova, A.; Beranek, J.; Stadnik, L. Optimizing the conventional method of sperm freezing in liquid nitrogen vapour for Wallachian sheep conservation program. Czech J. Anim. Sci. 2021, 66, 55–64. [Google Scholar] [CrossRef]
- Vozaf, J.; Makarevich, A.V.; Balazi, A.; Vasicek, J.; Svoradova, A.; Olexikova, L.; Chrenek, P. Cryopreservation of ram semen: Manual versus programmable freezing and different lengths of equilibration. Anim. Sci. J. 2021, 92, e13670. [Google Scholar] [CrossRef] [PubMed]
- Keskin, N.; Erdogan, C.; Bucak, M.N.; Ozturk, A.E.; Bodu, M.; Ili, P.; Baspinar, N.; Dursun, S. Cryopreservation Effects on Ram Sperm Ultrastructure. Biopreserv. Biobank. 2020, 18, 441–448. [Google Scholar] [CrossRef]
- Raju, R.; Bryant, S.J.; Wilkinson, B.L.; Bryant, G. The need for novel cryoprotectants and cryopreservation protocols: Insights into the importance of biophysical investigation and cell permeability. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129749. [Google Scholar] [CrossRef]
- Salamon, S.; Maxwell, W.M. Storage of ram semen. Anim. Reprod. Sci. 2000, 62, 77–111. [Google Scholar] [CrossRef]
- Nur, Z.; Zik, B.; Ustuner, B.; Sagirkaya, H.; Ozguden, C.G. Effects of different cryoprotective agents on ram sperm morphology and DNAintegrity. Theriogenology 2010, 73, 1267–1275. [Google Scholar] [CrossRef]
- Curry, M.R.; Watson, P.F. Osmotic effects on ram and human sperm membranes in relation to thawing injury. Cryobiology 1994, 31, 39–46. [Google Scholar] [CrossRef]
- Holt, W.V.; North, R.D. Effects of temperature and restoration of osmotic equilibrium during thawing on the induction of plasma membrane damage in cryopreserved ram spermatozoa. Biol. Reprod. 1994, 51, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Guthrie, H.D.; Liu, J.; Critser, J.K. Osmotic tolerance limits and effects of cryoprotectants on motility of bovine spermatozoa. Biol. Reprod. 2002, 67, 1811–1816. [Google Scholar] [CrossRef] [Green Version]
- Barrios, B.; Pérez-Pé, R.; Gallego, M.; Tato, A.; Osada, J.; Muiño-Blanco, T.; Cebrián-Pérez, J.A. Seminal plasma proteins revert the cold-shock damage on ram sperm membrane. Biol. Reprod. 2000, 63, 1531–1537. [Google Scholar] [CrossRef] [Green Version]
- Fiser, P.S.; Fairfull, R.W. The effect of glycerol concentration and cooling velocity on cryosurvival of ram spermatozoa frozen in straws. Cryobiology 1984, 21, 542–551. [Google Scholar] [CrossRef]
- Álvarez, M.; Tamayo-Canul, J.; Martínez-Rodríguez, C.; López-Urueña, E.; Gomes-Alves, S.; Anel, L.; Martínez-Pastor, F.; de Paz, P. Specificity of the extender used for freezing ram sperm depends of the spermatozoa source (ejaculate, electroejaculate or epididymis). Anim. Reprod. Sci. 2012, 132, 145–154. [Google Scholar] [CrossRef]
- De Paz, P.; Alvarez-Rodriguez, M.; Nicolas, M.; Alvarez, M.; Chamorro, C.; Borragán, S.; Martinez-Pastor, F.; Anel, L. Optimization of glycerol concentration and freezing rate in the cryopreservation of ejaculate from brown bear (Ursus arctos). Reprod. Domest. Anim. 2012, 47, 105–112. [Google Scholar] [CrossRef]
- Colas, G. Effect of initial freezing temperature, addition of glycerol and dilution on the survival and fertilizing ability of deep-frozen ram semen. J. Reprod. Fertil. 1975, 42, 277–285. [Google Scholar] [CrossRef]
- Alvarez, M.; Tamayo-Canul, J.; Anel, E.; Boixo, J.C.; Mata-Campuzano, M.; Martinez-Pastor, F.; Anel, L.; de Paz, P. Sperm concentration at freezing affects post-thaw quality and fertility of ram semen. Theriogenology 2012, 77, 1111–1118. [Google Scholar] [CrossRef]
- Mehta, V.; Kumar Pareek, P.; Kumar, A.; Narayan Purohit, G. Comparative Effect of Different Concentrations of Glycerol and Ethylene Glycol and Temperature on Cryopreservation of Ram Semen. RJVP 2020, 8, 37–41. [Google Scholar] [CrossRef]
- Salamon, S. Deep freezing of ram semen: Recovery of spermatozoa after pelleting and comparison with other methods of freezing. Aust. J. Biol. Sci. 1968, 21, 351–360. [Google Scholar] [CrossRef]
- Berndtson, W.E.; Foote, R.H. The freezability of spermatozoa after minimal pre-freezing exposure to glycerol or lactose. Cryobiology 1972, 9, 57–60. [Google Scholar] [CrossRef]
- Gao, D.Y.; Ashworth, E.; Watson, P.F.; Kleinhans, F.W.; Mazur, P.; Critser, J.K. Hyperosmotic tolerance of human spermatozoa: Separate effects of glycerol, sodium chloride, and sucrose on spermolysis. Biol. Reprod. 1993, 49, 112–123. [Google Scholar] [CrossRef]
- Gao, D.Y.; Liu, J.; Liu, C.; McGann, L.E.; Watson, P.F.; Kleinhans, F.W.; Mazur, P.; Critser, E.S.; Critser, J.K. Prevention of osmotic injury to human spermatozoa during addition and removal of glycerol. Hum. Reprod. 1995, 10, 1109–1122. [Google Scholar] [CrossRef]
- Akçay, E.; Kulaksız, R.; Daşkin, A.; Çebi, Ç.; Tekin, K. The effect of different dilution rates on post-thaw quality of ram semen frozen in two different egg-yolk free extenders. Slov. Vet. Res. 2012, 49, 97–102. [Google Scholar]
- Russel, A.J.F.; Doney, J.M.; Gunn, R.G. Subjective assessment of body fat in live sheep. J. Agric. Sci. 1969, 72, 451–454. [Google Scholar] [CrossRef]
- Fiser, P.S.; Batra, T.R. Effect of equilibration time at 5 °C and photoperiod on survival of ram spermatozoa frozen in straws. Can. J. Anim. Sci. 1984, 64, 777–780. [Google Scholar] [CrossRef]
- Ptáček, M.; Stádníková, M.; Savvulidi, F.; Stádník, L. Ram Semen Cryopreservation Using Egg Yolk or Egg Yolk-free Extenders: Preliminary Results. Sci. Agric. Bohem. 2019, 50, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, N.; Casao, A.; Domingo, J.; Quintín, F.; Laviña, A.; Fantova, E.; Cebrián-Pérez, J.Á.; Muiño-Blanco, T.; Pérez-Pe, R. Influence of Non-conventional Sperm Quality Parameters on Field Fertility in Ovine. Front. Vet. Sci. 2021, 8, 650572. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pastor, F.; Mata-Campuzano, M.; Alvarez-Rodríguez, M.; Alvarez, M.; Anel, L.; de Paz, P. Probes and techniques for sperm evaluation by flow cytometry. Reprod. Domest. Anim. 2010, 45 (Suppl. S2), 67–78. [Google Scholar] [CrossRef]
- Berndtson, W.E.; Foote, R.H. The survival of frozen bovine spermatozoa following minimum exposure to glycerol. Cryobiology 1969, 5, 398–402. [Google Scholar] [CrossRef]
- Sharafi, M.; Borghei-Rad, S.M.; Hezavehei, M.; Shahverdi, A.; Benson, J.D. Cryopreservation of Semen in Domestic Animals: A Review of Current Challenges, Applications, and Prospective Strategies. Animals 2022, 12, 3271. [Google Scholar] [CrossRef]
- Slavík, T. Effect of glycerol on the penetrating ability of fresh ram spermatozoa with zona-free hamster eggs. J. Reprod. Fertil. 1987, 79, 99–103. [Google Scholar] [CrossRef]
- Purdy, P.H. The post-thaw quality of ram sperm held for 0 to 48 h at 5 degrees C prior to cryopreservation. Anim. Reprod. Sci. 2006, 93, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Câmara, D.R.; Silva, S.V.; Almeida, F.C.; Nunes, J.F.; Guerra, M.M.P. Effects of antioxidants and duration of pre-freezing equilibration on frozen-thawed ram semen. Theriogenology 2011, 76, 342–350. [Google Scholar] [CrossRef]
- Jones, R.C. The use of dimethyl sulphoxide, glycerol, and reconstituted skim milk for the preservation of ram spermatozoa. II. The influence of diluent composition and processing time during freezing to minus 79 degrees C with dimethyl sulphoxide or glycerol or both compounds. Aust. J. Biol. Sci. 1965, 18, 887–900. [Google Scholar] [CrossRef] [Green Version]
- Jang, T.H.; Park, S.C.; Yang, J.H.; Kim, J.Y.; Seok, J.H.; Park, U.S.; Choi, C.W.; Lee, S.R.; Han, J. Cryopreservation and its clinical applications. Integr. Med. Res. 2017, 6, 12–18. [Google Scholar] [CrossRef]
- Delgado-Bermúdez, A.; Ribas-Maynou, J.; Yeste, M. Relevance of Aquaporins for Gamete Function and Cryopreservation. Animals 2022, 12, 573. [Google Scholar] [CrossRef]
- Mohri, H.; Masaki, J. Glycerokinase and its possible role in glycerol metabolism of bull spermatozoa. J. Reprod. Fertil. 1967, 14, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Papa, P.M. Effect of glycerol on the viability and fertility of cooled bovine semen. Theriogenology 2015, 83, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Savvulidi, F.G.; Ptacek, M.; Malkova, A.; Kratochvilova, I.; Simek, D.; Martinez-Pastor, F.; Stadnik, L. Inhibition of extracellular ice crystals growth for testing the cryodamaging effect of intracellular ice in a model of ram sperm ultra-rapid freezing. J. Appl. Anim. Res. 2023, 51, 182–192. [Google Scholar] [CrossRef]
- Rall, W.F.; Mazur, P.; Souzu, H. Physical-chemical basis of the protection of slowly frozen human erythrocytes by glycerol. Biophys. J. 1978, 23, 101–120. [Google Scholar] [CrossRef] [Green Version]
- Katkov, I.I.; Katkova, N.; Critser, J.K.; Mazur, P. Mouse spermatozoa in high concentrations of glycerol: Chemical toxicity vs. osmotic shock at normal and reduced oxygen concentrations. Cryobiology 1998, 37, 325–338. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| b1–7 × EQ | DAY | VAR | |
|---|---|---|---|
| MOTEQ | --- | *** | *** |
| PROGEQ | --- | *** | *** |
| VCLEQ | --- | *** | * |
| VAPEQ | --- | *** | n.s. |
| VSLEQ | --- | *** | *** |
| STREQ | --- | *** | *** |
| LINEQ | --- | *** | *** |
| MOTT0 | ** | *** | ** |
| PROGT0 | * | *** | n.s. |
| VCLT0 | *** | *** | n.s. |
| VAPT0 | *** | *** | *** |
| VSLT0 | *** | *** | n.s. |
| STRT0 | *** | * | n.s. |
| LINT0 | *** | ** | *** |
| GA (%) | GFA (%) | Significance of Factors in Statistical Model | ||||
|---|---|---|---|---|---|---|
| b1–2 × EQ | DAY | VAR | DAY × VAR | |||
| PAIEQ | 60.48 ± 1.61 a | 36.99 ± 1.61 b | --- | *** | *** | *** |
| PAIT0 | 22.38 ± 0.87 a | 27.12 ± 0.82 b | *** | *** | *** | *** |
| PAIT1 | 10.92 ± 0.74 a | 22.55 ± 0.69 b | *** | *** | *** | *** |
| PAIT2 | 9.61 ± 0.68 a | 20.53 ± 0.63 b | *** | *** | *** | *** |
| MMPEQ | 64.20 ± 1.75 a | 54.92 ± 1.75 b | --- | *** | *** | *** |
| MMPT0 | 36.67 ± 1.40 a | 41.03 ± 1.30 b | *** | *** | * | *** |
| MMPT1 | 7.16 ± 0.82 a | 18.97 ± 0.77 b | * | *** | *** | *** |
| MMPT2 | 7.49 ± 0.75 a | 11.72 ± 0.71 b | *** | *** | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Málková, A.; Savvulidi, F.G.; Ptáček, M.; Machová, K.; Janošíková, M.; Nagy, S.; Stádník, L. Glycerol-Free Equilibration with the Addition of Glycerol Shortly before the Freezing Procedure: A Perspective Strategy for Cryopreservation of Wallachian Ram Sperm. Animals 2023, 13, 1200. https://doi.org/10.3390/ani13071200
Málková A, Savvulidi FG, Ptáček M, Machová K, Janošíková M, Nagy S, Stádník L. Glycerol-Free Equilibration with the Addition of Glycerol Shortly before the Freezing Procedure: A Perspective Strategy for Cryopreservation of Wallachian Ram Sperm. Animals. 2023; 13(7):1200. https://doi.org/10.3390/ani13071200
Chicago/Turabian StyleMálková, Anežka, Filipp Georgijevič Savvulidi, Martin Ptáček, Karolína Machová, Martina Janošíková, Szabolcs Nagy, and Luděk Stádník. 2023. "Glycerol-Free Equilibration with the Addition of Glycerol Shortly before the Freezing Procedure: A Perspective Strategy for Cryopreservation of Wallachian Ram Sperm" Animals 13, no. 7: 1200. https://doi.org/10.3390/ani13071200