
Non-Invasive Wildlife Disease Surveillance Using Real Time PCR Assays: The Case of the Endangered Galemys pyrenaicus Populations from the Central System Mountains (Extremadura, Spain)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
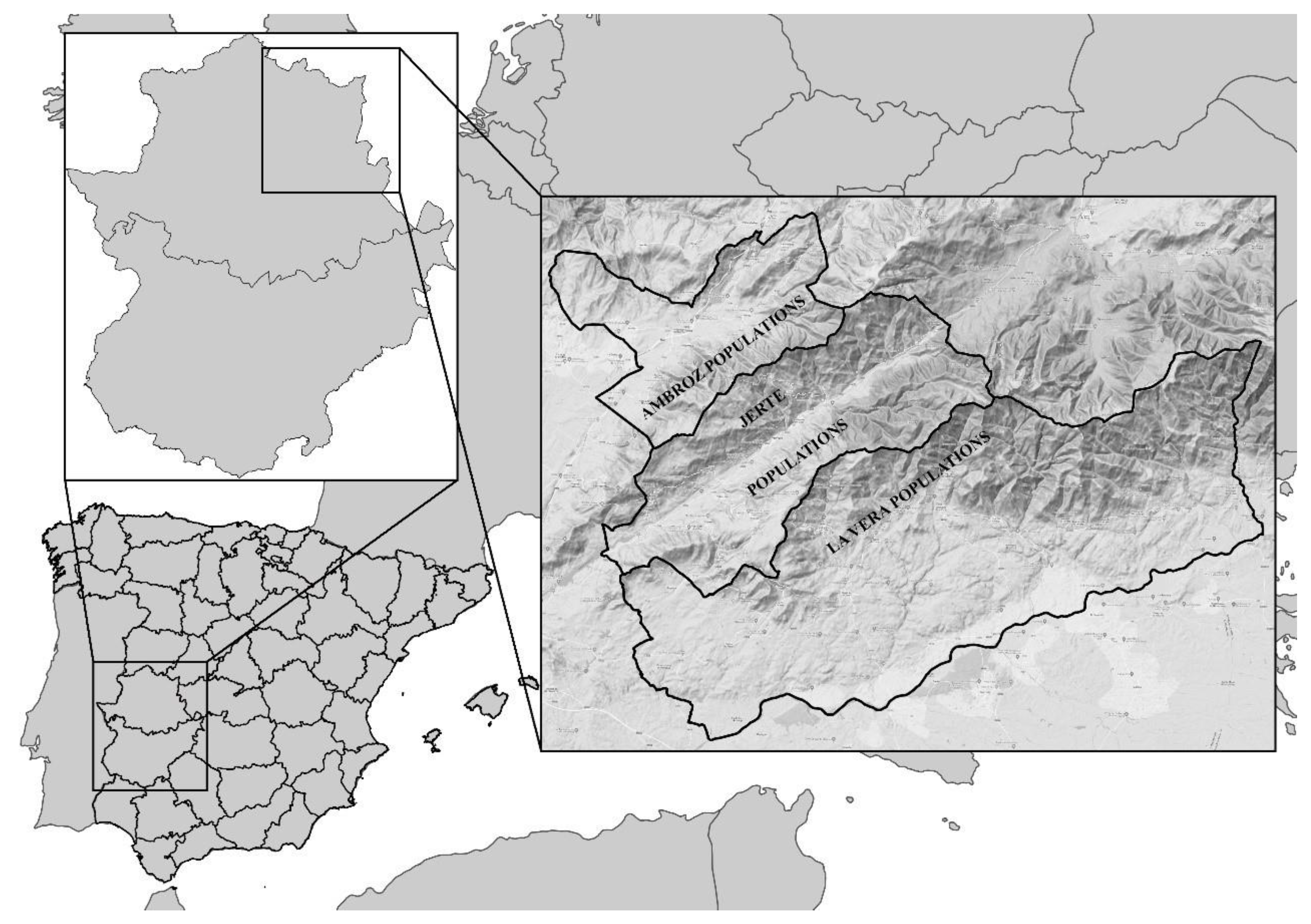
2.1. Study Area and Sampling
2.2. DNA Extraction
2.3. Sample Genotyping
2.4. Hydrolysis Probe Assay, End Point PCR and Design
2.4.1. Real-Time PCR Using Hydrolysis Probes
2.4.2. End Point and Real-Time PCR using Hydrolysis Probes to Detect Omphalometridae
2.5. Statistical Analysis
3. Results
3.1. Sampling Distribution According to the Recuperation Plan of the Desman in Extremadura
3.2. Bacteria and Parasites Study
3.2.1. Remarks on Microbes
3.2.2. Remarks on Parasitic Infections
3.2.3. Prevalence in the Entire Dataset
3.3. Ompahlometra Findings
4. Discussion
4.1. The First Overview of the Health Status of the Most Endangered Desman Populations: Central System
4.2. New Report of Omphalometra spp. in Desman
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernández-García, J.L.; Cedillo, M.d.P.V. Faecal DNA template as non-invasive tools in order to distinguish the endangered Pyrenean desman (Galemys pyrenaicus, Eulipotyphla, Talpidae) from Mediterranean water shrews (Neomys anomalus, Soricomorpha, Soricidae). Hystrix. Ital. J. Mamm. 2017, 28, 92–97. [Google Scholar] [CrossRef]
- Quaglietta, L. Galemys Pyrenaicus (Versión Modificada de la Evaluación de 2021). La Lista Roja de Especies Amenazadas de la UICN 2022 2021. Available online: www.iucnredlist.org (accessed on 9 December 2021).
- Mélanie, N.; Alain, B.; Mallorie, S.; Pascal, A. A French Conservation Action Plan for the Pyrenean Desman Galemys pyrenai-cus. Galemys Boletín Inf. Soc. Española Para Conserv. Estud. Mamíferos 2011, 23, 47–50. [Google Scholar]
- Quaglietta, L.; Paupério, J.; Martins, F.M.S.; Alves, P.C.; Beja, P. Recent range contractions in the globally threatened Pyrenean desman highlight the importance of stream headwater refugia. Anim. Conserv. 2018, 21, 515–525. [Google Scholar] [CrossRef]
- Gisbert, J.; García-Perea, R. Historia de la regresión del desmán ibérico Galemys pyrenaicus (É. Geoffroy Saint-Hilaire, 1811) en el Sistema Central (Península Ibérica). Munibe Monogr. Nat. Ser. 2014, 3, 19–35. [Google Scholar] [CrossRef]
- Escoda, L.; Castresana, J. The genome of the Pyrenean desman and the effects of bottlenecks and inbreeding on the genomic landscape of an endangered species. Evol. Appl. 2021, 14, 1898–1913. [Google Scholar] [CrossRef]
- Charbonnel, A.; Laffaille, P.; Biffi, M.; Blanc, F.; Maire, A.; Némoz, M.; Perez, J.M.S.; Sauvage, S.; Buisson, L. Can Recent Global Changes Explain the Dramatic Range Contraction of an Endangered Semi-Aquatic Mammal Species in the French Pyrenees? PLoS ONE 2016, 11, e0159941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meerburg, B.G.; Jacobs-Reitsma, W.F.; Wagenaar, J.A.; Kijlstra, A. Presence of Salmonella and Campylobacter spp. in Wild Small Mammals on Organic Farms. Appl. Environ. Microbiol. 2006, 72, 960–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, D.W.; Paterson, R.A.; Townsend, C.R.; Poulin, R.; Tompkins, D.M. Parasite spillback: A neglected concept in invasion ecology? Ecology 2009, 90, 2047–2056. [Google Scholar] [CrossRef]
- Mangombi, J.B.; N’Dilimabaka, N.; Lekana-Douki, J.-B.; Banga, O.; Maghendji-Nzondo, S.; Bourgarel, M.; Leroy, E.; Fenollar, F.; Mediannikov, O. First investigation of pathogenic bacteria, protozoa and viruses in rodents and shrews in context of forest-savannah-urban areas interface in the city of Franceville (Gabon). PLoS ONE 2021, 16, e0248244. [Google Scholar] [CrossRef]
- Singla, L.D.; Singla, N.; Parshad, V.R.; Juyal, P.D.; Sood, N.K. Rodents as reservoirs of parasites in India. Integr. Zool. 2008, 3, 21–26. [Google Scholar] [CrossRef]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 6783, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J. What is rarity? In Rarity; Springer Science and Business Media: Comwall, UK, 1994. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, A.; de Carvalho, L.; González, M.; Torres, R.; Pla, S.; Núñez-Arjona, J.; Rueda, C.; Vallverdú-Coll, N.; Silvestre, F.; Peña, J.; et al. Parasites of the Reintroduced Iberian Lynx (Lynx pardinus) and Sympatric Mesocarnivores in Extremadura, Spain. Pathogens 2021, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Ryser-Degiorgis, M.-P. Wildlife health investigations: Needs, challenges and recommendations. BMC Veter.-Res. 2013, 9, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timon-David, J. Un trématode parasite du Desman des Pyrénées (Galemys pyrenaicus Geoffr.). Bull. Société D’histoire Nat. Toulouse 1960, 95, 11–17. [Google Scholar]
- Vaucher, C. Trématodes parasites de Chiroptères et d’Insectivores. Bull. Soc. Neuchâtel. Sci. Nat. 1975, 98, 17–25. [Google Scholar]
- Deblock, S.; Combes, C. Contribution à l’étude des Microphallidae, Tranassos, 1920 (Trema- tada). X. Maritrema pyrenaica n. sp., parasite du Desman (Mammifère Insectivore). Essai de clé diagnostique des espèces du genre. Bull. Soc. Zool. Fr. 1965, 90, 101–117. [Google Scholar]
- Richard, J.; Jourdane, J. Chetotaxie des cercaires de deux Trematodes Microphallidés Microphallus gracilis (Baer,1943) et Maritrema pyrenaica (Deblock et Combes, 1965), parasites de Neomys fodiens (author´s transl). Ann. Parasitol. Hum. Comp. 1978, 6, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Jourdane, J. The life cycl of Maritrema pyrenaica Deblock et Combes, 1965, a parasite of insectivorous micromammals in the Pyrences (author’s transl). Ann. Parasitol. Hum. Comparée 1979, 54, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Casanova, J.C.; Villa, M.; Montoliu, I. First record of Maritrema pyrenaica (Digenea: Microphallidae) in Spain (Western Pyre-nees) in its intermediate hosts. Folia Parasitol. 1998, 45, 251–252. [Google Scholar]
- Mas-Coma, S.; Roset, F.; Mantoliu, I. Matovius galemydis sp. n.(Trematoda: Lecithodendrioidea), a parasite of Galemys pyre-naicus (Geoffroy, 1811)(Insectivora: Talpidae) in Spain. Proposal of Combesia gen. n. and Matoviinae subfam. Helminthologia 1985, 22, 87–89. [Google Scholar]
- Mascato, M.F.A. Contribución al estudio de la helmintofauna de mamíferos en la Comunidad Autónoma Gallega. Ph.D. Thesis, Universidade de Santiago de Compostela, Galicia, Spain, 1990. [Google Scholar]
- Alvarez, F.; Gijón-Botella, H.; Quinteiro, P.; Rey, J.; López-Román, F.; Sanmartiín, M.L. Paracuaria hispanica n. sp. (Nematoda: Acuariidae), a stomach parasite of the pyrenean desman Galemys pyrenaicus Geoffr. (Insectivora: Talpidae), with a redefinition of the genus Paracuaria Rao, 1951. Syst. Parasitol. 1994, 29, 105–112. [Google Scholar] [CrossRef]
- Diario Oficial de Extremadura. ORDEN de 3 de agosto de 2018 por la que se aprueba el Plan de Recuperación del Desmán Ibérico (Galemys pyrenaicus) en Extremadura. Extremadura 2018, 158, 32143–32165. [Google Scholar]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Vitullo, M.; Grant, K.A.; Sammarco, M.L.; Tamburro, M.; Ripabelli, G.; Amar, C.F.L. Real-time PCRs assay for serogrouping Listeria monocytogenes and differentiation from other Listeria spp. Mol. Cell. Probes 2012, 27, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Tkach, V.; Grabda-Kazubska, B.; Swiderski, Z. Systematic position and phylogenetic relationships of the family Omphalometridae (Digenea, Plagiorchiida) inferred from partial lsrDNA sequences. Int. J. Parasitol. 2001, 31, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Gillet, F.; Tiouchichine, M.-L.; Galan, M.; Blanc, F.; Némoz, M.; Aulagnier, S.; Michaux, J.R. A new method to identify the endangered Pyrenean desman (Galemys pyrenaicus) and to study its diet, using next generation sequencing from faeces. Mamm. Biol. 2015, 80, 505–509. [Google Scholar] [CrossRef]
- Alonso-Andicoberry, C.; García-Peña, F.; Pereira-Bueno, J.; Costas, E.; Ortega-Mora, L. Herd-level risk factors associated with Leptospira spp. seroprevalence in dairy and beef cattle in Spain. Prev. Veter.-Med. 2001, 52, 109–117. [Google Scholar] [CrossRef]
- Rudolphi, C.A. Entozoorum, Sive Vermium Intestinalium, Historia Naturalis. Amstelaedami; Harvard University, Museum of Comparative Zoology, Ernst Mayr Library: Cambridge, MA, USA, 1809; Volume 2. [Google Scholar]
- Mühling, P. Beiträge zur Kenntnis der Trematoden. Arch. F. Naturg. 1896, 62, 243–279. [Google Scholar]
- Looss, A. Weitere BeitraÈge zur Kenntnis der Trematodenfauna AÈ gyptens Zugleich Versuche einer natuÈrlichen Gliederung des Genus Distomum Reyzius. Zool. Jb. Abt. Syst. 1899, 12, 521–784. [Google Scholar]
- Odening, K. Das Exkretionssystem von Omphalometra und Brachycoelium (Trematoda, Digenea) und die Taxonomie der Unterordnung Plagiorchiata. Z. Für Parasitenkd. 1959, 19, 442–457. [Google Scholar] [CrossRef]
- Greiman, S.E.; Cook, J.A.; Tkach, V.V.; Hoberg, E.P.; Menning, D.; Hope, A.G.; Sonsthagen, S.A.; Talbot, S.L. Museum metabarcoding: A novel method revealing gut helminth communities of small mammals across space and time. Int. J. Parasitol. 2018, 48, 1061–1070. [Google Scholar] [CrossRef] [Green Version]
- Ribas, A.; Feliu, C.; Fons, R.; Casanova, J.C. Larval Digneans of Mammals in Freshwater Invertebrates as Intermediate Hosts. 2009. Available online: https://hal.sorbonne-universite.fr/hal-03253707 (accessed on 9 December 2021).
- Tkach, V.V. Maritrema neomi n. sp. (Digenea: Microphallidae) from water shrews (Neomys). J. Parasitol. 1998, 84, 846. [Google Scholar] [CrossRef]
- Tkach, V.V.; Littlewood, D.T.J.; Olson, P.D.; Kinsella, J.M.; Swiderski, Z. Molecular phylogenetic analysis of the Microphalloidea Ward, 1901 (Trematoda: Digenea). Syst. Parasitol. 2003, 56, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as Hosts of Pathogens and Related Zoonotic Disease Risk. Pathogens 2020, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Diagne, C.; Galan, M.; Tamisier, L.; D’Ambrosio, J.; Dalecky, A.; Bâ, K.; Kane, M.; Niang, Y.; Diallo, M.; Sow, A.; et al. Ecological and sanitary impacts of bacterial communities associated to biological invasions in African commensal rodent communities. Sci. Rep. 2017, 7, 14995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boundenga, L.; Ngoubangoye, B.; Ntie, S.; Moukodoum, N.-D.; Renaud, F.; Rougeron, V.; Prugnolle, F. Rodent malaria in Gabon: Diversity and host range. Int. J. Parasitol. Parasites Wildl. 2019, 10, 117–124. [Google Scholar] [CrossRef]
- Heuzenroeder, M. Salmonella in Domestic Animals. Aust. Veter.-J. 2000, 78, 570. [Google Scholar] [CrossRef]
- Millan, J.; Aduriz, G.; Moreno, B.; Juste, R.; Barral, M. Salmonella isolates from wild birds and mammals in the Basque Country (Spain). OIE Rev. Sci. Et Tech. 2004, 23, 905–911. [Google Scholar] [CrossRef]
- Obiegala, A.; Woll, D.; Karnath, C.; Silaghi, C.; Schex, S.; Eßbauer, S.; Pfeffer, M. Prevalence and Genotype Allocation of Pathogenic Leptospira Species in Small Mammals from Various Habitat Types in Germany. PLOS Neglected Trop. Dis. 2016, 10, e0004501. [Google Scholar] [CrossRef] [Green Version]
- Ospina-Pinto, C.; Rincon-Pardo, M.; Soler-Tovar, D.; Hernández-Rodríguez, P. Papel de los roedores en la transmisión de Leptospira spp. en granjas porcinas. Rev. Salud Pública 2017, 19, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, C.J.; Gerbig, G.R.; Sensius, L.D.; Patel, V.; Smith, T.C. Staphylococcus aureus Epidemiology in Wildlife: A Systematic Review. Antibiotics 2020, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauschild, T. Phenotypic and Genotypic Identification of Staphylococci Isolated from Wild Small Mammals. Syst. Appl. Microbiol. 2001, 24, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Hauschild, T.; Śliżewski, P.; Masiewicz, P. Species distribution of staphylococci from small wild mammals. Syst. Appl. Microbiol. 2010, 33, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.-H.; Lillehoj, H.; Lillehoj, E. Intestinal immune responses to coccidiosis. Dev. Comp. Immunol. 2000, 24, 303–324. [Google Scholar] [CrossRef]
- Duszynski, D.W.; Upton, S.J. Coccidia (Apicomplexa:Eimeriidae) of the Mammalian Order Insectivora. Special publication the Museum of Southwestern Biology; The University of New Mexico: New Mexico, NM, USA, 2000; No. 4; pp. 1–67. [Google Scholar]
- Duszynski, D.W. Biodiversity of the Coccidia (Apicomplexa: Conoidasida) in vertebrates: What we know, what we do not know, and what needs to be done. Folia Parasitol. 2021, 68, 1. [Google Scholar] [CrossRef]
- Kvicerova, J.; Hypsa, V. Host-Parasite Incongruences in Rodent Eimeria Suggest Significant Role of Adaptation Rather than Cophylogeny in Maintenance of Host Specificity. PLoS ONE 2013, 8, e63601. [Google Scholar] [CrossRef]
- Torres, J.; Gracenea, M.; Gómez, M.; Arrizabalaga, A.; González-Moreno, O. The occurrence of Cryptosporidium parvum and C. muris in wild rodents and insectivores in Spain. Veter.-Parasitol. 2000, 92, 253–260. [Google Scholar] [CrossRef]
- Čondlová, Š.; Horčičková, M.; Havrdová, N.; Sak, B.; Hlásková, L.; Perec-Matysiak, A.; Kicia, M.; McEvoy, J.; Kváč, M. Diversity of Cryptosporidium spp. in Apodemus spp. in Europe. Eur. J. Protistol. 2019, 69, 1–13. [Google Scholar] [CrossRef]
- Foo, C.; Farrell, J.; Boxell, A.; Robertson, I.; Ryan, U.M. Novel Cryptosporidium Genotype in Wild Australian Mice (Mus domesticus). Appl. Environ. Microbiol. 2007, 73, 7693–7696. [Google Scholar] [CrossRef] [Green Version]
- García-Livia, K.; Martín-Alonso, A.; Foronda, P. Diversity of Cryptosporidium spp. in wild rodents from the Canary Islands, Spain. Parasit. Vectors 2020, 13, 445. [Google Scholar] [CrossRef]
- Baer, J.-G. Contribution à la faune helminthologique de Suisse. Rev. Suisse Zool. 1928, 35, 27–41. [Google Scholar] [CrossRef]
- Soler, J.; Moreno, D.; Araujo, R.; Ramos, M.A. Diversidad y distribución de los moluscos de agua dulce en la Comunidad de Madrid (España). Graellsia 2006, 62, 201–252. [Google Scholar] [CrossRef] [Green Version]
- Yamaguti, S. Synopsis of Digenetic Trematodes of Vertebrates; Keigaku Publishing Co.: Tokyo, Japan, 1971; Volume I-II, p. 1074. [Google Scholar]
- Tkach, V.V.; Makarikov, A.A.; Kinsella, J.M. Morphological and molecular differentiation of Staphylocystis clydesengeri n. sp. (Cestoda, Hymenolepididae) from the vagrant shrew, Sorex vagrans (Soricomorpha, Soricidae), in North America. Zootaxa 2013, 3691, 389–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, M.J.; Beard, K.H.; Corlett, R.T.; Cumming, G.S.; Devictor, V.; Loyola, R.; Maas, B.; Miller-Rushing, A.J.; Pakeman, R.; Primack, R.B. Field work ethics in biological research. Biol. Conserv. 2016, 203, 268–271. [Google Scholar] [CrossRef]
- Waugh, C.A.; Monamy, V. Opposing Lethal Wildlife Research When Nonlethal Methods Exist: Scientific Whaling as a Case Study. J. Fish Wildl. Manag. 2016, 7, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.; Cheeseman, C. A mathematical model for the control of diseases in wildlife populations: Culling, vaccination and fertility control. Ecol. Model. 2002, 150, 45–53. [Google Scholar] [CrossRef]
- Zemanova, M.A.; Broennimann, O.; Guisan, A.; Knop, E.; Heckel, G. Slimy invasion: Climatic niche and current and future biogeography of Arion slug invaders. Divers. Distrib. 2018, 24, 1627–1640. [Google Scholar] [CrossRef] [Green Version]
- Zemanova, M.A. Towards more compassionate wildlife research through the 3Rs principles: Moving from invasive to non-invasive methods. Wildl. Biol. Nord. Board Wildl. Res. 2020, 2020, 11–19. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Area Zoning | A.D. | S.S. | S.S. with Desman | Total Samples | Desman | Water Shrews | Both | No Target |
|---|---|---|---|---|---|---|---|---|
| 238 | 207 | 8 | 4 | 19 | ||||
| Critical Zone | Ambroz | n = 4 | n = 2 | 13.45% | 10.92% | 0.84% | 0.00% | 1.68% |
| Jerte | n = 4 | n = 3 | 30.25% | 28.57% | 0.00% | 0.42% | 1.26% | |
| La Vera | n = 2 | n = 2 | 31.93% | 26.89% | 0.42% | 0.84% | 3.78% | |
| Subtotal | 75.63% | 66.39% | 1.26% | 1.26% | 6.72% | |||
| Importance Zone | Jerte | n = 1 | n = 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| Subtotal | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | |||
| Favourable Zone | Ambroz | n = 3 | n = 2 | 22.27% | 20.59% | 0.42% | 0.42% | 0.84% |
| Jerte | n = 4 | n = 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | |
| La Vera | n = 6 | n = 0 | 2.10% | 0.00% | 1.68% | 0.00% | 0.42% | |
| Subtotal | 24.37% | 20.59% | 2.10% | 0.42% | 1.26% |
| Size (Gene: 28S rDNA) | Technique | Primer Names | In Silico Species Range |
|---|---|---|---|
| Long PCR: ~934 bp | Conventional PCR | LahpmoF 5′TWCCGBRAGGGAAAGTTGAAA LahpmoR 5′TCACCATCYTTCGGGTCWCA | All in Table S2. |
| Short PCR: ~699 bp | Conventional, nested sequencing PCR | LahpmoF 5′TWCCGBRAGGGAAAGTTGAAA Ompahlshort_R 5’TCTCCTTGGTCCGTGTTT | All in Table S2 except Cercaria nigrospora |
| 76 bp | Real time PCR | Ompha_F 5′TCAAGTGTGTGCGCTCCG Ompha_R 5′TGCCGGTCGTGGTGACTA Probe 6-FAM-5′TCTCCGGCCTGCTCGTCAGT-BHQ-1 | Omphalometra; Neoglyphe and Rubenstrema in Table S2. |
| C | O | E | S | St | L | ac (1; 2 or 3) | |
|---|---|---|---|---|---|---|---|
| Year:2019/2020/2021 n = 71/90/46 | 1/2/0 0.797 (0.789–0.805) | 5/8/4 0.899 (0.893–0.905) | 63/83/45 0.201 (0.193–0.209) | 4/1/4 0.073 (0.068–0.078) | 9/18/3 0.095 (0.089–0.101) | 1/2/2 0.720 (0.711–0.729) | 16;1;1/19;6;0/ 11;1;0 0.591 (0.582–0.601) |
| Admin. District (A vs. J vs. V) n = 75/68/64 | 3/0/0 0.109 (0.103–0.115) | 5/8/4 0.489 (0.479–0.498) | 67/67/57 0.042 * (0.038–0.046) | 6/0/3 0.043 * (0.039–0.047) | 19 a/6 b/5 b 0.006 ** (0.005–0.008) | 2/3/0 0.321 (0.312–0.331) | 24;6;0 a/ 12;1;1 b/ 10;1;0 b 0.013 ** (0.011–0.053) |
| Zone (Crit. vs. Fav.) n = 158/49 | 1/2 0.140 | 15/2 0.371 | 149/42 0.650 | 4/5 0.036 * | 22/8 0.649 | 5/0 0.594 | 31;7;1/15;1;0 0.426 (0.416–0.436) |
| Sampling sections | |||||||
| 8 (A) n = 30 | 2 | 2 | 26 | 1 | 4 a,b | 0 | 7/1/0 a,b |
| 9 (A) n =19 | 1 | 3 | 19 | 0 | 10 a | 2 | 6/5/0 a |
| 13 (A) n= 7 | 0 | 0 | 6 | 1 | 1 a,b | 0 | 3/0/0 a,b |
| 14 (A) n= 19 | 0 | 0 | 16 | 4 | 4 a,b | 0 | 8/0/0 a,b |
| 16 (J) n = 22 | 0 | 2 | 22 | 0 | 2 a,b | 0 | 4/0/0 a,b |
| 21 (J) n = 7 | 0 | 2 | 7 | 0 | 1 a,b | 1 | 1/0/1 a,b |
| 24 (J) n = 39 | 0 | 4 | 38 | 0 | 3 b | 2 | 7/1/0 a,b |
| 10 (V) n= 31 | 0 | 3 | 28 | 0 | 3 b | 0 | 6/0/0 a,b |
| 19 (V) n = 33 | 0 | 1 | 29 | 3 | 2 b | 0 | 4/1/0 b |
| 0.256 (0.248–0.265) | 0.335 (0.326–0.344) | 0.194 (0.186–0.202) | 0.007 ** (0.005–0.008) | 0.004 ** (0.002–0.005) | 0.063 (0.059–0.068) | 0.008 ** (0.006–0.009) | |
| + | 1.15% | 8.2% | 92.3% | 4.3% | 14.5% | 2.4% | 26.6% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ripa, A.; Díaz-Caballero, J.A.; Palacios-González, M.J.; Zalba, J.; Espinosa, A.; García-Zapata, J.L.; Gómez-Martín, A.; Tkach, V.; Fernández-Garcia, J.L. Non-Invasive Wildlife Disease Surveillance Using Real Time PCR Assays: The Case of the Endangered Galemys pyrenaicus Populations from the Central System Mountains (Extremadura, Spain). Animals 2023, 13, 1136. https://doi.org/10.3390/ani13071136
Ripa A, Díaz-Caballero JA, Palacios-González MJ, Zalba J, Espinosa A, García-Zapata JL, Gómez-Martín A, Tkach V, Fernández-Garcia JL. Non-Invasive Wildlife Disease Surveillance Using Real Time PCR Assays: The Case of the Endangered Galemys pyrenaicus Populations from the Central System Mountains (Extremadura, Spain). Animals. 2023; 13(7):1136. https://doi.org/10.3390/ani13071136
Chicago/Turabian StyleRipa, Adriana, José A. Díaz-Caballero, María Jesús Palacios-González, Javier Zalba, Antonio Espinosa, Juan Luis García-Zapata, Ana Gómez-Martín, Vasyl Tkach, and José Luis Fernández-Garcia. 2023. "Non-Invasive Wildlife Disease Surveillance Using Real Time PCR Assays: The Case of the Endangered Galemys pyrenaicus Populations from the Central System Mountains (Extremadura, Spain)" Animals 13, no. 7: 1136. https://doi.org/10.3390/ani13071136