
Detection of Selected Canine Viruses in Nigerian Free-Ranging Dogs Traded for Meat Consumption
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
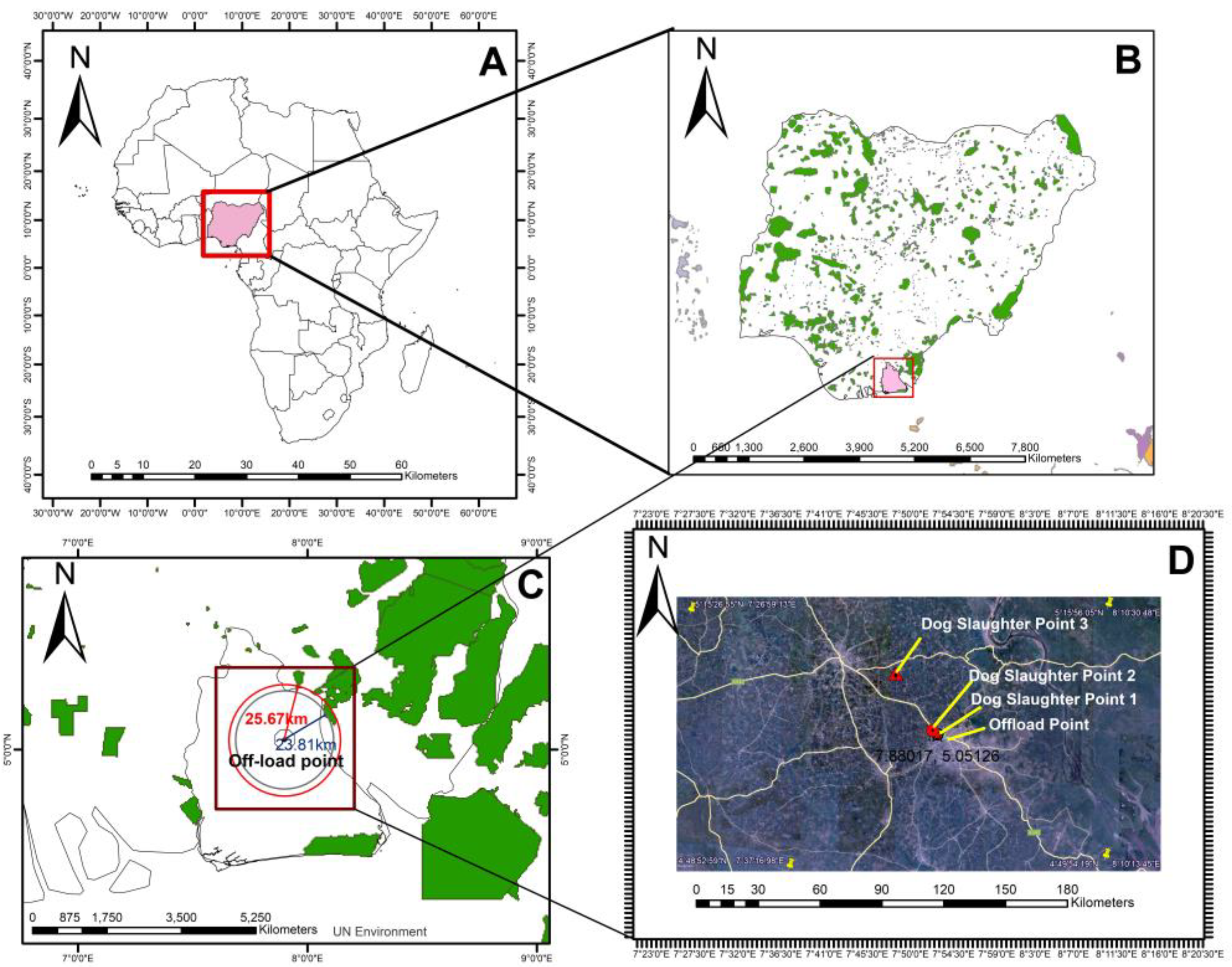
2.1. Sample Collection and Nucleic Acid Extraction
2.2. Screening for Canine Viruses
2.3. PCR Amplification, Sequencing, and Sequence Analyses
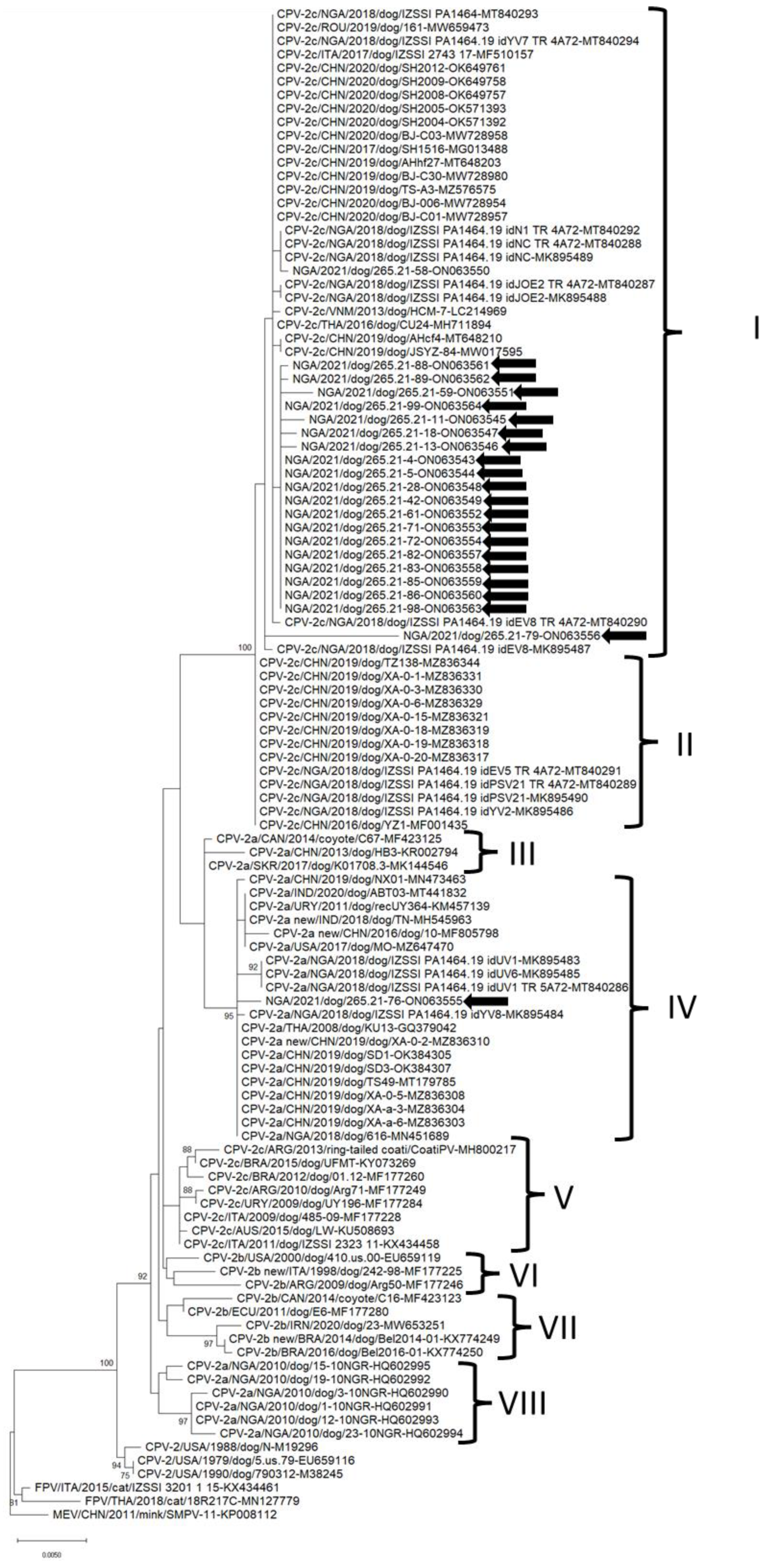
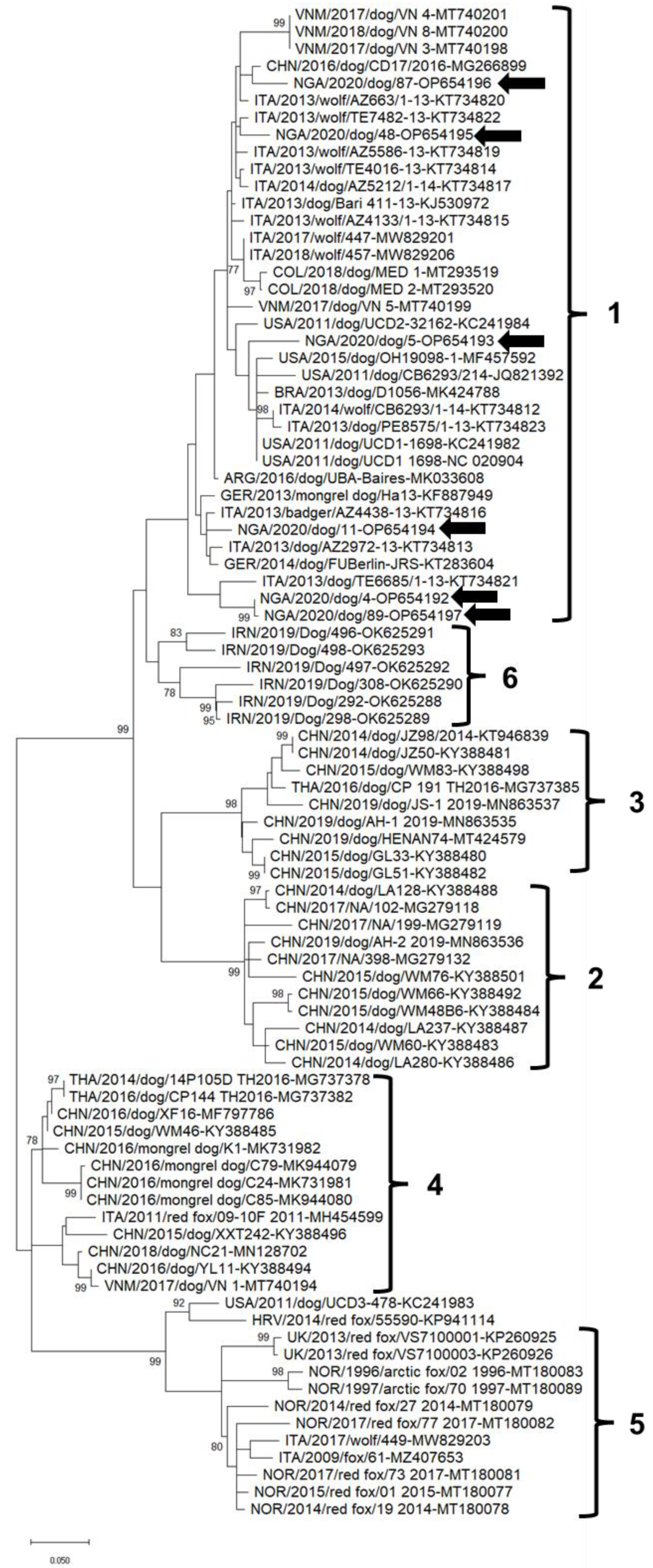
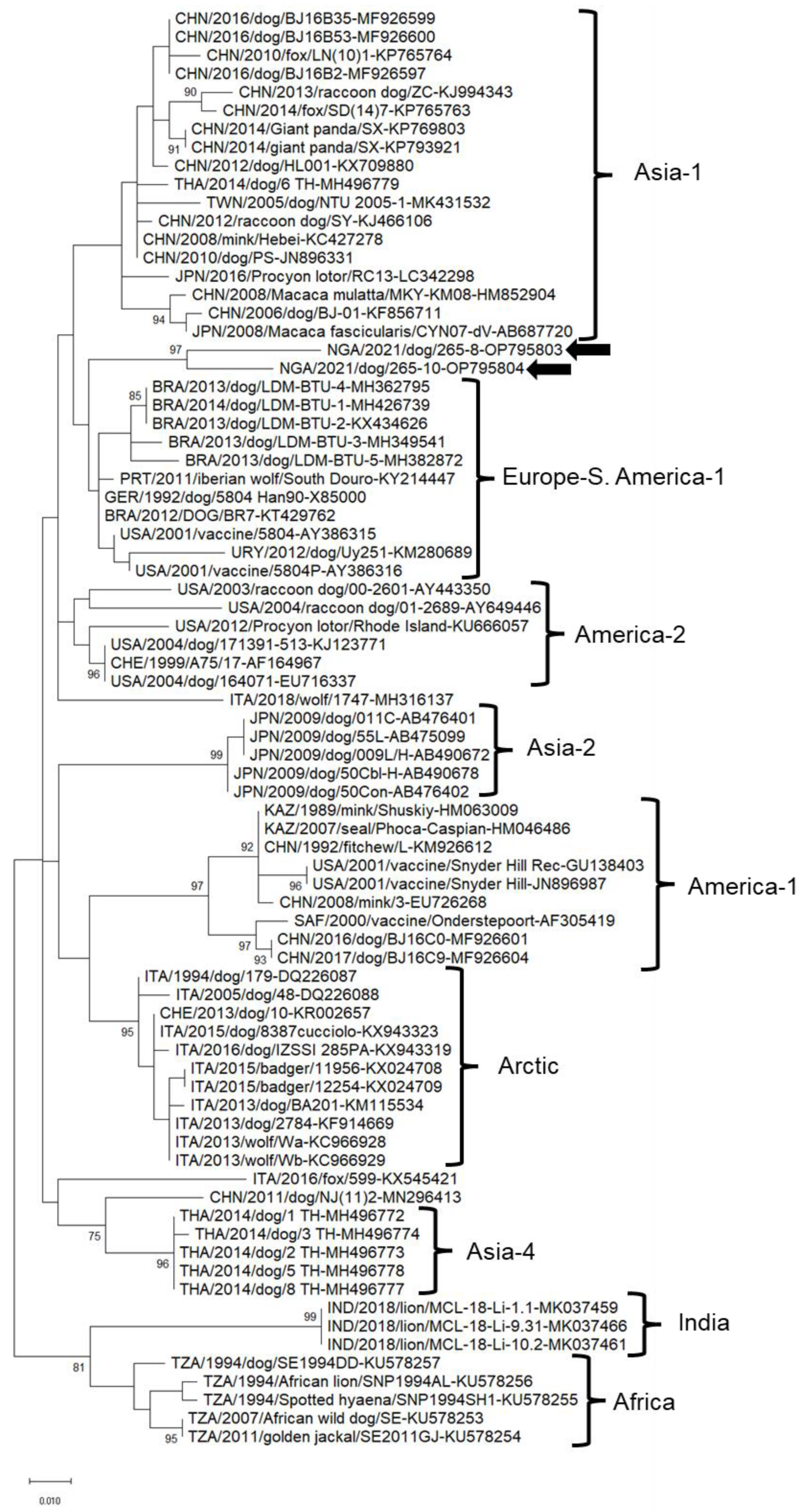
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Decaro, N.; Buonavoglia, C.; Barrs, V.R. Canine parvovirus vaccination and immunisation failures: Are we far from disease eradication? Vet. Microbiol. 2020, 247, 108760. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Elia, G.; Buonavoglia, C. Challenge studies for registration of canine core vaccines: Is it time to update the European Pharmacopeia? Vet. Microbiol. 2020, 244, 108659. [Google Scholar] [CrossRef] [PubMed]
- Ling, M.; Norris, J.; Kelman, M.; Ward, M.P. Risk factors for death from canine parvoviral-related disease in Australia. Vet. Microbiol. 2012, 158, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef]
- Reed, A.P.; Jones, E.V.; Miller, T.J. Nucleotide sequence and genome organization of canine parvovirus. J. Virol. 1988, 62, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Goetschius, D.J.; Hartmann, S.R.; Organtini, L.J.; Callaway, H.; Huang, K.; Bator, C.M.; Ashley, R.E.; Makhov, A.M.; Conway, J.F.; Parrish, C.R.; et al. High-resolution asymmetric structure of a Fab–virus complex reveals overlap with the receptor binding site. Proc. Natl. Acad. Sci. USA 2021, 118, e2025452118. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine parvovirus—A review of epidemiological and diagnostic aspects, with emphasis on type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine parvovirus post-vaccination shedding: Interference with diagnostic assays and correlation with host immune status. Vet. J. 2017, 221, 23–24. [Google Scholar] [CrossRef]
- Kamalu, B.P. Canine parvovirus infection in Nigeria. J. Small Anim. Pract. 1985, 26, 663–668. [Google Scholar] [CrossRef]
- Dogonyaro, B.B.; Bosman, A.-M.; Sibeko, K.P.; Venter, E.H.; van Vuuren, M. Genetic analysis of the VP2-encoding gene of canine parvovirus strains from Africa. Vet. Microbiol. 2013, 165, 460–465. [Google Scholar] [CrossRef]
- Fagbohun, O.A.; Omobowale, T.O. Sequence and phylogenetic analysis of canine parvovirus-2 isolates in dogs revealed circulation of three subtypes in Nigeria. Virusdisease 2018, 29, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Ndiana, L.A.; Odaibo, G.N.; Olaleye, D.O. Molecular characterization of canine parvovirus from domestic dogs in Nigeria: Introduction and spread of a CPV-2c mutant and replacement of older CPV-2a by the “new CPV-2a” strain. Virusdisease 2021, 32, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M.; Delwart, E.; Rosario, K.; Segalés, J.; Varsani, A. ICTV Report Consortium ICTV Virus Taxonomy Profile: Circoviridae. J. Gen. Virol. 2017, 98, 1997–1998. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; McGraw, S.; Zhu, K.; Leutenegger, C.M.; Marks, S.L.; Kubiski, S.; Gaffney, P.; Cruz, F.N., Jr.; Wang, C.; Delwart, E.; et al. Circovirus in Tissues of Dogs with Vasculitis and Hemorrhage. Emerg. Infect. Dis. 2013, 19, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Martella, V.; Desario, C.; Lanave, G.; Circella, E.; Cavalli, A.; Elia, G.; Camero, M.; Buonavoglia, C. Genomic Characterization of a Circovirus Associated with Fatal Hemorrhagic Enteritis in Dog, Italy. PLoS ONE 2014, 9, e105909. [Google Scholar] [CrossRef]
- Anderson, A.; Hartmann, K.; Leutenegger, C.M.; Proksch, A.L.; Mueller, R.S.; Unterer, S. Role of canine circovirus in dogs with acute haemorrhagic diarrhoea. Vet. Rec. 2017, 180, 542. [Google Scholar] [CrossRef]
- Tuong, N.M.; Piewbang, C.; Rungsipipat, A.; Techangamsuwan, S. Detection and molecular characterization of two canine circovirus genotypes co-circulating in Vietnam. Vet. Q. 2021, 41, 232–241. [Google Scholar] [CrossRef]
- Van Kruiningen, H.J.; Heishima, M.; Kerr, K.M.; Garmendia, A.E.; Helal, Z.; Smyth, J.A. Canine circoviral hemorrhagic enteritis in a dog in Connecticut. J. Vet. Diagn. Investig. 2019, 31, 732–736. [Google Scholar] [CrossRef]
- Dankaona, W.; Mongkholdej, E.; Satthathum, C.; Piewbang, C.; Techangamsuwan, S. Epidemiology, genetic diversity, and association of canine circovirus infection in dogs with respiratory disease. Sci. Rep. 2022, 12, 15445. [Google Scholar] [CrossRef]
- Piewbang, C.; Jo, W.K.; Puff, C.; van der Vries, E.; Kesdangsakonwut, S.; Rungsipipat, A.; Kruppa, J.; Jung, K.; Baumgärtner, W.; Techangamsuwan, S.; et al. Novel canine circovirus strains from Thailand: Evidence for genetic recombination. Sci. Rep. 2018, 8, 7524. [Google Scholar] [CrossRef] [Green Version]
- Cruz, T.F.; Batista, T.N.; Vieira, E.M.; Portela, L.M.F.; Baccarin, A.M.; Gradiz, J.J.; Araújo Junior, J.P. Genomic characterization of Canine circovirus detected in a dog with intermittent hemorrhagic gastroenteritis in Brazil. Ciência Rural 2020, 50. [Google Scholar] [CrossRef]
- Kotsias, F.; Bucafusco, D.; Nuñez, D.A.; Borisovsky, L.A.L.; Rodriguez, M.; Bratanich, A.C. Genomic characterization of canine circovirus associated with fatal disease in dogs in South America. PLoS ONE 2019, 14, e0218735. [Google Scholar] [CrossRef]
- Benkő, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarría, M.; Hess, M.; Jones, M.S.; Kaján, G.L.; Kajon, A.E.; Mittal, S.K.; et al. ICTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef]
- Decaro, N.; Martella, V.; Buonavoglia, C. Canine Adenoviruses and Herpesvirus. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 799–814. [Google Scholar] [CrossRef] [PubMed]
- Rima, B.; Balkema-Buschmann, A.; Dundon, W.G.; Duprex, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.; Lee, B.; Rota, P.; et al. ICTV Virus Taxonomy Profile: Paramyxoviridae. J. Gen. Virol. 2019, 100, 1593–1594. [Google Scholar] [CrossRef] [PubMed]
- Lempp, C.; Spitzbarth, I.; Puff, C.; Cana, A.; Kegler, K.; Techangamsuwan, S.; Baumgärtner, W.; Seehusen, F. New Aspects of the Pathogenesis of Canine Distemper Leukoencephalitis. Viruses 2014, 6, 2571–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuki, N.; Nakatsu, Y.; Kubota, T.; Sekizuka, T.; Seki, F.; Sakai, K.; Kuroda, M.; Yamaguchi, R.; Takeda, M. The V Protein of Canine Distemper Virus Is Required for Virus Replication in Human Epithelial Cells. PLoS ONE 2013, 8, e82343. [Google Scholar] [CrossRef]
- Rendon-Marin, S.; da Fontoura Budaszewski, R.; Canal, C.W.; Ruiz-Saenz, J. Tropism and molecular pathogenesis of canine distemper virus. Virol. J. 2019, 16, 30. [Google Scholar] [CrossRef] [Green Version]
- Mlanga, S.; Ibu, J.; Ezeokoli, C. Epidemiology of canine distemper in Makurdi, Nigeria. Niger. Vet. J. 2018, 39, 45–50. [Google Scholar] [CrossRef]
- Kia, G.; Huang, Y.; Zhou, M.; Zhou, Z.; Gnanadurai, C.; Leysona, C.; Umoh, J.; Kazeem, H.; Ehizibolo, D.; Kwaga, J.; et al. Molecular characterization of a rabies virus isolated from trade dogs in Plateau State, Nigeria. Sokoto J. Vet. Sci. 2018, 16, 54–62. [Google Scholar] [CrossRef]
- Odeh, L.E.; Umoh, J.U.; Dzikwi, A.A. Assessment of Risk of Possible Exposure to Rabies among Processors and Consumers of Dog Meat in Zaria and Kafanchan, Kaduna State, Nigeria. Glob. J. Health Sci. 2014, 6, 142–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurumyen, B.D.; Akanle, O.; Yikwabs, Y.P.; Nomishan, T.S. Zootherapy: The use of dog meat for traditional African medicine in Kanke local government area, Plateau state, Nigeria. J. Tour. Herit. Stud. 2020, 9, 1–14. [Google Scholar] [CrossRef]
- Ekanem, E.; Eyong, K.; Philip-Ephraim, E.; Eyong, M.; Adams, E.; Asindi, A. Stray dog trade fuelled by dog meat consumption as a risk factor for rabies infection in Calabar, southern Nigeria. Afr. Health Sci. 2013, 13, 1170–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaro, N.; Elia, G.; Martella, V.; Desario, C.; Campolo, M.; Di Trani, L.; Tarsitano, E.; Tempesta, M.; Buonavoglia, C. A real-time PCR assay for rapid detection and quantitation of canine parvovirus type 2 in the feces of dogs. Vet. Microbiol. 2005, 105, 19–28. [Google Scholar] [CrossRef]
- Decaro, N.; Elia, G.; Martella, V.; Campolo, M.; Desario, C.; Camero, M.; Cirone, F.; Lorusso, E.; Lucente, M.S.; Narcisi, D.; et al. Characterisation of the canine parvovirus type 2 variants using minor groove binder probe technology. J. Virol. Methods 2006, 133, 92–99. [Google Scholar] [CrossRef]
- Dowgier, G.; Mari, V.; Losurdo, M.; Larocca, V.; Colaianni, M.L.; Cirone, F.; Lucente, M.S.; Martella, V.; Buonavoglia, C.; Decaro, N. A duplex real-time PCR assay based on TaqMan technology for simultaneous detection and differentiation of canine adenovirus types 1 and 2. J. Virol. Methods 2016, 234, 1–6. [Google Scholar] [CrossRef]
- Elia, G.; Decaro, N.; Martella, V.; Cirone, F.; Lucente, M.S.; Lorusso, E.; Di Trani, L.; Buonavoglia, C. Detection of canine distemper virus in dogs by real-time RT-PCR. J. Virol. Methods 2006, 136, 171–176. [Google Scholar] [CrossRef]
- DeCaro, N.; Desario, C.; Miccolupo, A.; Campolo, M.; Parisi, A.; Martella, V.; Amorisco, F.; Lucente, M.S.; Lavazza, A.; Buonavoglia, C. Genetic analysis of feline panleukopenia viruses from cats with gastroenteritis. J. Gen. Virol. 2008, 89, 2290–2298. [Google Scholar] [CrossRef]
- Ikeda, Y.; Mochizuki, M.; Naito, R.; Nakamura, K.; Miyazawa, T.; Mikami, T.; Takahashi, E. Predominance of Canine Parvovirus (CPV) in Unvaccinated Cat Populations and Emergence of New Antigenic Types of CPVs in Cats. Virology 2000, 278, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Kapoor, A.; Slikas, B.; Bamidele, O.S.; Wang, C.; Shaukat, S.; Alam Masroor, M.; Wilson, M.L.; Ndjango, J.-B.N.; Peeters, M.; et al. Multiple Diverse Circoviruses Infect Farm Animals and Are Commonly Found in Human and Chimpanzee Feces. J. Virol. 2010, 84, 1674–1682. [Google Scholar] [CrossRef] [Green Version]
- Frisk, A.L.; König, M.; Moritz, A.; Baumgärtner, W. Detection of Canine Distemper Virus Nucleoprotein RNA by Reverse Transcription-PCR Using Serum, Whole Blood, and Cerebrospinal Fluid from Dogs with Distemper. J. Clin. Microbiol. 1999, 37, 3634–3643. [Google Scholar] [CrossRef] [Green Version]
- Loots, A.K.; Mokgokong, P.S.; Mitchell, E.; Venter, E.H.; Kotze, A.; Dalton, D.L. Phylogenetic analysis of canine distemper virus in South African wildlife. PLoS ONE 2018, 13, e0199993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ogbu, K.I.; Mira, F.; Purpari, G.; Nwosuh, C.; Loria, G.R.; Schirò, G.; Chiaramonte, G.; Tion, M.T.; Di Bella, S.; Ventriglia, G.; et al. Nearly full-length genome characterization of canine parvovirus strains circulating in Nigeria. Transbound. Emerg. Dis. 2020, 67, 635–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogbu, K.I.; Chukwudi, I.C.; Mira, F.; Eze, U.U.; Di Bella, S.; Olaolu, O.S.; Tion, M.T.; Purpari, G.; Cannella, V.; Nwosuh, I.C.; et al. Current status and risk factors of canine parvovirus type 2 in North Central Nigeria. Comp. Immunol. Microbiol. Infect. Dis. 2021, 74, 101578. [Google Scholar] [CrossRef]
- Allison, A.B.; Organtini, L.J.; Zhang, S.; Hafenstein, S.L.; Holmes, E.; Parrish, C.R. Single Mutations in the VP2 300 Loop Region of the Three-Fold Spike of the Carnivore Parvovirus Capsid Can Determine Host Range. J. Virol. 2015, 90, 753–767. [Google Scholar] [CrossRef] [Green Version]
- Moon, B.; Jang, J.; Kim, S.; Kim, Y.; Lee, H.; So, B.; Park, C.; Lee, K. Genetic characterization of canine parvovirus type 2c from domestic dogs in Korea. Transbound. Emerg. Dis. 2020, 67, 1645–1653. [Google Scholar] [CrossRef] [Green Version]
- Chollom, S.C.; Fyaktu, E.J.; Okwori, A.E.J.; Agada, G.O.A.; Hashimu, G.; Akele, R.Y.; Voumangai, E.I.; Dashe, T.; Egah, D.Z. Molecular detection of canine parvovirus in Jos, Nigeria. J. Vet. Med. Anim. Health 2013, 5, 57–59. [Google Scholar] [CrossRef]
- Miranda, C.; Carvalheira, J.; Parrish, C.R.; Thompson, G. Factors affecting the occurrence of canine parvovirus in dogs. Vet. Microbiol. 2015, 180, 59–64. [Google Scholar] [CrossRef]
- Pedroza-Roldán, C.; Páez-Magallan, V.; Charles-Niño, C.; Elizondo-Quiroga, D.; De Cervantes-Mireles, R.L.; López-Amezcua, M.A. Genotyping of Canine parvovirus in western Mexico. J. Vet. Diagn. Investig. 2015, 27, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Ohneiser, S.; Hills, S.; Cave, N.; Passmore, D.; Dunowska, M. Canine parvoviruses in New Zealand form a monophyletic group distinct from the viruses circulating in other parts of the world. Vet. Microbiol. 2015, 178, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Mira, F.; Purpari, G.; Di Bella, S.; Colaianni, M.L.; Schirò, G.; Chiaramonte, G.; Gucciardi, F.; Pisano, P.; Lastra, A.; Decaro, N.; et al. Spreading of canine parvovirus type 2c mutants of Asian origin in southern Italy. Transbound. Emerg. Dis. 2019, 66, 2297–2304. [Google Scholar] [CrossRef] [PubMed]
- Ndiana, L.A.; Lanave, G.; Desario, C.; Berjaoui, S.; Alfano, F.; Puglia, I.; Fusco, G.; Colaianni, M.L.; Vincifori, G.; Camarda, A.; et al. Circulation of diverse protoparvoviruses in wild carnivores, Italy. Transbound. Emerg. Dis. 2021, 68, 2489–2502. [Google Scholar] [CrossRef]
- Wang, J.; Lin, P.; Zhao, H.; Cheng, Y.; Jiang, Z.; Zhu, H.; Wu, H.; Cheng, S. Continuing evolution of canine parvovirus in China: Isolation of novel variants with an Ala5Gly mutation in the VP2 protein. Infect. Genet. Evol. 2016, 38, 73–78. [Google Scholar] [CrossRef]
- Geng, Y.; Guo, D.; Li, C.; Wang, E.; Wei, S.; Wang, Z.; Yao, S.; Zhao, X.; Su, M.; Wang, X.; et al. Co-Circulation of the Rare CPV-2c with Unique Gln370Arg Substitution, New CPV-2b with Unique Thr440Ala Substitution, and New CPV-2a with High Prevalence and Variation in Heilongjiang Province, Northeast China. PLoS ONE 2015, 10, e0137288. [Google Scholar] [CrossRef]
- Simpson, A.A.; Chandrasekar, V.; Hébert, B.; Sullivan, G.M.; Rossmann, M.G.; Parrish, C.R. Host range and variability of calcium binding by surface loops in the capsids of canine and feline parvoviruses. J. Mol. Biol. 2000, 300, 597–610. [Google Scholar] [CrossRef] [Green Version]
- Govindasamy, L.; Hueffer, K.; Parrish, C.R.; Agbandje-McKenna, M. Structures of Host Range-Controlling Regions of the Capsids of Canine and Feline Parvoviruses and Mutants. J. Virol. 2003, 77, 12211–12221. [Google Scholar] [CrossRef] [Green Version]
- Palermo, L.M.; Hueffer, K.; Parrish, C.R. Residues in the Apical Domain of the Feline and Canine Transferrin Receptors Control Host-Specific Binding and Cell Infection of Canine and Feline Parvoviruses. J. Virol. 2003, 77, 8915–8923. [Google Scholar] [CrossRef] [Green Version]
- Truyen, U.; Parrish, C.R. Canine and feline host ranges of canine parvovirus and feline panleukopenia virus: Distinct host cell tropisms of each virus in vitro and in vivo. J. Virol. 1992, 66, 5399–5408. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, P.S.B.; Cargnelutti, J.F.; Masuda, E.K.; Weiblen, R.; Flores, E.F. New variants of canine parvovirus in dogs in southern Brazil. Arch. Virol. 2019, 164, 1361–1369. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, X.; Zhu, J.; Liao, L.; Bao, E. Molecular Epidemiological Survey of Canine Parvovirus Circulating in China from 2014 to 2019. Pathogens 2021, 10, 588. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Desario, C.; Parisi, A.; Martella, V.; Lorusso, A.; Miccolupo, A.; Mari, V.; Colaianni, M.L.; Cavalli, A.; Di Trani, L.; et al. Genetic analysis of canine parvovirus type 2c. Virology 2009, 385, 5–10. [Google Scholar] [CrossRef]
- Kapoor, A.; Dubovi, E.J.; Henriquez-Rivera, J.A.; Lipkin, W.I. Complete Genome Sequence of the First Canine Circovirus. J. Virol. 2012, 86, 7018. [Google Scholar] [CrossRef] [Green Version]
- Beikpour, F.; Ndiana, L.A.; Sazmand, A.; Capozza, P.; Nemati, F.; Pellegrini, F.; Zafari, S.; Zolhavarieh, S.M.; Cardone, R.; Faraji, R.; et al. Detection and Genomic Characterization of Canine Circovirus in Iran. Animals 2022, 12, 507. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Ramirez, S.; Rendon-Marin, S.; Vargas-Bermudez, D.S.; Jaime, J.; Ruiz-Saenz, J. First detection and full genomic analysis of Canine Circovirus in CPV-2 infected dogs in Colombia, South America. Sci. Rep. 2020, 10, 17579. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Wang, Z.; Zhao, L.; Wang, Y.; Cui, X.; Shi, Y.; Chen, H.; Ge, J. Detection and molecular characterization of canine circovirus circulating in northeastern China during 2014–2016. Arch. Virol. 2020, 165, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Turan, T.; Işıdan, H. Molecular characterization of canine astrovirus, vesivirus and circovirus, isolated from diarrheic dogs in Turkey. Iran. J. Vet. Res. 2020, 21, 172–179. [Google Scholar] [CrossRef]
- Weber, M.; Cibulski, S.; Olegário, J.; da Silva, M.; Puhl, D.; Mósena, A.; Alves, C.; Paim, W.; Baumbach, L.; Mayer, F.; et al. Characterization of dog serum virome from Northeastern Brazil. Virology 2018, 525, 192–199. [Google Scholar] [CrossRef]
- Panzera, Y.; Sarute, N.; Iraola, G.; Hernández, M.; Pérez, R. Molecular phylogeography of canine distemper virus: Geographic origin and global spreading. Mol. Phylogenet. Evol. 2015, 92, 147–154. [Google Scholar] [CrossRef]
- Haas, L.; Hofer, H.; East, M.; Wohlsein, P.; Liess, B.; Barrett, T. Canine distemper virus infection in Serengeti spotted hyaenas. Vet. Microbiol. 1996, 49, 147–152. [Google Scholar] [CrossRef]
- Roelke-Parker, M.E.; Munson, L.; Packer, C.; Kock, R.; Cleaveland, S.; Carpenter, M.; O'Brien, S.J.; Pospischil, A.; Hofmann-Lehmann, R.; Lutz, H.; et al. A canine distemper virus epidemic in Serengeti lions (Panthera leo). Nature 1996, 379, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T.; Mazet, J.K.; Holekamp, K.E.; Dubovi, E.; Engh, A.L.; Nelson, K.; Van Horn, R.C.; Munson, L. Antibodies to canine and feline viruses in spotted hyenas (Crocuta crocuta) in the masai mara national reserve. J. Wildl. Dis. 2004, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Almberg, E.S.; Cross, P.C.; Smith, D.W. Persistence of canine distemper virus in the Greater Yellowstone Ecosystem's carnivore community. Ecol. Appl. 2010, 20, 2058–2074. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.C.; Wilkes, R.P. Sequencing of emerging canine distemper virus strain reveals new distinct genetic lineage in the United States associated with disease in wildlife and domestic canine populations. Virol. J. 2015, 12, 219. [Google Scholar] [CrossRef] [Green Version]
- Alfano, F.; Lanave, G.; Lucibelli, M.G.; Miletti, G.; D’Alessio, N.; Gallo, A.; Auriemma, C.; Amoroso, M.G.; Lucente, M.S.; De Carlo, E.; et al. Canine Distemper Virus in Autochtonous and Imported Dogs, Southern Italy (2014–2021). Animals 2022, 12, 2852. [Google Scholar] [CrossRef]
- Nikolin, V.M.; Olarte-Castillo, X.A.; Osterrieder, N.; Hofer, H.; Dubovi, E.; Mazzoni, C.J.; Brunner, E.; Goller, K.V.; Fyumagwa, R.D.; Moehlman, P.D.; et al. Canine distemper virus in the Serengeti ecosystem: Molecular adaptation to different carnivore species. Mol. Ecol. 2017, 26, 2111–2130. [Google Scholar] [CrossRef]
- Karki, M.; Rajak, K.K.; Singh, R.P. Canine morbillivirus (CDV): A review on current status, emergence and the diagnostics. Virusdisease 2022, 33, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.D.; Gräf, T.; Ikuta, N.; Lehmann, F.K.; Passos, D.T.; Makiejczuk, A.; Silveira, M.A.; Fonseca, A.S.; Canal, C.W.; Lunge, V.R. Phylogenetic analysis of canine distemper virus in South America clade 1 reveals unique molecular signatures of the local epidemic. Infect. Genet. Evol. 2016, 41, 135–141. [Google Scholar] [CrossRef]
- Curi, N.H.D.A.; Massara, R.L.; Paschoal, A.M.D.O.; Soriano-Araújo, A.; Lobato, Z.I.P.; Demetrio, G.R.; Chiarello, A.G.; Passamani, M. Prevalence and risk factors for viral exposure in rural dogs around protected areas of the Atlantic forest. BMC Vet. Res. 2016, 12, 21. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Real-Time Results | CT Values | ||||
|---|---|---|---|---|---|---|
| Pos | Neg | Total | Range | Mean | Median | |
| CPPV-1 | 83 | 17 | 100 | 15 to 34 | 33 | 34 |
| CanineCV | 14 | 86 | 100 | 28 to 35 | 33 | 35 |
| CDV | 17 | 83 | 100 | 22 to 40 | 33 | 33 |
| CAdV-1 | 0 | 100 | 100 | - | - | - |
| CAdV-2 | 0 | 100 | 100 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndiana, L.A.; Lanave, G.; Desario, C.; Odigie, A.E.; Madubuike, K.G.; Lucente, M.S.; Ezeifeka, C.A.; Patruno, G.; Lorusso, E.; Elia, G.; et al. Detection of Selected Canine Viruses in Nigerian Free-Ranging Dogs Traded for Meat Consumption. Animals 2023, 13, 1119. https://doi.org/10.3390/ani13061119
Ndiana LA, Lanave G, Desario C, Odigie AE, Madubuike KG, Lucente MS, Ezeifeka CA, Patruno G, Lorusso E, Elia G, et al. Detection of Selected Canine Viruses in Nigerian Free-Ranging Dogs Traded for Meat Consumption. Animals. 2023; 13(6):1119. https://doi.org/10.3390/ani13061119
Chicago/Turabian StyleNdiana, Linda A., Gianvito Lanave, Costantina Desario, Amienwanlen E. Odigie, Kelechi G. Madubuike, Maria Stella Lucente, Chukwuemeka A. Ezeifeka, Giovanni Patruno, Eleonora Lorusso, Gabriella Elia, and et al. 2023. "Detection of Selected Canine Viruses in Nigerian Free-Ranging Dogs Traded for Meat Consumption" Animals 13, no. 6: 1119. https://doi.org/10.3390/ani13061119