
Differences in the Natural Swimming Behavior of Schizothorax prenanti Individual and Schooling in Spatially Heterogeneous Turbulent Flows
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Apparatus
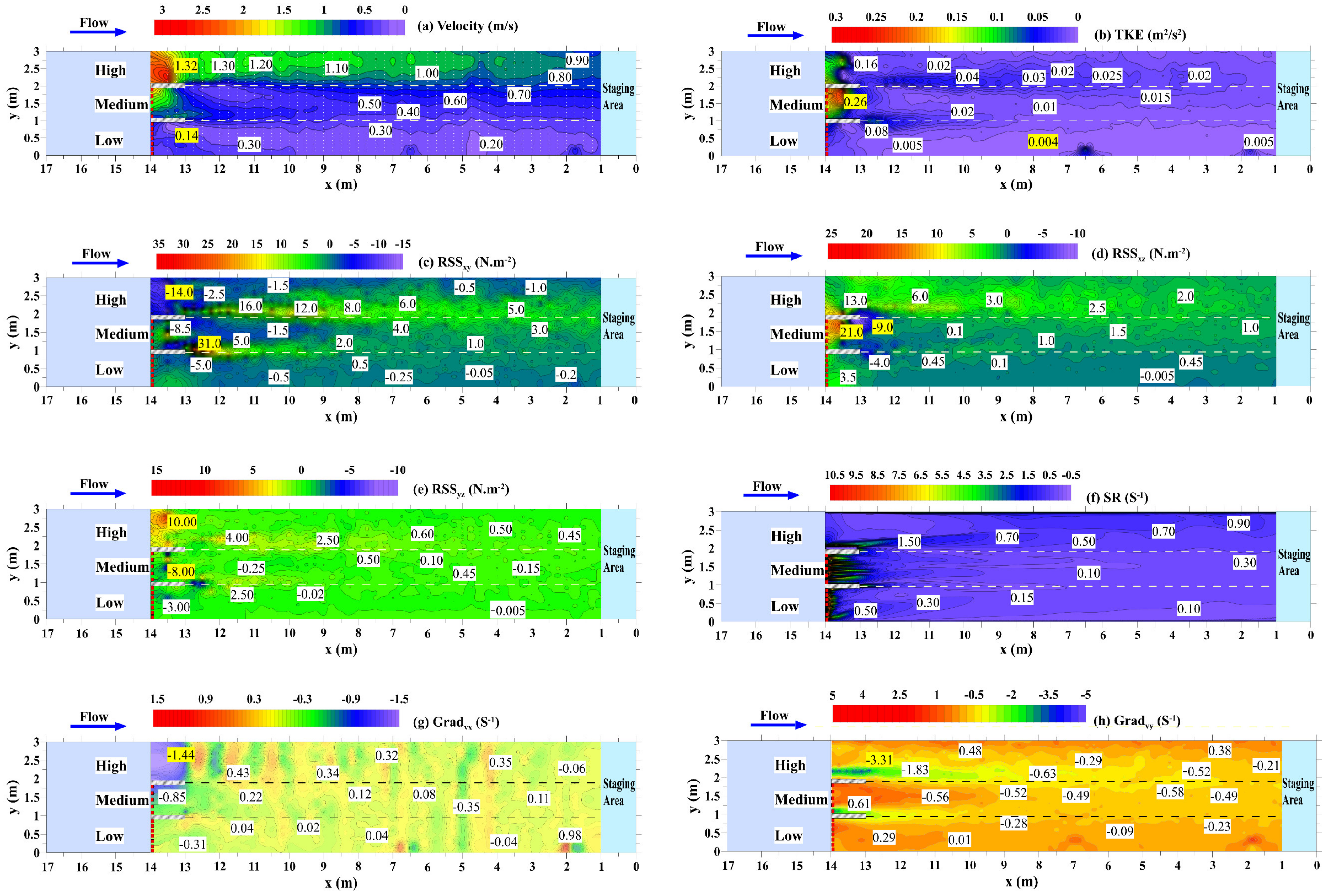
2.2. Hydraulic Characteristics
2.3. Fish Species Husbandry
2.4. Experimental Methods
2.5. Swimming Performance
2.5.1. Swimming Capability
2.5.2. Swimming Trajectory
2.5.3. Preferred Hydrodynamic Range
2.6. Data Analysis
3. Results
3.1. Flume Hydraulics and Tolerances
3.2. Swimming Speed Difference
3.3. Spatial-Temporal Distributions of Swimming Trajectories
3.3.1. Swimming Trajectories
Fish Individual
Fish Schooling
3.3.2. Occupied Positions
3.3.3. Residence Time
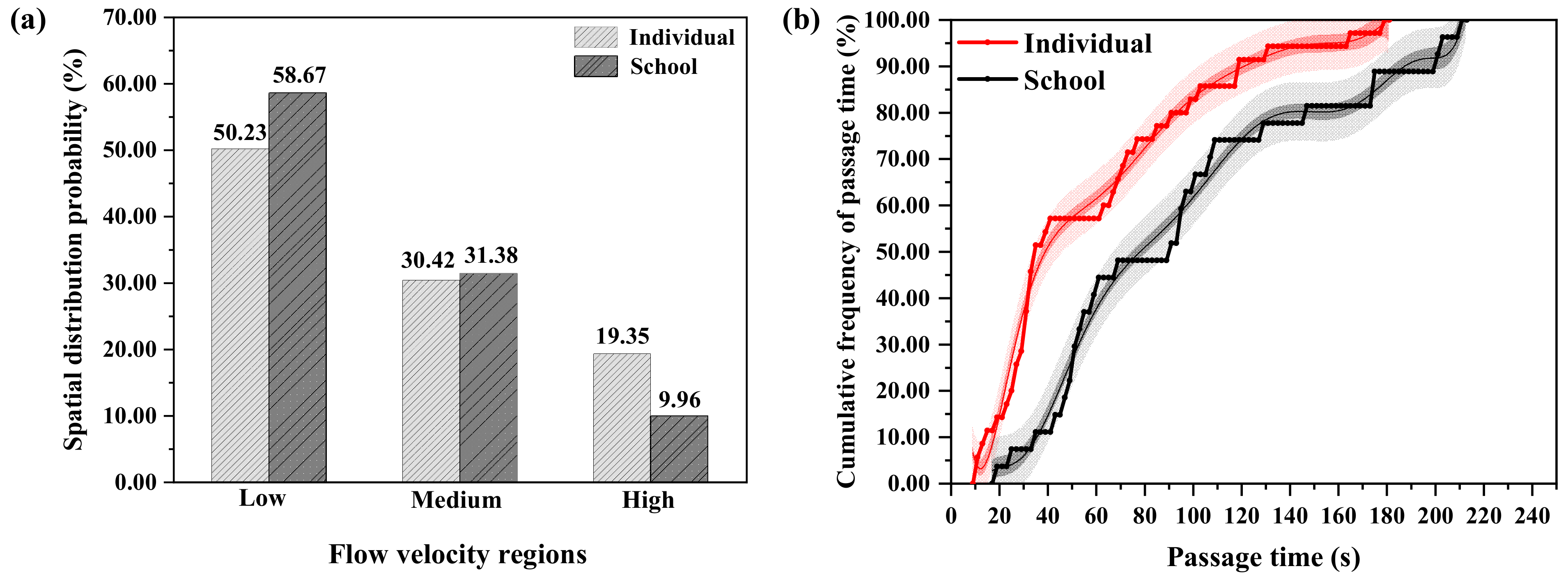
3.3.4. Spatial Distribution Probability and Passage Time
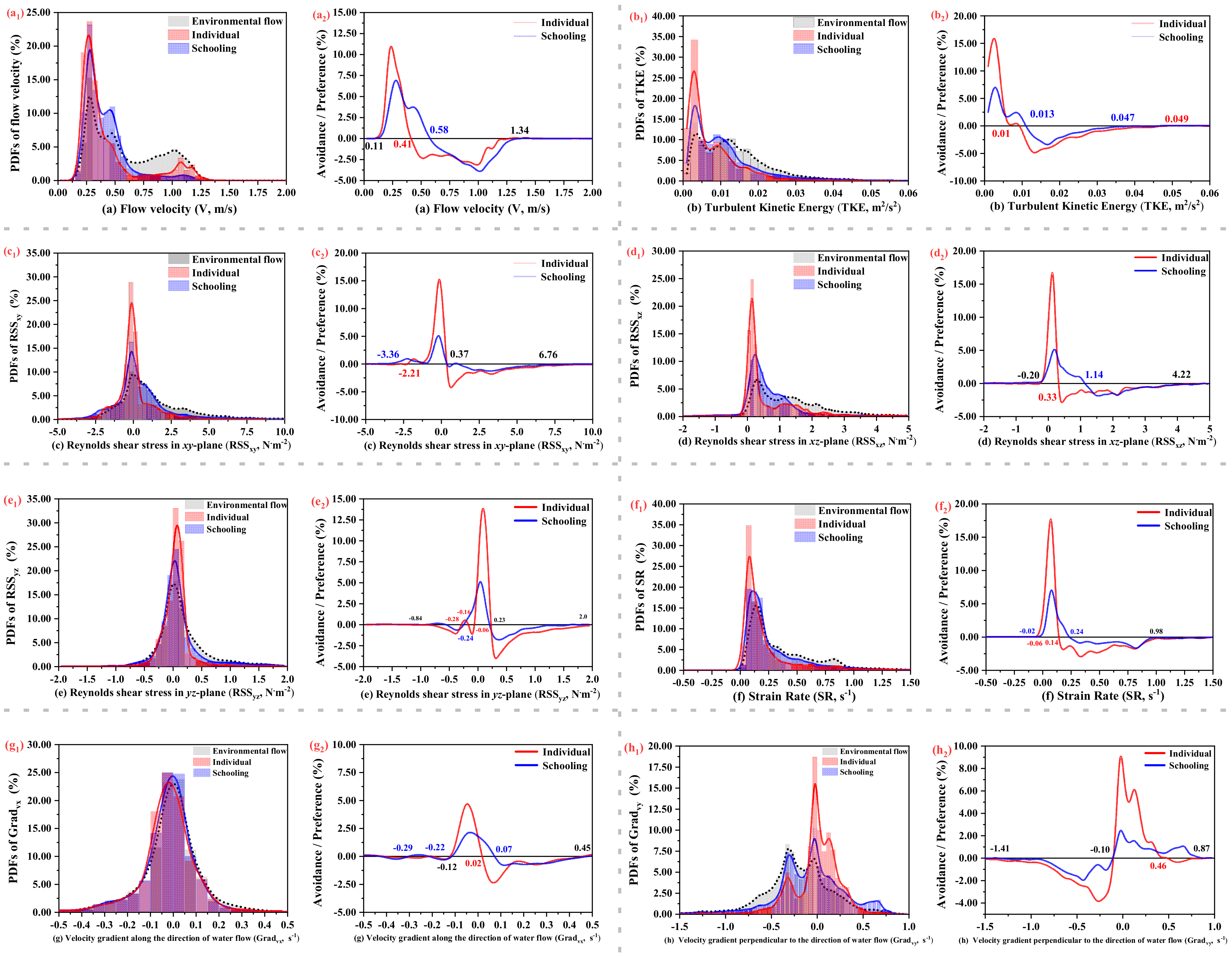
3.4. Hydrodynamic Parameter Preferences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bednarek, A.T. Undamming Rivers: A Review of the Ecological Impacts of Dam Removal. Environ. Manag. 2001, 27, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.M.; Martins, E.G.; Algera, D.A.; Rytwinski, T.; Mossop, B.; Leake, A.J.; Power, M.; Cooke, S.J. Turbine entrainment and passage of potadromous fish through hydropower dams: Developing conceptual frameworks and metrics for moving beyond turbine passage mortality. Fish Fish. 2019, 20, 403–418. [Google Scholar] [CrossRef]
- Barbarossa, V.; Schmitt, R.J.P.; Huijbregts, M.A.J.; Zarfl, C.; King, H.; Schipper, A.M. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide. Proc. Natl. Acad. Sci. USA 2020, 117, 3648–3655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Shi, X.; Jin, Z.; Ke, S.; Lin, C.; An, R.; Li, J.; Katopodis, C. Behaviour and ability of a cyprinid (Schizopygopsis younghusbandi) to cope with accelerating flows when migrating downstream. River Res. Appl. 2021, 37, 1168–1179. [Google Scholar] [CrossRef]
- Zhang, P.; Li, K.; Liu, Q.; Liu, R.; Qin, L.; Wang, H.; Zhang, Z.; Wang, K.; Wang, Y.; Liang, R.; et al. Linking bait and feeding opportunities to fish foraging habitat for the assessment of environmental flows and river restoration. Sci. Total Environ. 2021, 768, 144580. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, P.; Cheng, B.; Li, Y.; Li, J.; Zhou, H.; Sun, G.; Qing, J.; Zhu, Z.; Lu, Y.; et al. Incorporating the life stages of fish into habitat assessment frameworks: A case study in the Baihetan Reservoir. J. Environ. Manag. 2021, 299, 113663. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Ke, S.; Tu, Z.; Wang, Y.; Tan, J.; Guo, W. Swimming capability of target fish from eight hydropower stations in China relative to fishway design. Can. J. Fish. Aquat. Sci. 2021, 79, 124–132. [Google Scholar] [CrossRef]
- Tan, J.; Gao, Z.; Dai, H.; Yang, Z.; Shi, X. Effects of turbulence and velocity on the movement behaviour of bighead carp (Hypophthalmichthys nobilis) in an experimental vertical slot fishway. Ecol. Eng. 2019, 127, 363–374. [Google Scholar] [CrossRef]
- Shi, X.; Kynard, B.; Liu, D.; Qiao, Y.; Chen, Q. Development of Fish Passage in China. Fisheries 2015, 40, 161–169. [Google Scholar] [CrossRef]
- Lin, C.; Dai, H.; Shi, X.; Deng, Z.D.; Mao, J.; Zhao, S.; Luo, J.; Tan, J. An experimental study on fish attraction using a fish barge model. Fish. Res. 2019, 210, 181–188. [Google Scholar] [CrossRef]
- Ke, S.; Li, Z.; Jiang, Z.; Goerig, E.; Kynard, B.; Liu, D.; Shi, X. Effect of a vertical half cylinder on swimming of silver carp, Hypophthalmichthys molitrix: Implications for microhabitat restoration and fishway design. River Res. Appl. 2019, 35, 436–441. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Katopodis, C.; Johnson, D.; Zhang, P.; Zhao, P. Case study: Targeting species and applying swimming performance data to fish lift design for the Huangdeng Dam on the upper Mekong River. Ecol. Eng. 2018, 122, 32–38. [Google Scholar] [CrossRef]
- Mao, X. Review of fishway research in China. Ecol. Eng. 2018, 115, 91–95. [Google Scholar] [CrossRef]
- Feng, R. Strategy of Stock Enhancement Using EnhanceFish Model: A Case of Acanthopagrus latus Enhancement in Nanlang Water Area, Zhongshan City. Prog. Fish. Sci. 2022, 43, 1–10. [Google Scholar]
- Qin, C.X.; Pan, W.N.; Yu, G.; Zuo, T.; Xi, S.G.; Zhu, W.T.; Ma, H.M. Review on Environmental Enrichment for Aquatic Organisms. Prog. Fish. Sci. 2020, 41, 185–193. [Google Scholar]
- Silva, A.T.; Bærum, K.M.; Hedger, R.D.; Baktoft, H.; Fjeldstad, H.-P.; Gjelland, K.; Økland, F.; Forseth, T. The effects of hydrodynamics on the three-dimensional downstream migratory movement of Atlantic salmon. Sci. Total Environ. 2020, 705, 135773. [Google Scholar] [CrossRef]
- Katopodis, C.; Cai, L.; Johnson, D. Sturgeon survival: The role of swimming performance and fish passage research. Fish. Res. 2019, 212, 162–171. [Google Scholar] [CrossRef]
- Wang, Y.; Wai, O.; Chen, Q. Laboratory study on fish behavioral response to meandering flow and riffle-pool sequence driven by deflectors in straight concrete flood channels. J. Hydrol. 2021, 598, 125736. [Google Scholar] [CrossRef]
- Li, G.; Sun, S.; Liu, H.; Zheng, T. Schizothorax prenanti swimming behavior in response to different flow patterns in vertical slot fishways with different slot positions. Sci. Total Environ. 2021, 754, 142142. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.A.; Politano, M.; Garvin, J.W.; Nestler, J.M.; Hay, D.; Anderson, J.J.; Weber, L.J.; Dimperio, E.; Smith, D.L.; Timko, M. Fish navigation of large dams emerges from their modulation of flow field experience. Proc. Natl. Acad. Sci. USA 2014, 111, 5277–5282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhawenimana, V.; Wilson, C.A.M.E.; Ouro, P.; Cable, J. Spanwise Cylinder Wake Hydrodynamics and Fish Behavior. Water Resour. Res. 2019, 55, 8569–8582. [Google Scholar] [CrossRef]
- Zha, W.; Zeng, Y.; Katul, G.; Li, Q.; Liu, X.; Chen, X. Laboratory study on behavioral responses of hybrid sturgeon, Acipenseridae, to wake flows induced by cylindrical bluff bodies. Sci. Total Environ. 2021, 799, 149403. [Google Scholar] [CrossRef]
- Hinch, S.G.; Rand, P.S. Optimal swimming speeds and forward-assisted propulsion: Energy-conserving behaviours of upriver-migrating adult salmon. Can. J. Fish. Aquat. Sci. 2000, 57, 2470–2478. [Google Scholar] [CrossRef]
- Chen, M.; An, R.; Li, J.; Li, K.; Li, F. Identifying operation scenarios to optimize attraction flow near fishway entrances for endemic fishes on the Tibetan Plateau of China to match their swimming characteristics: A case study. Sci. Total Environ. 2019, 693, 133615. [Google Scholar] [CrossRef]
- Castro-Santos, T.; Sanz-Ronda, F.J.; Ruiz-Legazpi, J. Breaking the speed limit—Comparative sprinting performance of brook trout (Salvelinus fontinalis) and brown trout (Salmo trutta). Can. J. Fish. Aquat. Sci. 2013, 70, 280–293. [Google Scholar] [CrossRef]
- Weihs, D. Hydromechanics of Fish Schooling. Nature 1973, 241, 290–291. [Google Scholar] [CrossRef]
- Capoccioni, F.; Leone, C.; Pulcini, D.; Cecchetti, M.; Rossi, A.; Ciccotti, E. Fish movements and schooling behavior across the tidal channel in a Mediterranean coastal lagoon: An automated approach using acoustic imaging. Fish. Res. 2019, 219, 105318. [Google Scholar] [CrossRef]
- Liao, L.; Chen, M.; An, R.; Li, J.; Tang, X.; Yan, Z. Identifying three-dimensional swimming corridors for fish to match their swimming characteristics under different hydropower plant operations: Optimization of entrance location for fish-passing facilities. Sci. Total Environ. 2022, 822, 153599. [Google Scholar] [CrossRef]
- An, R.-D.; Li, J.; Yi, W.-M.; Mao, X. Hydraulics and swimming behavior of Schizothorax prenanti in vertical slot fishways. J. Hydrodyn. 2019, 31, 169–176. [Google Scholar] [CrossRef]
- Zeng, Q.; Hu, P.; Wang, H.; Pan, J.; Yang, Z.; Liu, H. The influence of cascade hydropower development on the hydrodynamic conditions impacting the reproductive process of fish with semi-buoyant eggs. Sci. Total Environ. 2019, 689, 865–874. [Google Scholar] [CrossRef]
- Li, M.; An, R.; Chen, M.; Li, J. Evaluation of volitional swimming behavior of Schizothorax prenanti using an open-channel flume with spatially heterogeneous turbulent flow. Animals 2022, 12, 752. [Google Scholar] [CrossRef]
- Cai, L.; Liu, G.; Taupier, R.; Fang, M.; Johnson, D.; Tu, Z.; Huang, Y. Effect of temperature on swimming performance of juvenile Schizothorax prenanti. Fish. Physiol. Biochem. 2014, 40, 491–498. [Google Scholar] [CrossRef]
- Silva, A.T.; Santos, J.M.; Ferreira, M.T.; Pinheiro, A.N.; Katopodis, C. Effects of water velocity and turbulence on the behaviour of Iberian barbel (Luciobarbus bocagei, Steindachner 1864) in an experimental pool-type fishway. River Res. Appl. 2011, 27, 360–373. [Google Scholar] [CrossRef]
- Wahl, T.L. Analyzing ADV Data Using WinADV. In Proceedings of the Joint Conference on Water Resource Engineering and Water Resources Planning and Management, Minneapolis, MN, USA, 30 July–2 August 2000. [Google Scholar]
- Goodwin, R.A.; Nestler, J.M.; Anderson, J.J.; Weber, L.J.; Loucks, D.P. Forecasting 3-D fish movement behavior using a Eulerian–Lagrangian–agent method (ELAM). Ecol. Model. 2006, 192, 197–223. [Google Scholar] [CrossRef]
- Fu, J.J.; Li, J.; An, R.D.; Mao, X.; Yi, W.M. Study of creating vertical slot fishway flow field based on swimming ability of Schizothorax Prenanti. J. Sichuan Univ. Eng. Sci. Ed. 2013, 45, 12–17. [Google Scholar]
- Sanz-Ronda, F.J.; Ruiz-Legazpi, J.; Bravo-Córdoba, F.J.; Makrakis, S.; Castro-Santos, T. Sprinting performance of two Iberian fish: Luciobarbus bocagei and Pseudochondrostoma duriense in an open channel flume. Ecol. Eng. 2015, 83, 61–70. [Google Scholar] [CrossRef]
- Goerig, E.; Bergeron, N.; Castro-Santos, T. Swimming behaviour and ascent paths of brook trout in a corrugated culvert. River Res. Appl. 2017, 33, 1463–1471. [Google Scholar] [CrossRef]
- Xiao, G.; Cheng, Z.P.; Mao, J.F. Semantic Models of Fish Behavior and Water Quality Warning; Science Press: Beijing, China, 2018; p. 31. (In Chinese) [Google Scholar]
- Li, L.; Nagy, M.; Graving, J.M.; Bak-Coleman, J.; Xie, G.; Couzin, I.D. Vortex phase matching as a strategy for schooling in robots and in fish. Nat. Commun. 2020, 11, 5408. [Google Scholar] [CrossRef]
- Liao, J.C. The role of the lateral line and vision on body kinematics and hydrodynamic preference of rainbow trout in turbulent flow. J. Exp. Biol. 2006, 209, 4077–4090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteiza, P.; Odstrcil, I.; Lauder, G.; Portugues, R.; Engert, F. A novel mechanism for mechanosensory-based rheotaxis in larval zebrafish. Nature 2017, 547, 445–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flow Region | Velocity (m/s) | TKE (m2·s−2) | RSSxy | RSSxz | RSSyz | SR | Gradvx | Gradvy |
|---|---|---|---|---|---|---|---|---|
| (N·m−2) | (N·m−2) | (N·m−2) | (s−1) | (s−1) | (s−1) | |||
| Low | 0.14–0.38 | 0.004–0.08 | −5.00–0.50 | −4.35–3.50 | −3.00–2.75 | 0.07–8.00 | −0.31–0.98 | −0.28–0.29 |
| Medium | 0.33–0.78 | 0.01–0.26 | −8.50–31.00 | −9.00–21.00 | −8.00–4.80 | 0.08–8.50 | −0.85–0.22 | −0.58–0.61 |
| High | 0.86–1.32 | 0.02–0.16 | −14.00–16.00 | −1.98–13.00 | −1.19–10.00 | 0.70–6.50 | −1.44–0.43 | −3.31–0.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Chen, M.; Wu, W.; Li, J.; An, R. Differences in the Natural Swimming Behavior of Schizothorax prenanti Individual and Schooling in Spatially Heterogeneous Turbulent Flows. Animals 2023, 13, 1025. https://doi.org/10.3390/ani13061025
Li M, Chen M, Wu W, Li J, An R. Differences in the Natural Swimming Behavior of Schizothorax prenanti Individual and Schooling in Spatially Heterogeneous Turbulent Flows. Animals. 2023; 13(6):1025. https://doi.org/10.3390/ani13061025
Chicago/Turabian StyleLi, Minne, Min Chen, Weixiong Wu, Jia Li, and Ruidong An. 2023. "Differences in the Natural Swimming Behavior of Schizothorax prenanti Individual and Schooling in Spatially Heterogeneous Turbulent Flows" Animals 13, no. 6: 1025. https://doi.org/10.3390/ani13061025