
Effects of Environmental Enrichment on the Behavior of Octopus vulgaris in a Recirculating Aquaculture System
 , ,
, ,  ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Recirculating Aquaculture System (RAS)
2.3. Experimental Design
- -
- Basic (BAS), consisting of a blue, factory-like environment, with only social enrichment (contact with conspecifics and operator) and food (ad libitum feeding and live food);
- -
- Enriched (ENR) environment, with the presence of physical enrichment: substrate (sand), wall color (the walls were “naturalized” with beige-colored polypropylene panels); cognitive enrichment: the presence of seashells, stones, and plastic toys; social enrichment (contact with conspecifics and the operator) and food (ad libitum feeding and live food).
- -
- the total number of observations in the two environments;
- -
- the total number of observations in both sexes (regardless of the environment);
- -
- the number of weekly observations in the two environments;
- -
- the percentage of observations of various behaviors in each environment.
2.4. Growth Performance
2.5. Statistical Analyses
2.6. Ethics
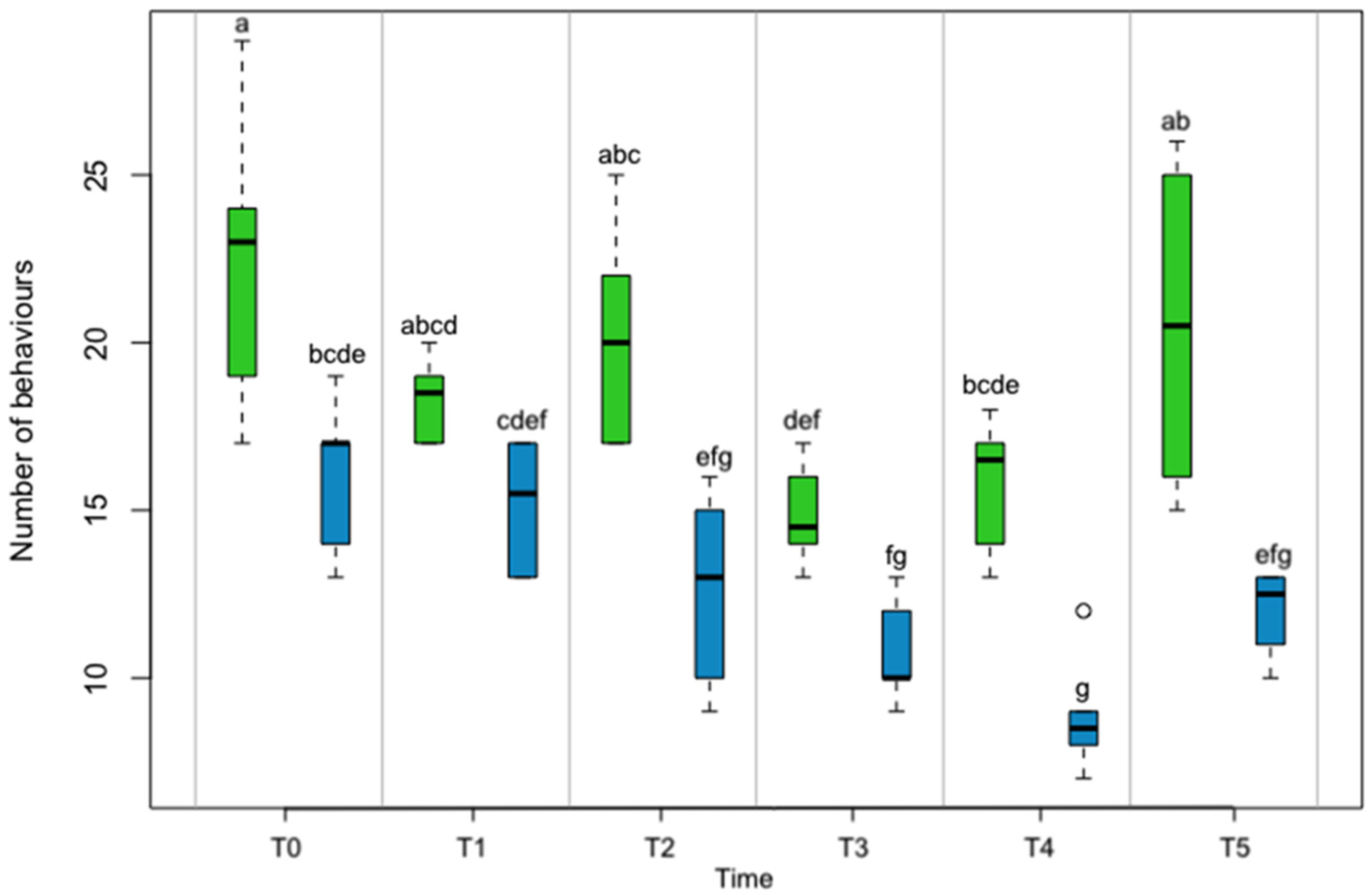
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iglesias, J.; Fuentes, L.; Villanueva, R. Octopus vulgaris. Paralarval Culture. In Cephalopod Culture; Iglesias, J., Fuentes, L., Villanueva, R., Eds.; Springer Dordrecht: New York, NY, USA; Berlin, Germany, 2014; pp. 427–450. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022—Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Sauer, W.H.H.; Gleadall, I.G.; Downey-Breedt, N.; Doubleday, Z.; Gillespie, G.; Haimovici, M.; Ibáñez, C.M.; Katugin, O.N.; Leporati, S.; Lipinski, M.R.; et al. World octopus fisheries. Rev. Fish. Sci. Aquac. 2021, 29, 279–429. [Google Scholar] [CrossRef]
- Lishchenko, F.; Perales-Raya, C.; Barret, C.; Oesterwind, D.; Power, A.M.; Larivain, A.; Laptikhovsky, V.; Karatza, A.; Badouvas, N.; Lishchenko, A.; et al. A review of recent studies on the life history and ecology of European cephalopods with emphasis on species with the greatest commercial fishery and culture potential. Fish. Res. 2021, 236, 105847. [Google Scholar] [CrossRef]
- Jereb, P.; Allcock, A.L.; Lefkaditou, E.; Piatkowski, U.; Hastie, L.C.; Pierce, G.J. Cephalopod biology and fisheries in Europe: II. Species Accounts. ICES Coop. Res. Rep. 2015, 325, 375. [Google Scholar]
- Vaz-Pires, P.; Seixas, P.; Barbosa, A. Aquaculture potential of the common octopus (Octopus vulgaris Cuvier, 1797): A review. Aquaculture 2004, 238, 221–238. [Google Scholar] [CrossRef]
- Mangold, K. Food, feeding and growth in Cephalopods. Mem. Natl. Mus. Vict. 1983, 44, 81–93. [Google Scholar] [CrossRef]
- Iglesias, J.; Sánchez, F.; Otero, J.; Moxica, C. Culture of octopus (Octopus vulgaris): Present knowledge, problems and perspectives. Cah. Opt. Méditerr. 2000, 47, 313–321. [Google Scholar]
- García, J.G.; Luaces, M.; Veiga, C.; Rey-Méndez, M. Farming costs and benefits, marketing details, investment risks: The case of Octopus vulgaris in Spain. In Cephalopod Culture; Iglesias, J., Fuentes, L., Villanueva, R., Eds.; Springer: Dordrecht, The Netherlands; New York, NY, USA; Berlin/Heidelberg, Germany, 2014; pp. 149–161. [Google Scholar]
- Vizcaíno, R.; Guardiola, F.A.; Prado-Alvarez, M.; Machado, M.; Costas, B.; Gestal, C. Functional and molecular immune responses in Octopus vulgaris skin mucus and haemolymph under stressful conditions. Aquac. Rep. 2023, 29, 101484. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Cabrera-Álvarez, M.J.; Maia, C.M.; Saraiva, J.L. Environmental enrichment in fish aquaculture: A review of fundamental and practical aspects. Rev. Aquac. 2022, 14, 704–728. [Google Scholar] [CrossRef]
- Martos-Sitcha, J.A.; Mancera, J.M.; Prunet, P.; Magnoni, L.J. Editorial: Welfare and Stressors in Fish: Challenges Facing Aquaculture. Front. Physiol. 2020, 11, 162. [Google Scholar] [CrossRef] [Green Version]
- Segner, H.; Reiser, S.; Ruane, N.; Rösch, R.; Steinhagen, D.; Vehanen, T. Welfare of Fishes in Aquaculture 2019; FAO: Budapest, Hungary, 2019. [Google Scholar]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Jones, N.A.R.; Webster, M.M.; Salvanes, A.G.V. Physical enrichment research for captive fish: Time to focus on the DETAILS. J. Fish Biol. 2021, 99, 704–725. [Google Scholar] [CrossRef] [PubMed]
- Sykes, A.V.; Galligioni, V.; Estefanell, J.; Hetherington, S.; Brocca, M.; Correia, J.; Ferreira, A.; Pieroni, E.M.; Fiorito, G. FELASA Working Group Report Capture and Transport of live cephalopods: Recommendations for scientific purposes. Lab. Anim. 2022. under review. [Google Scholar]
- Cooke, G.M.; Tonkins, B.M.; Mather, J.A. Care and Enrichment for Captive Cephalopods. In The Welfare of Invertebrate Animals; Carere, C., Mather, J., Eds.; Springer: Cham, Switzerland, 2019; pp. 179–208. [Google Scholar]
- Wells, D.L. Sensory stimulation as environmental enrichment for captive animals: A review. Appl. Anim. Behav. Sci. 2009, 118, 1–11. [Google Scholar] [CrossRef]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1195, 44, 229–243. [Google Scholar] [CrossRef]
- Young, R.J. Environmental Enrichment for Captive Animals; Wiley-Blackwell: Oxford, UK, 2003; p. 240. [Google Scholar]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2014, 17, 1–30. [Google Scholar] [CrossRef]
- Gerber, B.; Stamer, A.; Stadtlander, T. Environmental enrichment and its effect on fish welfare: Review. Excellence Sustain. 2015, 74. [Google Scholar]
- Alligood, C.A.; Dorey, N.R.; Mehrkam, L.R.; Leighty, N.R. Applying behavior-analytic methodology to the science and practice of environmental enrichment in zoos and aquariums. Zoo Biol. 2017, 36, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, E.A.; Richardson, R. Effects of environmental enrichment on rate of contextual processing and discriminative ability in adult rats. Neurobiol. Learn. Mem. 2000, 73, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Leggio, M.G.; Mandolesi, L.; Federico, F.; Spirito, F.; Ricci, B.; Gelfo, F.; Petrosini, L. Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the rat. Behav. Brain. Res. 2005, 163, 78–90. [Google Scholar] [CrossRef]
- Celli, M.L.; Tomonaga, M.; Udono, T.; Teramoto, M.; Nagano, K. Tool use task as environmental enrichment for captive chimpanzees. Appl. Anim. Behav. Sci. 2003, 81, 171–182. [Google Scholar] [CrossRef]
- Deguchi, Y.; Tokunaga, M.; Yamamoto, A.; Takahashi, S.; Ono, Y.; Maruyama, M.; Kimura, K.; Tsujimoto, T.; Iwase, T. Seasonal variation of effects of environmental enrichments for captive Asiatic black bears (Ursus thibetanus). Anim. Behav. Manag. 2008, 44, 159–165. [Google Scholar]
- Ojelade, O.C.; Durosaro, S.O.; Akinde, A.O.; Abdulraheem, I.; Oladepo, M.B.; Sopein, C.A.; Bhadmus, A.S.; Olateju, M. Environmental enrichment improves the growth rate, behavioral and physiological response of juveniles of Clarias gariepinus under laboratory conditions. Front. Vet. Sci. 2022, 9, 980364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Fu, Y.; Zhao, H.; Zhang, X. Social enrichment affects fish growth and aggression depending on fish species: Applications for aquaculture. Front. Mar. Sci. 2022, 9, 1011780. [Google Scholar] [CrossRef]
- De Pasquale, C.; Kemerer, N.; White, N.; Yost, M.; Wolfkill, J.; Sturgill, J.; Li, X. The influence of an enriched environment in enhancing recognition memory in zebrafish (Danio rerio). Front. Vet. Sci. 2021, 8, 749746. [Google Scholar]
- Lee, C.J.; Paull, G.C.; Tyler, C.R. Effects of environmental enrichment on survivorship, growth, sex ratio and behaviour in laboratory maintained zebrafish Danio rerio. J. Fish. Biol. 2019, 94, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Rosburg, A.J.; Fletcher, B.L.; Barnes, M.E.; Treft, C.E.; Bursell, B.R. Vertically suspended environmental enrichement structures improve the growth of juvenile landlocked fall chinook Salmon. Int. J. Innov. Stud. Aquat. Biol. Fish. 2019, 5, 17–24. [Google Scholar]
- Brunet, V.; Kleiber, A.; Patinote, A.; Sudan, P.L. Positive welfare effects of physical enrichments from the nature, functions and feeling-based approaches in farmed rainbow trout (Oncorhynchus mykiss). Aquaculture 2022, 550, 737825. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Diaz-Gil, C.; Saraiva, J.L.; Moranta, D.; Castanheira, M.F.; Nuñez-Velázqueza, S.; Ledesma-Corvi, S.; Mora-Ruiz, M.R.; Grau, A. Effects of structural environmental enrichment on welfare of juvenile seabream (Sparus aurata). Aquacult. Rep. 2019, 15, 100224. [Google Scholar] [CrossRef]
- Tonkins, B.M.; Tyers, A.M.; Cooke, G.M. Cuttlefish in captivity: An investigation into housing and husbandry for improving welfare. Appl. Anim. Behav. Sci. 2015, 168, 77–83. [Google Scholar] [CrossRef]
- Dickel, L.; Boal, J.G.; Budelmann, B.U. The effect of early experience on learning and memory in cuttlefish. Dev. Psychobiol. 2000, 36, 101–110. [Google Scholar] [CrossRef]
- Yasumuro, H.; Ikeda, Y. Environmental enrichment affects the ontogeny of learning, memory, and depth perception of the pharaoh cuttlefish Sepia pharaonis. Zool. 2018, 128, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Yasumuro, H.; Ikeda, Y. Environmental enrichment accelerates the ontogeny of cryptic behavior in pharaoh cuttlefish (Sepia pharaonis). Zool. Sci. 2016, 33, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Boal, J.G. Behavioural research methods for octopuses and cuttlefishes. Vie Milieu 2011, 61, 203–210. [Google Scholar]
- Beigel, M.; Boal, J.G. The effect of habitat enrichment on the mudflat octopus. Shape Enrich. 2006, 15, 3–6. [Google Scholar]
- Yasumuro, H.; Ikeda, Y. Effects of environmental enrichment on the behavior of the tropical octopus Callistoctopus aspilosomatis. Mar. Freshwater. Behav. Physiol. 2011, 44, 143–157. [Google Scholar] [CrossRef]
- Anderson, R.C.; Wood, J.B. Enrichment for giant Pacific octopuses: Happy as a clam? J. Appl. Anim. Welf. Sci. 2001, 4, 57–168. [Google Scholar] [CrossRef]
- Vergara-Ovalle, F.; Ayala-Guerrero, F.; Rosas, C.; Sanchez-Castillo, H. Novel object recognition in Octopus maya. Anim. Cognit. 2023, 26, 1065–1072. [Google Scholar] [CrossRef]
- Borrelli, L.; Fiorito, G. Behavioral analysis of learning and memory in cephalopods. In Learning Theory and Behaviour; Byrne, J.H., Ed.; Elsevier: Oxford, UK, 2008; pp. 605–628. [Google Scholar]
- Fiorito, G.; Scotto, P. Observational learning in Octopus vulgaris. Science 1992, 256, 545–547. [Google Scholar] [CrossRef] [Green Version]
- Tricarico, E.; Borrelli, L.; Gherardi, F.; Fiorito, G. I know my neighbour: Individual recognition in Octopus vulgaris. PLoS ONE 2011, 6, e18710. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, L. Testing the Contribution of Relative Brain Size and Learning Capabilities on the Evolution of Octopus vulgaris and Other Cephalopods. Ph.D. Thesis, Open University, London, UK.
- Fiorito, G.; Gherardi, F. Prey-handling behaviour of Octopus vulgaris (Mollusca, Cephalopoda) on bivalve preys. Behav. Process. 1999, 46, 75–88. [Google Scholar] [CrossRef]
- Nixon, M.; Maconnachie, E. Drilling by Octopus vulgaris (Mollusca: Cephalopoda) in the Mediterranean. J. Zool. Lond. 1988, 216, 687–716. [Google Scholar] [CrossRef]
- Fiorito, G.; von Planta, C.; Scotto, P. Problem solving ability in Octopus vulgaris Lam. (Mollusca, Cephalopoda). Behav. Neural Biol. 1990, 53, 217–230. [Google Scholar] [CrossRef] [PubMed]
- O’brien, C.E.; Di Miccoli, V.; Fiorito, G. A preliminary investigation of the response of Octopus vulgaris to experimental stimuli in the wild. J. Molluscan Stud. 2021, 87, eyab032. [Google Scholar]
- Richter, J.N.; Hochner, B.; Kuba, M.J. Pull or push? Octopuses solve a puzzle problem. PLoS ONE 2016, 11, e0152048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuba, M.J.; Meisel, D.V.; Byrne, R.A.; Griebel, U.; Mather, J.A. Looking at play in Octopus vulgaris. Berliner Palaontol. Abh. 2003, 19, 426–438. [Google Scholar]
- Kuba, M.J.; Byrne, R.A.; Meisel, D.V.; Mather, J.A. When do Octopus play? Effects of repeated testing, object type, age and food deprivation on object play in Octopus vulgaris. J. Comp. Psychol. 2006, 120, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, R.T.; Messenger, J.B. Cephalopod Behaviour, 2nd ed.; Cambridge University Press: Cambridge, UK, 2018; p. 376. [Google Scholar]
- Packard, A.; Sanders, G.D. Body patterns of Octopus vulgaris and maturation of the response to disturbance. Anim. Behav. 1971, 19, 780–790. [Google Scholar] [CrossRef]
- Ikeda, Y. Color Change in Cephalopods. In Pigments, Pigment Cells and Pigment Patterns; Hashimoto, H., Goda, M., Futahashi, R., Kelsh, R., Akiyama, T., Eds.; Springer: Singapore, 2021; pp. 425–449. [Google Scholar]
- Boyle, P.; Rodhouse, P. Physiological Ecology. In Cephalopods: Ecology and fisheries; Boyle, P., Rodhouse, P., Eds.; Wiley-Blackwell: Ames, IA, USA, 2005; p. 452. [Google Scholar]
- Bertapelle, C.; Polese, G.; Di Cosmo, A. Enriched environment increases PCNA and PARP1 levels in Octopus vulgaris central nervous system: First evidence of adult neurogenesis in lophotrochozoan. J. Exp. Zool. 2017, 328, 347–359. [Google Scholar] [CrossRef]
- Martins, C.I.; Galhardo, L.; Noble, C.; Damsgård, B.; Spedicato, M.T.; Zupa, W.; Beauchaud, M.; Kulczykowska, E.; Massabuau, J.C.; Carter, T.; et al. Behavioural indicators of welfare in farmed fish. Fish Physiol. Biochem. 2012, 38, 17–41. [Google Scholar] [CrossRef] [Green Version]
- Dawkins, M.S. Behaviour as a tool in the assessment of animal welfare. Zool 2003, 106, 383–387. [Google Scholar] [CrossRef]
- Fiorito, G.; Affuso, A.; Basil, J.; Cole, A.; De Girolamo, P.; D’Angelo, L.; Dickel, L.; Gestal, C.; Grasso, F.; Kuba, M.; et al. Guidelines for the Care and Welfare of Cephalopods in Research—A consensus based on an initiative by CephRes, FELASA and the Boyd Group. Lab. Anim. 2015, 49, 1–90. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, L.; Gherardi, F.; Fiorito, G. A Catalogue of Body Patterning in Cephalopoda; Firenze Univerity Press, Stazione Zoologica A. Dohrn: Firenze, Italy, 2006; p. 622. [Google Scholar]
- Casalini, A.; Roncarati, A.; Emmanuele, P.; Guercilena, N.; Bonaldo, A.; Parma, L.; Mordenti, O. Evaluation of reproductive performances of the common octopus (Octopus vulgaris) reared in water recirculation systems and fed different diets. Sci. Rep. 2020, 10, 15261. [Google Scholar] [CrossRef] [PubMed]
- Cowdry, E.V. The Colour Changes of Octopus Vulgaris Lmk; University Library, The Librarian: Toronto, ON, Canada, 1911; p. 53. [Google Scholar]
- Packard, A.; Sanders, G.D. What the octopus shows to the world. Endeavour 1969, 28, 92–99. [Google Scholar] [PubMed]
- Messenger, J.B. Cephalopod chromatophores: Neurobiology and natural history. Biol. Rev. 2001, 76, 473–528. [Google Scholar] [CrossRef] [PubMed]
- Packard, A. The behaviour of Octopus vulgaris. Bull. Inst. Oceanogr. 1963, 1, 35–49. [Google Scholar]
- Mather, J.A.; Anderson, R.C. Personalities of octopuses (Octopus rubescens). J. Comp. Psychol. 1993, 107, 336. [Google Scholar] [CrossRef]
- Carere, C.; Grignani, G.; Bonanni, R.; Della Gala, M.; Carlini, A.; Angeletti, D.; Mather, J.A. Consistent individual differences in the behavioural responsiveness of adult male cuttlefish (Sepia officinalis). Appl. Anim. Behav. Sci. 2015, 167, 89–95. [Google Scholar] [CrossRef]
- McLean, E. Fish tank color: An overview. Aquaculture 2021, 530, 735–750. [Google Scholar] [CrossRef]
- Okamoto, M.; Anraku, K.; Kawamura, G.; Tanaka, Y. Selectivity of color of shelter by Octopus vulgaris and O. aegina under different background colors. Nippon Suisan Gakkaishi 2001, 67, 672–677. [Google Scholar] [CrossRef]
- McLean, E. Background color and cultured invertebrates–A review. Aquaculture 2021, 537, 736523. [Google Scholar] [CrossRef]
- Batzina, A.; Kalogiannis, D.; Dalla, C.; Papadopoulou-Daifoti, Z.; Chadio, S.; Karakat-souli, N. Blue substrate modifies the time course of stress response ingilthead seabream Sparus aurata. Aquaculture 2014, 420, 247–253. [Google Scholar] [CrossRef]
- Batzina, A.; Dalla, C.; Papadopoulou-Daifoti, Z.; Karakatsouli, N. Effects of environmental enrichment on growth, aggressive behaviour and brain monoamines of gilthead seabream Sparus aurata reared under different social conditions. Comp. Biochem. Physiol. 2014, 169, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Batzina, A.; Karakatsouli, N. Is it the blue gravel substrate or only its blue color that improves growth and reduces aggressive behavior of gilthead seabream Sparus aurata? Aquac. Eng. 2014, 62, 49–53. [Google Scholar] [CrossRef]
- Opiyo, M.A.; Ngugi, C.C.; Rasowo, J. Combined effects of stocking density and background colour on growth performance and survival of Nile tilapia (Oreochromis niloticus, L.) fry reared in aquaria. J. Fish. Sci. 2014, 8, 228–237. [Google Scholar] [CrossRef]
- Wang, J.; Peng, K.; Lu, H.; Li, R.; Song, W.; Liu, L.; Wang, H.; Wang, C.; Shi, C. The effect of tank colour on growth performance, stress response and carapace colour of juvenile swimming crab Portunus trituberculatus. Aquac. Res. 2019, 50, 2735–2742. [Google Scholar] [CrossRef]
- Volpato, G.L.; Duarte, C.R.A.; Luchiari, A.C. Environmental color affects Nile tilapia reproduction. Braz. J. Med. Biol. Res. 2004, 37, 479–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mather, J.A. Sand digging in Sepia officinalis: Assessment of a Cephalopod mollusc’s “fixed” behavior pattern. J. Comp. Psychol. 1986, 100, 315. [Google Scholar] [CrossRef]
- Kang, D.Y.; Kim, H.C.; Myeong, J.I.; Min, B.H. Effect of infiltrating substrate on growth and double-sided pigmentation of flounder (Paralichthys olivaceus) fry. KOSFAS 2014, 47, 406–412. [Google Scholar]
- Ottesen, O.H.; Noga, E.J.; Sandaa, W. Effect of substrate on progression and healing of skin erosions and epidermal papillomas of Atlantic halibut, Hippoglossus hippoglossus (L.). J. Fish Dis. 2007, 30, 43–53. [Google Scholar] [CrossRef]
- Mather, J.A.; Dickel, L. Cephalopod complex cognition. Curr. Opin. Behav. Sci. 2017, 16, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Batzina, A.; Karakatsouli, N. The presence of substrate as a means of environmental enrichment in intensively reared gilthead seabream Sparus aurata: Growth and behavioral effects. Aquaculture 2012, 370, 54–60. [Google Scholar] [CrossRef]
- Rosengren, M.; Kvingedal, E.; Näslund, J.; Johnsson, J.I.; Sundell, K. Born to be wild: Effects of rearing density and environmental enrichment on stress, welfare, and smolt migration in hatchery-reared Atlantic salmon. Can. J. Fish. Aquat. Sci. 2016, 74, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, F.J.; Valverde, C.J.; Garcìa, B. Octopus vulgaris: Ongrowing. In Cephalopod Culture; Iglesias, J., Fuentes, L., Villanueva, R., Eds.; Springer Dordrecht: New York, NY, USA; Berlin, Germany, 2014; pp. 451–466. [Google Scholar]
- Rodriguez-González, T.; Valverde, J.C.; Sykes, A.V.; García, B.J. Performance of raw material thermal treatment on formulated feeds for common octopus (Octopus vulgaris) ongrowing. Aquaculture 2015, 442, 37–43. [Google Scholar] [CrossRef]
- Etim, N.N.; Williams, M.E.; Evans, E.I.; Offiong, E.E. Physiological and behavioural responses of farm animals to stress: Implications to animal productivity. Am. J. Adv. Agric. Res. 2013, 1, 53–61. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
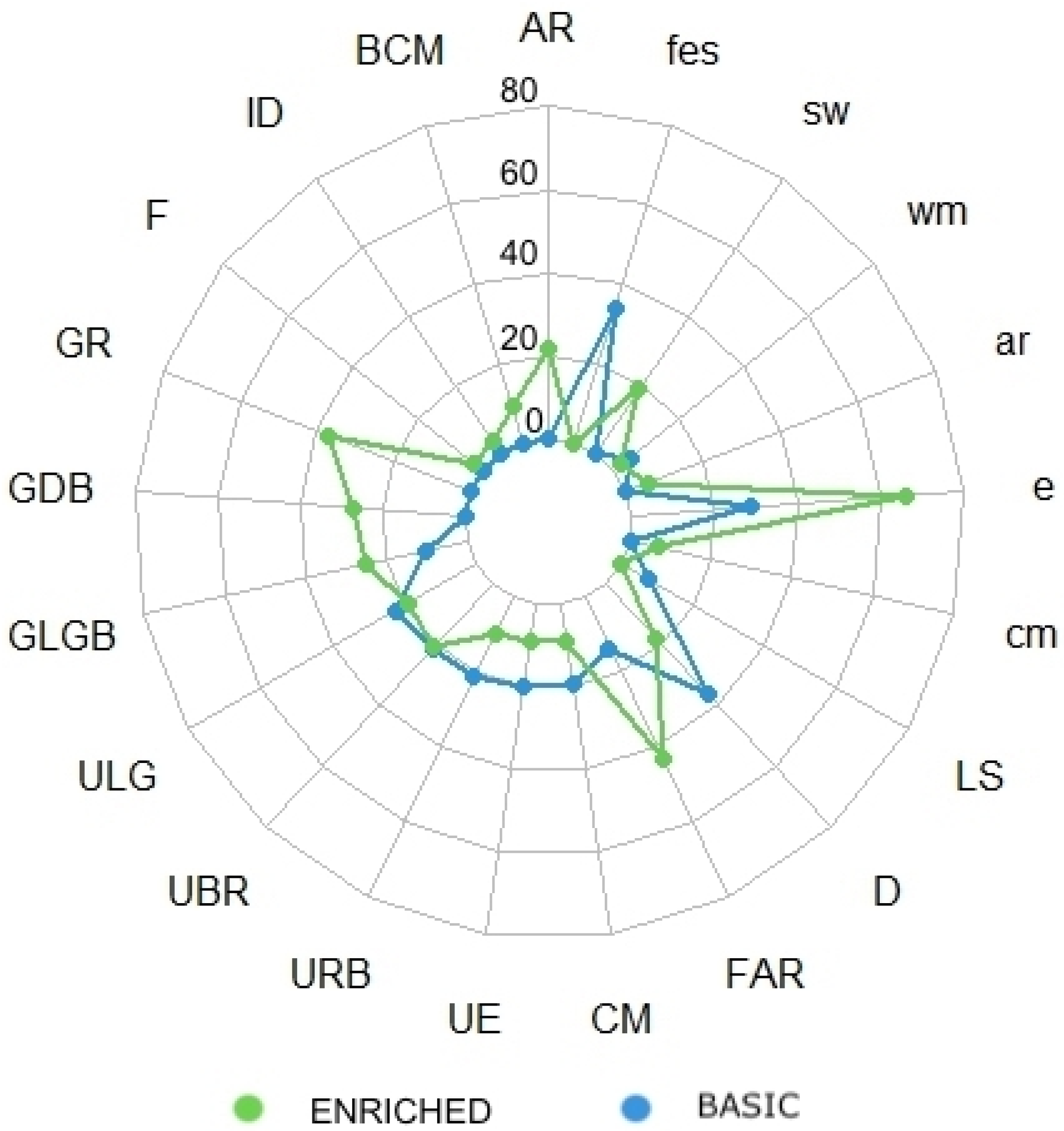
| N | Body Pattern | Meaning | BAS | ENR | BAS% | ENR% | References |
| 1 | Conflict Mottle—CM | Disturbance | ✓ | ✓ | 11.3 | 3.4 | [56,65] |
| 2 | Unilateral Effect—UE | Disturbance | ✓ | ✓ | 11.5 | 3.4 | [56] |
| 3 | Uniform Brownish- Red—UBR | Intraspecific encounters, disturbance | ✓ | ✓ | 12.4 | 8.2 | [65] |
| 4 | Uniform Reddish- Brown—URB | Social interactions, disturbance | ✓ | ✓ | 12.4 | 3.9 | [65] |
| 5 | Full Attack Response—FAR | Feeding behavior | ✓ | ✓ | 7.9 | 16.9 | [63] |
| 6 | Denning—D | Rest, feeding, disturbance | ✓ | ✓ | 21.2 | 7.1 | [55] |
| 7 | Uniform Light Gray—ULG | Camouflage | ✓ | ✓ | 13.0 | 7.4 | [65] |
| 8 | Ground Light Grayish- brown—GLGB | Camouflage, rest | ✓ | ✓ | 6.0 | 9.8 | [65] |
| 9 | Longitudinal Stripes—LS | Intraspecific interactions, disturbance | ✓ | ✗ | 4.3 | - | [56,65] |
| 10 | Fighting—F | Intraspecific interactions | ✗ | ✓ | - | 1.2 | [63] |
| 11 | Incomplete Dymantic—ID | Disturbance | ✗ | ✓ | - | 1.5 | [56] |
| 12 | Broad Conflict Mottle—BCM | Disturbance | ✗ | ✓ | - | 3.6 | [66] |
| 13 | Acute Resemblance—AR | Camouflage | ✗ | ✓ | - | 8.5 | [67] |
| 14 | Ground Dark Brown—GDB | Camouflage, rest | ✗ | ✓ | - | 10.7 | [65] |
| 15 | General Resemblance—GR | Camouflage | ✗ | ✓ | - | 14.4 | [63,67] |
| N | Components | Meaning | BAS | ENR | BAS% | ENR% | References |
| 1 | Envelope—e | Prey capture | ✓ | ✓ | 42.7 | 66.4 | [63] |
| 2 | Withdrawal Manoeuvre—wm | Defensive posture | ✓ | ✓ | 7.3 | 2.3 | [56] |
| 3 | Funnel Directed Toward External Stimulus—fes | Reaction against the disturbance | ✓ | ✗ | 50 | - | [56] |
| 4 | Cleaning Manoeuvre—cm | Rapid twirling of the arms | ✗ | ✓ | - | 6.9 | [56] |
| 5 | Arms Raised—ar | Postural component | ✗ | ✓ | - | 5.7 | [68] |
| 6 | Swimming—sw | Locomotor component | ✗ | ✓ | - | 18.7 | [66] |
| Animal | Wi (g) | Wf (g) | SGR (%) | AGR (g/d) | |
|---|---|---|---|---|---|
| ENR | EF1 | 731.7 | 1354.8 | 1.47 | 14.8 |
| EF2 | 689.3 | 1270.8 | 1.46 | 13.8 | |
| EF3 | 764.1 | 1371.4 | 1.39 | 14.5 | |
| Mean F | 728.4 ± 37.5 | 1332. 3 ± 53.9 | 1.44 ± 0.04 | 14.4 ± 0.5 | |
| EM1 | 705.4 | 1374.1 | 1.59 | 15.9 | |
| EM2 | 758.8 | 1418.4 | 1.49 | 15.7 | |
| EM3 | 725.7 | 1454.2 | 1.65 | 17.3 | |
| Mean M | 730 ± 27 | 1415.6 ± 40.1 | 1.58 ± 0.1 | 16.3 ± 0.9 | |
| MEAN ENR | 729.2 ± 29.2 | 1373.9 ± 62.3 ** | 1.51 ± 0.1 ** | 15.4 ± 1.2 ** | |
| BAS | PF1 | 728.7 | 911.4 | 0.53 | 4.4 |
| PF2 | 675.6 | 862.7 | 0.58 | 4.5 | |
| PF3 | 752.8 | 907.1 | 0.44 | 3.7 | |
| Mean F | 719 ± 39.5 | 893.7 ± 27 | 0.52 ± 0.1 | 4.2 ± 0.4 | |
| PM1 | 778.3 | 921.4 | 0.40 | 3.4 | |
| PM2 | 814.2 | 968.9 | 0.41 | 3.7 | |
| PM3 | 733.6 | 851.4 | 0.35 | 2.8 | |
| Mean M | 775.4 ± 40.4 | 913.9 ± 59.1 | 0.39 ± 0.03 | 3.3 ± 0.4 | |
| MEAN BAS | 747.2 ± 47.2 | 903.8 ± 42.5 | 0.5 ± 0.1 | 3.7 ± 0.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casalini, A.; Gentile, L.; Emmanuele, P.; Brusa, R.; Elmi, A.; Parmeggiani, A.; Galosi, L.; Roncarati, A.; Mordenti, O. Effects of Environmental Enrichment on the Behavior of Octopus vulgaris in a Recirculating Aquaculture System. Animals 2023, 13, 1862. https://doi.org/10.3390/ani13111862
Casalini A, Gentile L, Emmanuele P, Brusa R, Elmi A, Parmeggiani A, Galosi L, Roncarati A, Mordenti O. Effects of Environmental Enrichment on the Behavior of Octopus vulgaris in a Recirculating Aquaculture System. Animals. 2023; 13(11):1862. https://doi.org/10.3390/ani13111862
Chicago/Turabian StyleCasalini, Antonio, Laura Gentile, Pietro Emmanuele, Riccardo Brusa, Alberto Elmi, Albamaria Parmeggiani, Livio Galosi, Alessandra Roncarati, and Oliviero Mordenti. 2023. "Effects of Environmental Enrichment on the Behavior of Octopus vulgaris in a Recirculating Aquaculture System" Animals 13, no. 11: 1862. https://doi.org/10.3390/ani13111862