
Activity Patterns of Captive Red Panda (Ailurus fulgens)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Observation Methods
2.3. Data Analysis
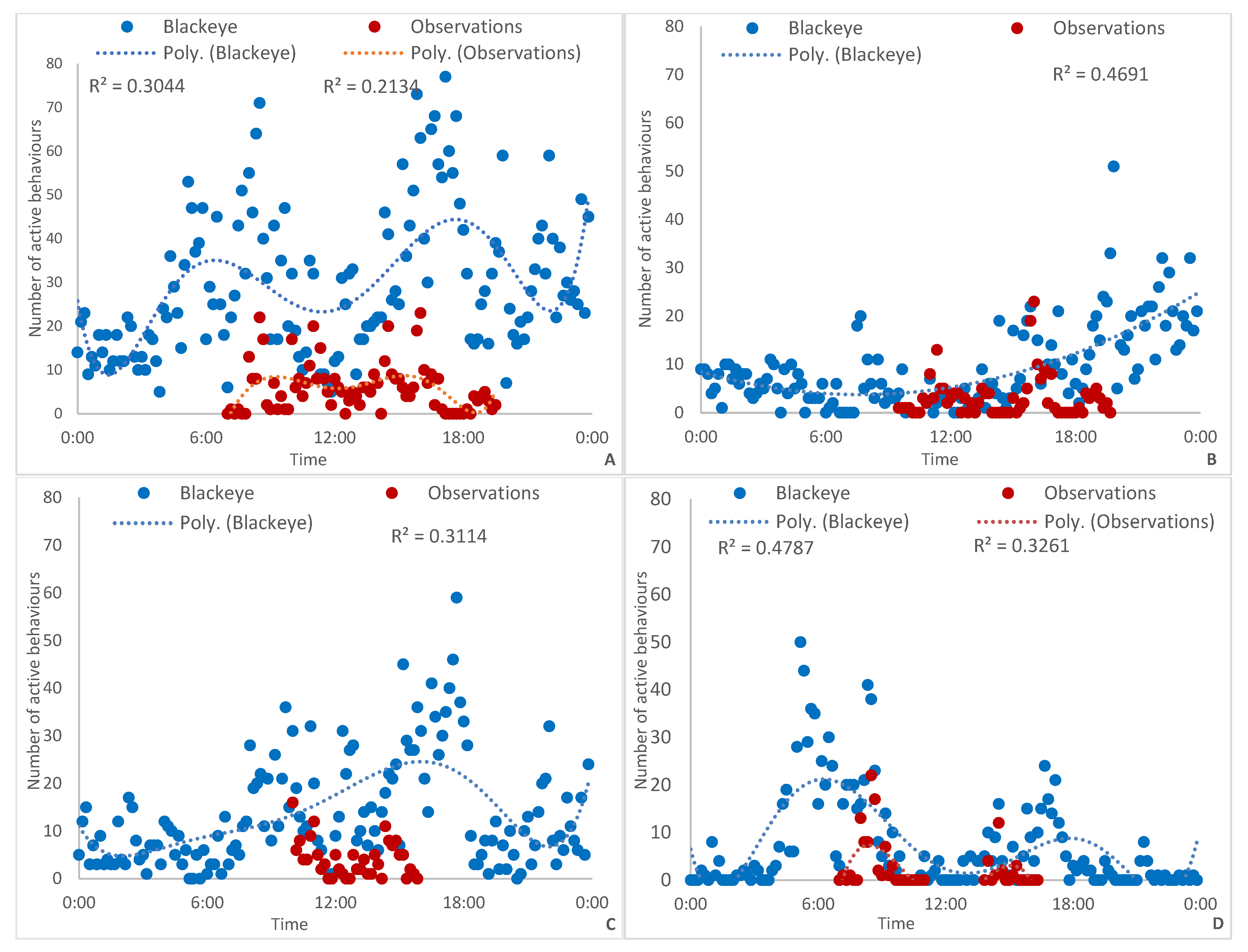
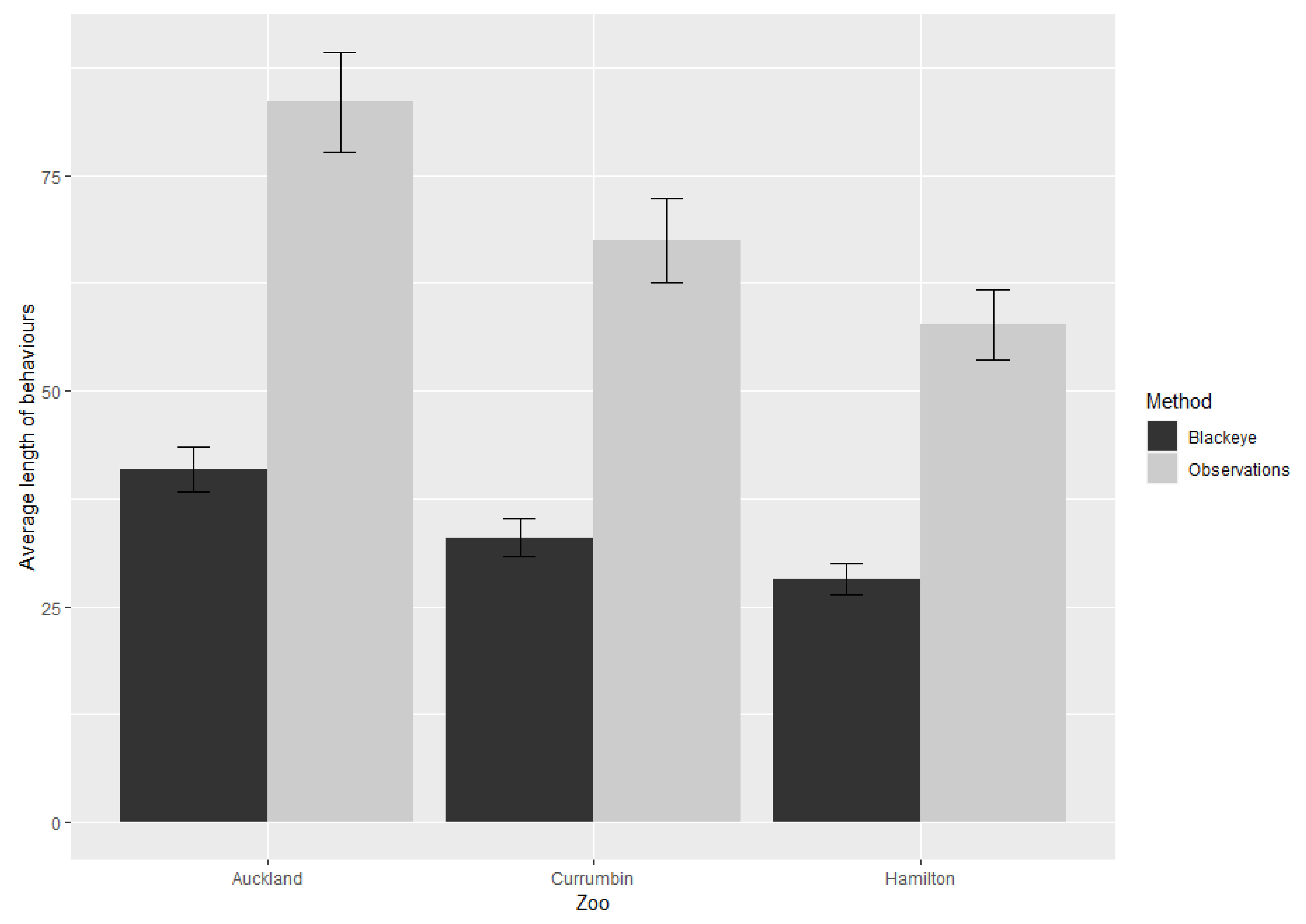
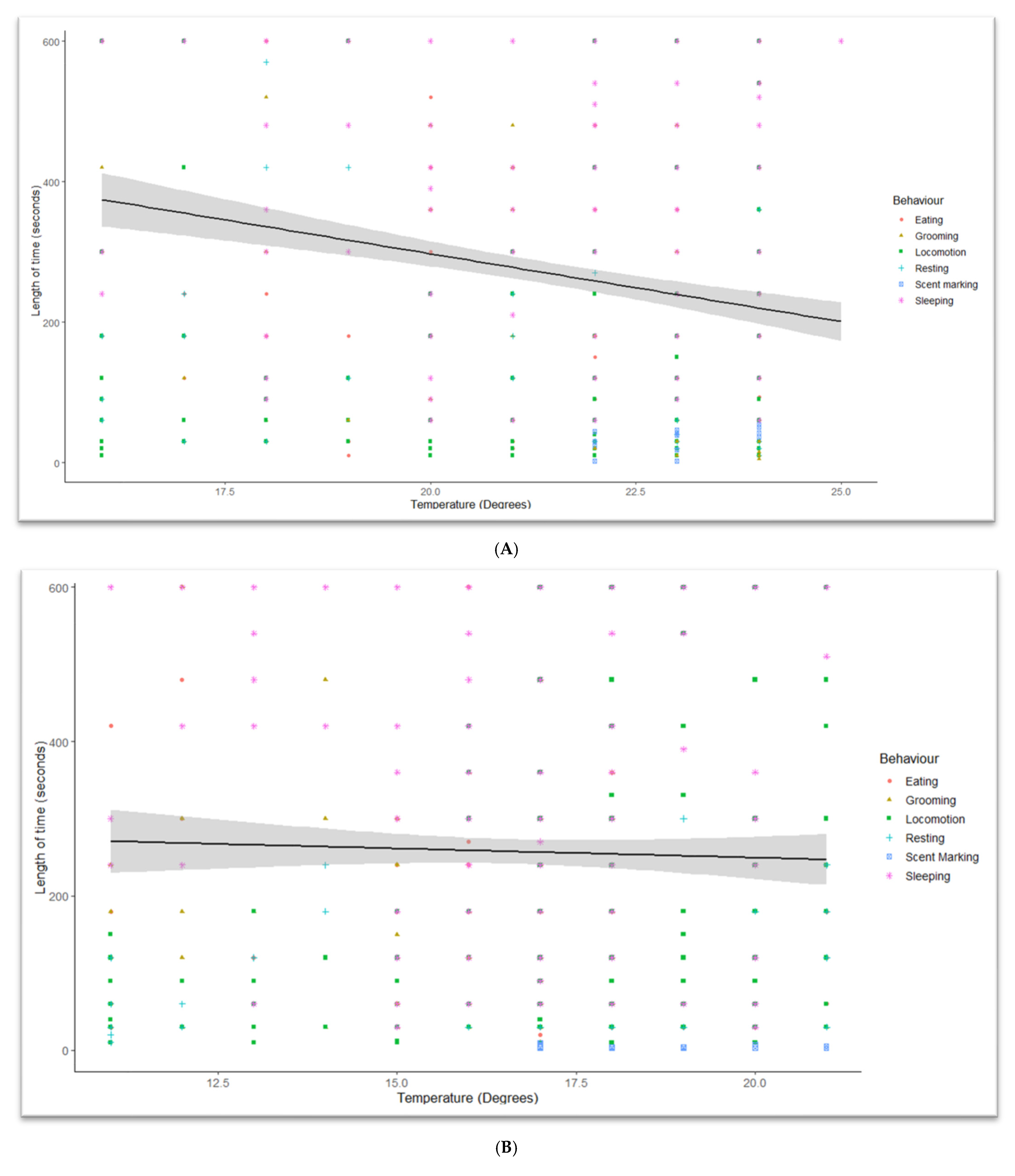
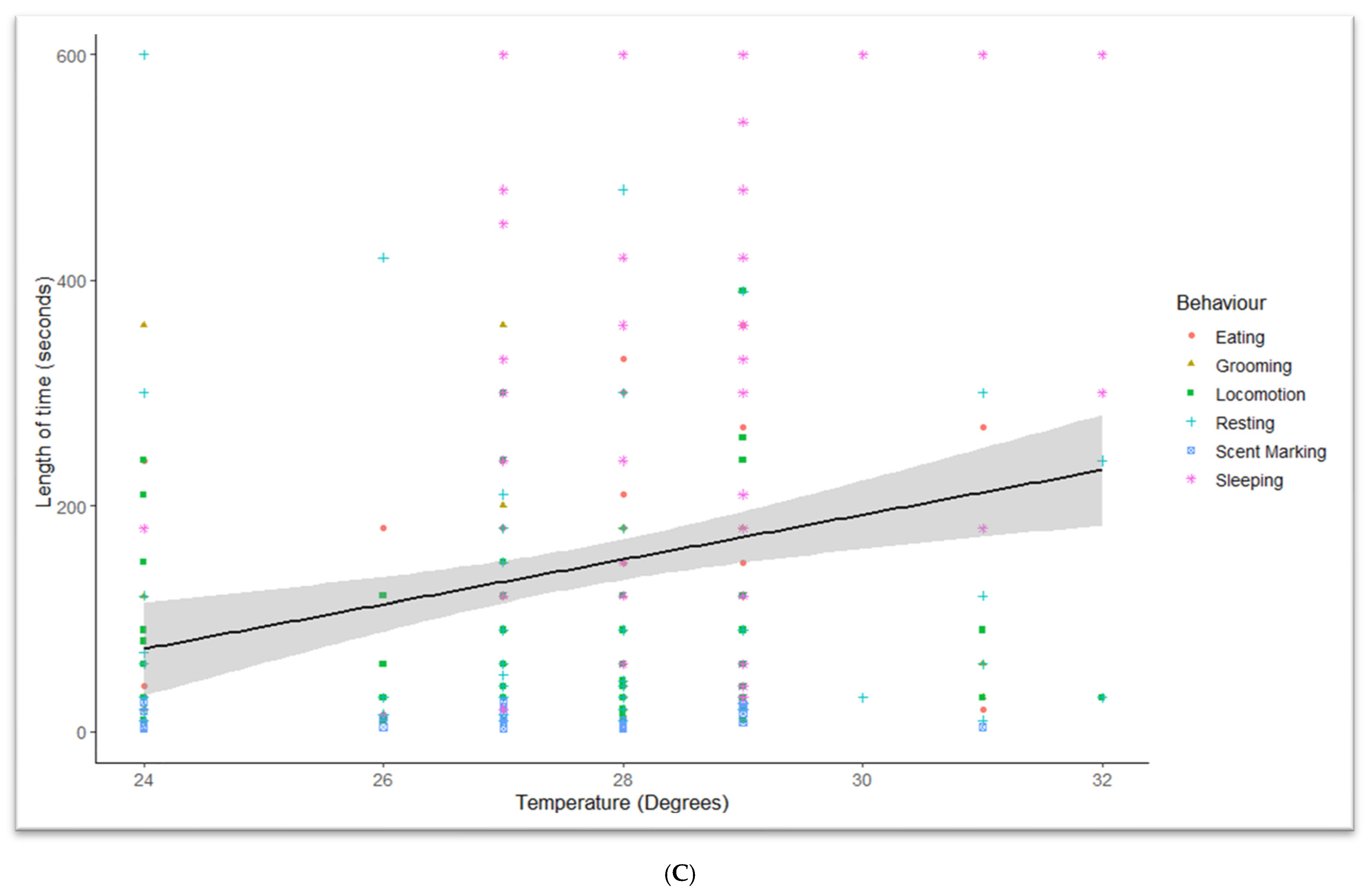
3. Results
3.1. Active Behaviours
3.2. All Behaviours
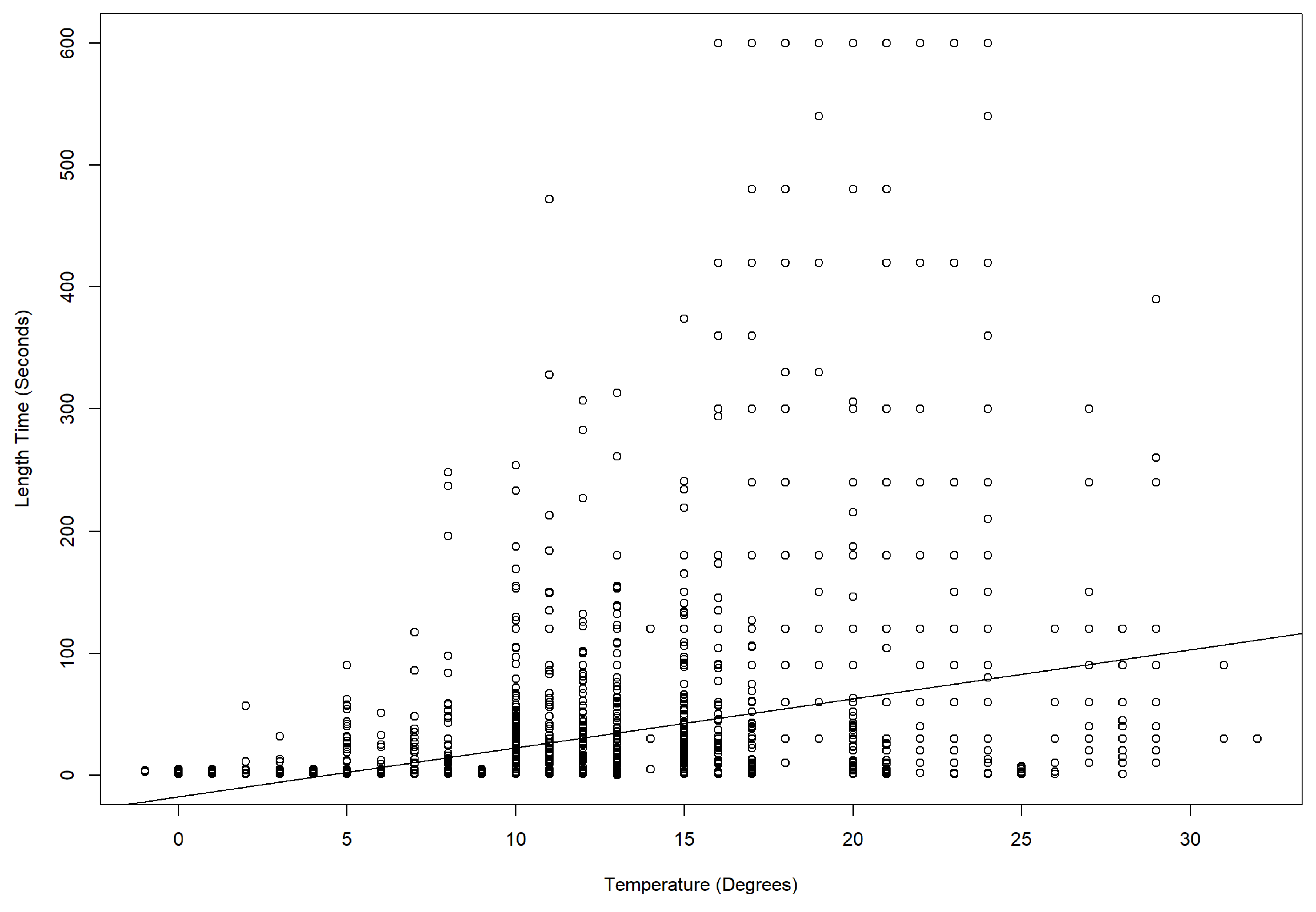
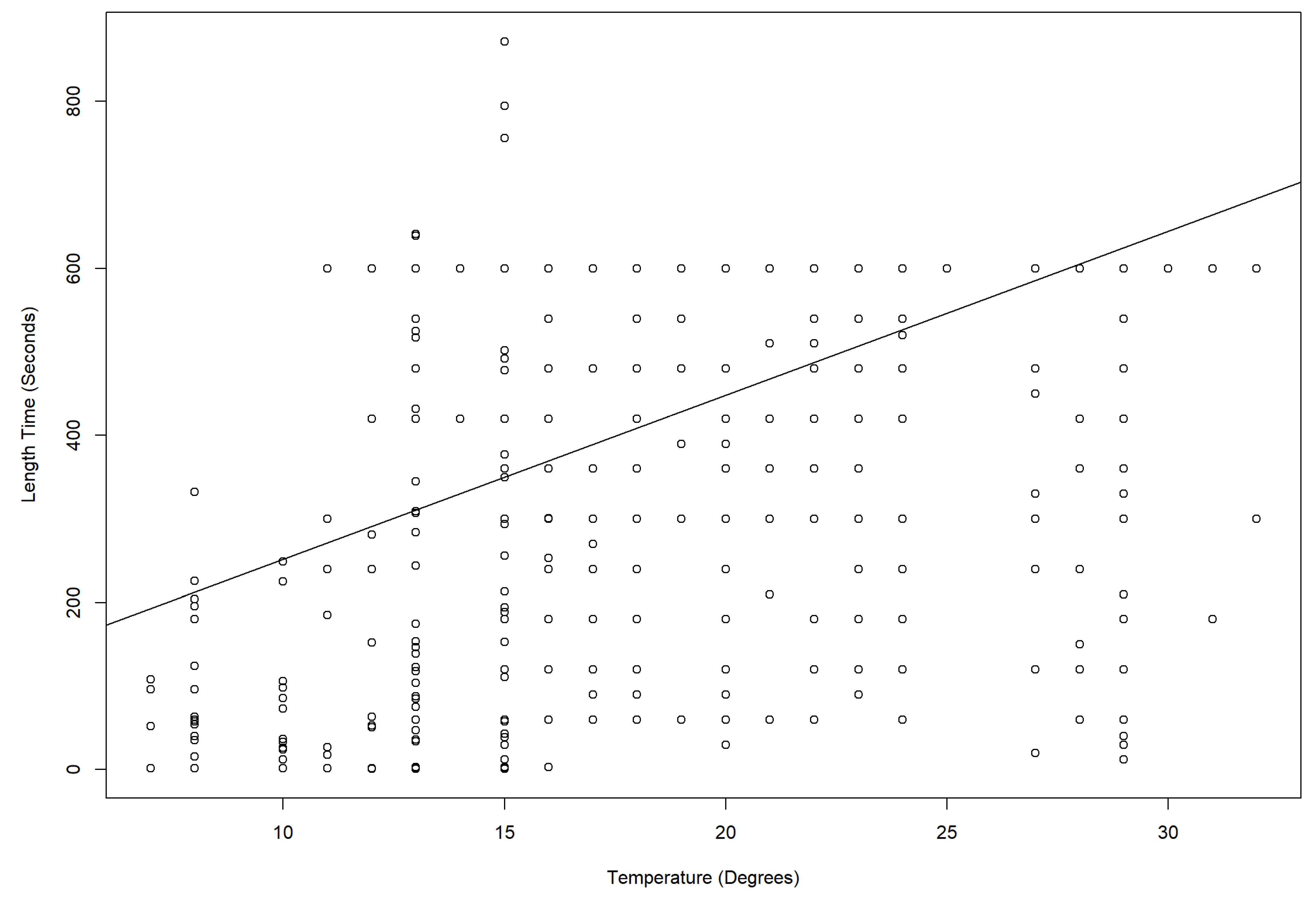
3.3. Temperature Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glatston, A.; Wei, F.; Than, Z.; Sherpa, A. Ailurus fulgens (errata versions published in 2017). The IUCN Red List of Threatened Species 2015. Available online: https://www.iucnredlist.org/species/714/110023718 (accessed on 20 October 2021).
- CITES. Lesser Panda. Available online: https://cites.org/eng/gallery/species/mammal/lesser_panda.html (accessed on 20 October 2021).
- Choudhury, A. An overview of the status and conservation of the red panda Ailurus fulgens in India, with reference to its global status. Oryx 2001, 35, 250–259. [Google Scholar] [CrossRef]
- Groves, C. The Taxonomy and Phylogeny of Ailurus. In Red Panda: Biology and Conservation of the First Panda, 1st ed.; Glatston, A.R., Ed.; Elsevier Inc.: London, UK, 2011; pp. 101–124. [Google Scholar] [CrossRef]
- Princée, F.P.; Glatston, A.R. Influence of climate on the survivorship of neonatal red pandas in captivity. Zoo Biol. 2016, 35, 104–110. [Google Scholar] [CrossRef] [PubMed]
- AZA Small Carnivore TAG. Red Panda Care Manual. Association of Zoos and Aquariums, Silver Spring. Available online: https://assets.speakcdn.com/assets/2332/red_panda_care_manual_2012.pdf (accessed on 25 October 2021).
- Glatston, A.R. Synthesis. In Red Panda: Biology and Conservation of the First Panda, 1st ed.; Glatston, A.R., Ed.; Elsevier Inc.: London, UK, 2011; pp. 447–467. [Google Scholar] [CrossRef]
- Eriksson, P.; Zidar, J.; White, D.; Westander, J.; Andersson, M. Current husbandry of red pandas (Ailurus fulgens) in zoos. Zoo Biol. 2010, 29, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Leus, K. The Global Captive Population of the Red Panda—Possibilities for the Future. In Red Panda: Biology and Conservation of the First Panda, 1st ed.; Glatston, A.R., Ed.; Elsevier Inc.: London, UK, 2011; pp. 335–356. [Google Scholar] [CrossRef]
- Hu, Y.; Thapa, A.; Wei, F. Ailurus fulgens (Himalayan Red Panda) and Ailurus styani (Chinese Red Panda). Trends Genet. 2020, 36, 624–625. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, K. Red Panda Husbandry for Reproductive Management. In Red Panda: Biology and Conservation of the First Panda, 1st ed.; Glatston, A.R., Ed.; Elsevier Inc.: London, UK, 2011; pp. 231–255. [Google Scholar] [CrossRef]
- Roberts, M.S.; Gittleman, J.L. Ailurus fulgens. Mamm. Species 1984, 222, 1–8. [Google Scholar] [CrossRef]
- Lama, S.; Shrestha, S.; Koju, N.P.; Sherpa, A.P.; Tamang, M. Assessment of the Impacts of Livestock Grazing on Endangered Red Panda (Ailurus fulgens) Habitat in Eastern Nepal. Open J. Ecol. 2020, 10, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Thapa, A.; Hu, Y.; Aryal, P.C.; Singh, P.B.; Shah, K.B.; Wei, F. The endangered red panda in Himalayas: Potential distribution and ecological habitat associates. Glob. Ecol. 2020, 21, e00890. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, J.; Han, Z.; Wei, F. Activity patterns of wild red pandas in Fengtongzhai Nature Reserve, China. Ital. J. Zool. 2011, 78, 398–404. [Google Scholar] [CrossRef]
- Reid, D.G.; Jinchu, H.; Yan, H. Ecology of the red panda Ailurus fulgens in the Wolong Reserve, China. J. Zool. 1991, 225, 347–364. [Google Scholar] [CrossRef]
- Johnson, K.G.; Schaller, G.B.; Jinchu, H. Comparative behavior of red and giant pandas in the Wolong Reserve, China. J. Mammal. 1988, 69, 552–564. [Google Scholar] [CrossRef]
- Yonzon, P.B.; Hunter, M.L.; Hunter, M.L. jnr Cheese, Tourists, and Red Pandas in the Nepal Himalayas. Conserv. Biol. 1991, 5, 196–202. [Google Scholar] [CrossRef]
- Wei, F.; Zhang, Z. Red Panda Ecology. In Red Panda: Biology and Conservation of the First Panda, 1st ed.; Glatston, A.R., Ed.; Elsevier Inc.: London, UK, 2011; pp. 193–212. [Google Scholar] [CrossRef]
- Andres-Bray, T.C.; Moller, P.; Powell, D.M. Preliminary model of personality structure in captive red pandas (Ailurus fulgens). J. Zoo Aqua. Res. 2020, 8, 29–36. [Google Scholar] [CrossRef]
- Krebs, B.L.; Torres, E.; Chesney, C.; Moon, V.K.; Watters, J.V. Applying Behavioral Conditioning to Identify Anticipatory Behaviors. J. App. Ani. Welf. Sci. 2017, 20, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Conover, G.K.; Gittlemen, J.L. Scent-marking in captive red pandas (Ailurus fulgens). Zoo Biol. 1989, 8, 193–205. Available online: https://psycnet.apa.org/doi/10.1002/zoo.1430080210 (accessed on 26 November 2022). [CrossRef]
- Bugler, K. Monitoring the ‘original’ panda: Impacts and outcomes of using infra-red trail cameras on captive red panda (Ailurus fulgens) behaviour; Lincoln University: Lincoln, New Zealand, 2021; Available online: https://researcharchive.lincoln.ac.nz/handle/10182/14171 (accessed on 26 November 2021).
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 26 November 2021).
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means_. R package version 1.8.3. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 26 November 2021).
- Lama, S.T. Assessing mammal diversity, distribution, and abundance: Piloting arboreal camera trapping as a tool for monitoring endangered red panda in temperate forest of Eastern Nepal; Lincoln University: Lincoln, New Zealand, 2018; Available online: https://hdl.handle.net/10182/12053 (accessed on 10 September 2020).
- Bista, D.; Baxter, G.S.; Hudson, N.J.; Lama, S.T.; Weerman, J.; Murray, P.S. Movement and dispersal of a habitat specialist in human-dominated landscapes: A case study of the red panda. Move Ecol. 2021, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Bista, D.; Baxter, G.S.; Hudson, N.J.; Lama, S.T.; Murray, P.J. Effect of disturbances and habitat fragmentation on an arboreal habitat specialist mammal using GPS telemetry: A case of the red panda. Landsc. Ecol. 2022, 37, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Spiexio, C.; Altamura, M.; Weerman, J.; Regaiolli, B. Behaviour of zoo-housed red panda (Ailurus fulgens): A case-study testing the behavioural variety index. J. Zool. Bot. Gard. 2022, 3, 223–237. [Google Scholar] [CrossRef]
- Khan, A.; Baskaran, N.; Rai, U. Drivers of psychological stress among captive red panda (Ailurus fulgens) in north-eastern India. Proceedings of Student Conference on Conservation Sciences at Indian Institute of Science, Bangalore, India, 21–24 September 2017. [Google Scholar] [CrossRef]
- Blasetti, A.; Boitani, L.; Riviello, M.C.; Visalberghi, E. Activity budgets and use of enclosed space by wild boars (Sus scrofa) in captivity. Zoo Biol. 1988, 7, 69–79. [Google Scholar] [CrossRef]
- Leus, K.; Weerman, J. The role of ex situ management in the conservation of red panda. In Red Panda: Biology and Conservation of the First Panda, 2nd ed.; Glatston, A.R., Ed.; Academic press: Cambridge, MA, USA, 2022; pp. 289–309. [Google Scholar] [CrossRef]
- Zidar, J. Keeping red pandas in captivity. SLU, Dept. of Animal Environment and Health, Skara, Sweden. 2008. Available online: https://stud.epsilon.slu.se/10802/ (accessed on 26 November 2021).
- Kapplehof, J.; Weerman, J. The development of the Red panda Ailurus fulgens EEP: From a failing captive population to a stable population that provides effective support to in situ conservation. Int. Zoo. Yb. 2020, 54, 102–112. [Google Scholar] [CrossRef]
- Dechanupong, J. Maternal Behavior and Time Allocation of Red Panda (Ailurus fulgens) in Captivity. School of Integrative Studies, George Mason University, United States of America. 2019. Available online: https://hdl.handle.net/1920/11515 (accessed on 8 September 2020).
- Loeffler, K. Management, Husbandry and Veterinary Medicine of Red Pandas Living ex situ in China. In Red Panda: Biology and Conservation of the First Panda, 1st ed.; Glatston, A.R., Ed.; Elsevier Inc.: London, UK, 2011; pp. 323–333. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zoo | Panda Details | Enclosure Details | Enclosure Size |
|---|---|---|---|
| Auckland Zoo | 2.1.0 Ramesh, male, 9 years Khela, female, 6 years Tashi, male, 4 months | Lots of large natural trees, most of which are not native to their natural habitat. Large pond (empty due to juvenile). One main covered platform where feeding occurred. Two nest boxes hidden from public view and an off-display den. | 170 m2 |
| Hamilton Zoo | 1.2.0 Chito, male, 16 years Taylor, female, 10 years Jamuna, female, 4 years | Multiple large exotic tree species, large (empty) pond, a series of off-ground walkways and ramps interconnecting four covered platforms, and a visible large den. One tall man-made tower. A large bamboo stand wrapped around the back half of the perimeter. | 529 m2 |
| Currumbin Wildlife Sanctuary | 1.0.0 Pasang, male, 16 years | Three main platforms at differing heights, interconnected via off-ground walkways. One platform had a roof and the other was fully enclosed, where food and water were given. Natural trees were small. Large off-display den with resting areas and water access. Several water misters. | 117 m2 |
| Behaviour | Explanation of Behaviour |
|---|---|
| Locomotion | Movement, on all four paws, that would shift an individual from one area to another, such as by running, walking, or climbing |
| Resting | Lying, sitting, or standing, eyes open, responsive to surroundings, staying in one area |
| Sleeping | Lying, curled in a ball or flat, eyes closed, unresponsive to surrounding noise or activity |
| Eating | Selection and chewing of food brought by keepers, usually fruit, bamboo, or pellets (grass or leaves already in the enclosure may also be consumed) |
| Drinking | Licking up water in an enclosure, either from an artificial bowl or stream |
| Grooming/Scratching | Repeated licking or chewing motions of fur or quick paw movements, with claw, across own body |
| Scent marking | Frequent rubbing of genitals and/or urination on objects around the enclosure |
| Defecation | Urination, or passing of bowel movement (usually at a latrine site) |
| Playing | Spontaneous actions that are voluntary and internally motivated. These actions are not associated with the direct need for survival, e.g., eating or predator avoidance. Can be one or more, but is always non-aggressive |
| Interaction | Between two panda, overall non-aggressive |
| Keeper interaction | Non-aggressive actions towards a keeper, such as taking food from a keeper or sniffing a keeper’s boots |
| In den | Out of sight while in the den |
| Aggressive behaviour | Vocalisation, staring, or aggressive displays towards a conspecific or keeper. Aggressive displays include standing on hind paws with forepaws raised above the head, head bobbing, and slamming forepaws on ground (this behaviour was not seen during the study) |
| Behaviour | Observation | Camera | Zoo |
|---|---|---|---|
| Defecation | 0.24% | 2.34% | Auckland |
| 0.14% | 5.71% | Hamilton | |
| 0% 0.17% | 0% 1.65% | CWS All zoos | |
| Eating | 6.84% | 28.50% | Auckland |
| 3.93% | 21.66% | Hamilton | |
| 7.86% | 0.37% | CWS | |
| 5.89% | 12.02% | All zoos | |
| Grooming | 4.86% | 0.63% | Auckland |
| 6.9% | 1.71% | Hamilton | |
| 4.49% | 5.57% | CWS | |
| 5.6% | 3.69% | All zoos | |
| Interaction | 0% | 0.01% | Auckland |
| 0.33% | 0.95% | Hamilton | |
| 0% | 0% | CWS | |
| 0.16% | 0.16% | All zoos | |
| Locomotion | 13.99% | 33.04% | Auckland |
| 11.57% | 49.78% | Hamilton | |
| 11.32% | 6.63% | CWS | |
| 12.83% | 21.66% | All zoos | |
| Playing | 0.04% | 3.05% | Auckland |
| 0% | 0% | Hamilton | |
| n/a | n/a | CWS | |
| 0% | 0.9% | All zoos | |
| Resting | 6.8% | 19.66% | Auckland |
| 2.85% | 18.78% | Hamilton | |
| 20.35% | 10.75% | CWS | |
| 7.3% | 15% | All zoos | |
| Scent marking | 0.32% | 0.08% | Auckland |
| 0.21% | 0.92% | Hamilton | |
| 0.41% | 0.47% | CWS | |
| 0.21% | 0.45% | All zoos | |
| Sleeping | 66.92% | 12.59% | Auckland |
| 74.27% | 0.35% | Hamilton | |
| 41.91% | 76.19% | CWS | |
| 67.66% | 44.47% | All zoos |
| Zoo | Average | Minimum | Maximum | Season |
|---|---|---|---|---|
| Auckland Zoo | 22.77 °C | 18 °C | 25 °C | Early Autumn (March) |
| Hamilton Zoo | 15.5 °C | 11 °C | 18 °C | Mid-Autumn (April/May) |
| CWS | 27.49 °C | 23 °C | 32 °C | Early Summer (December) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bugler, K.A.; Ross, J.G.; Paterson, A.M. Activity Patterns of Captive Red Panda (Ailurus fulgens). Animals 2023, 13, 846. https://doi.org/10.3390/ani13050846
Bugler KA, Ross JG, Paterson AM. Activity Patterns of Captive Red Panda (Ailurus fulgens). Animals. 2023; 13(5):846. https://doi.org/10.3390/ani13050846
Chicago/Turabian StyleBugler, Kathryn A., James G. Ross, and Adrian M. Paterson. 2023. "Activity Patterns of Captive Red Panda (Ailurus fulgens)" Animals 13, no. 5: 846. https://doi.org/10.3390/ani13050846