
Evaluation of a Model (RUMINANT) for Prediction of DMI and CH4 from Tropical Beef Cattle
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Area of Study
2.2. Experimental Design
2.3. Forage Diets
- Toledo grass (Brachiaria brizantha cv. CIAT 26110);
- Cayman grass (Brachiaria hybrid cv. CIAT BR 02/1752);
- Star grass (Cynodon plectostachius) and tropical kudzu (Pueraria phaseoloides) in a 70:30 ratio;
- Cayman grass and Leucaena (Leucaena diversifolia) in a 70:30 ratio;
- Cayman grass and Leucaena (Leucaena leucocephala) in a 70:30 ratio;
- Toledo grass, Leucaena (L. diversifolia), and Canavalia (Canavalia brasiliensis) in a 70:15:15 ratio.
2.4. Animals
2.5. Determination of Intake
2.6. Gas Measurement
2.7. Simulation of Intake and Methane Emissions
2.8. Predictive Capability of the RUMINANT Model
2.9. Sensitivity Analysis
3. Results
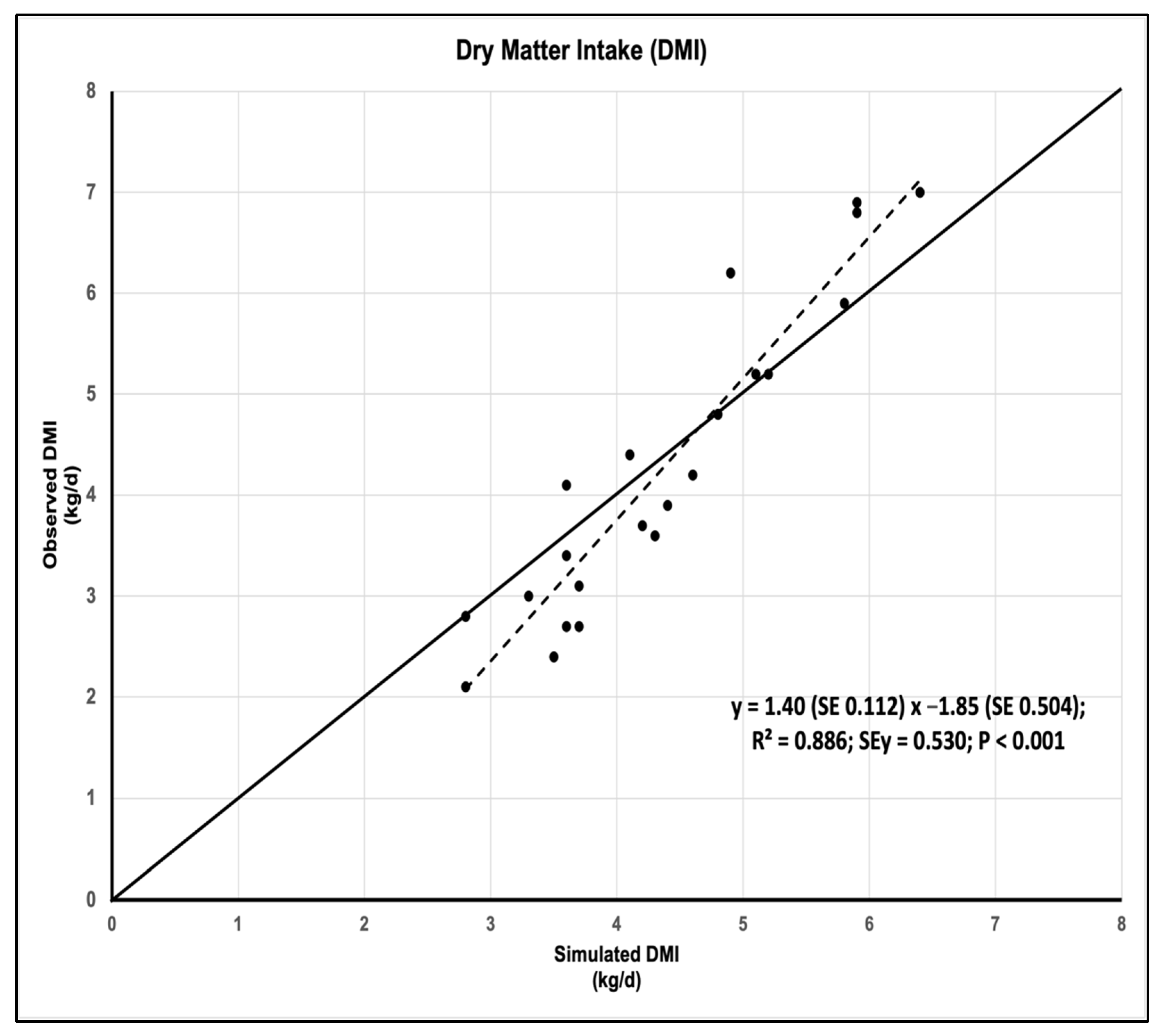
3.1. Accuracy and Precision of DMI Simulation
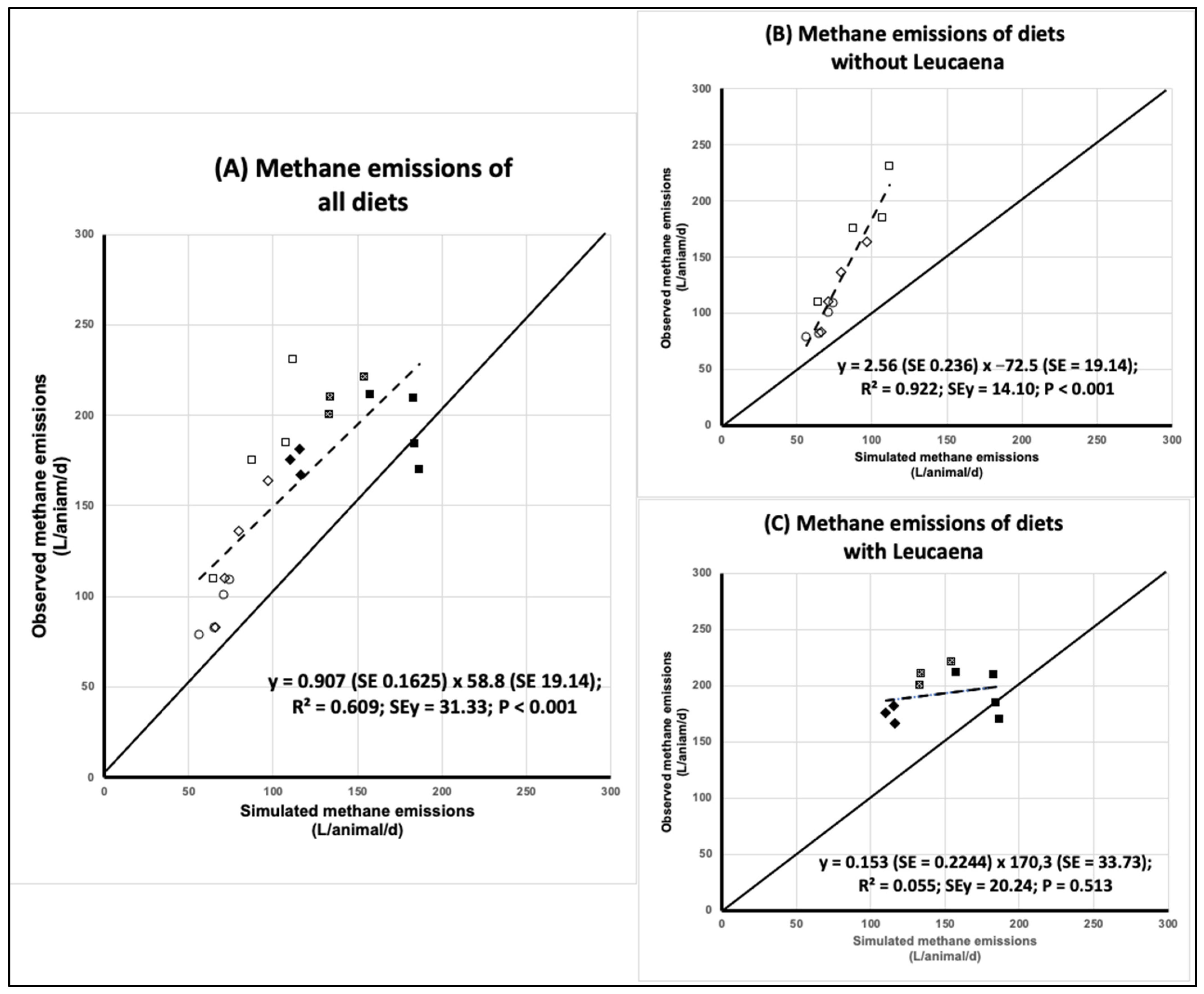
3.2. Simulation of Methane Emissions
3.2.1. Accuracy
3.2.2. Precision
3.2.3. Combined Accuracy and Precision
3.3. Sensitivity Analysis of the RUMINANT Model to Forage Quality Values
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tedeschi, L.O.; Abdalla, A.L.; Álvarez, C.; Anuga, S.W.; Arango, J.; Beauchemin, K.A.; Becquet, P.; Berndt, A.; Burns, R.; De Camillis, C. Quantification of methane emitted by ruminants: A review of methods. J. Anim. Sci. 2022, 100, skac197. [Google Scholar]
- Dent, J.B.; Blackie, M.J. Systems Simulation in Agriculture; Applied Science Publishers: London, UK, 1980. [Google Scholar]
- Ellis, J.; Kebreab, E.; Odongo, N.; McBride, B.; Okine, E.; France, J. Prediction of methane production from dairy and beef cattle. J. Dairy Sci. 2007, 90, 3456–3466. [Google Scholar]
- Ramin, M.; Huhtanen, P. Development of equations for predicting methane emissions from ruminants. J. Dairy Sci. 2013, 96, 2476–2493. [Google Scholar]
- Hristov, A.N.; Kebreab, E.; Niu, M.; Oh, J.; Bannink, A.; Bayat, A.R.; Boland, T.M.; Brito, A.F.; Casper, D.P.; Crompton, L.A.; et al. Symposium review: Uncertainties in enteric methane inventories, measurement techniques, and prediction models. J. Dairy Sci. 2018, 101, 6655–6674. [Google Scholar] [CrossRef]
- Kebreab, E.; Clark, K.; Wagner-Riddle, C.; France, J. Methane and nitrous oxide emissions from Canadian animal agriculture: A review. Can. J. Anim. Sci. 2006, 86, 135–157. [Google Scholar]
- Banco Mundial; CIPAV; CIAT; Fedegán; Fondo Acción; TNC. Acción de Mitigación Nacionalmente Apropiada NAMA de la ganadería Bovina Sostenible en Colombia. 2021. Available online: https://cgspace.cgiar.org/bitstream/handle/10568/114670/Reporte%20NAMA%2008_10_2021%20final.pdf?sequence=1 (accessed on 25 September 2022).
- Herrero, M.; Thornton, P.K.; Kruska, R.; Reid, R.S. Systems dynamics and the spatial distribution of methane emissions from African domestic ruminants to 2030. Agric. Ecosyst. Environ. 2008, 126, 122–137. [Google Scholar]
- Herrero, M.; Havlík, P.; Valin, H.; Notenbaert, A.; Rufino, M.C.; Thornton, P.K.; Blümmel, M.; Weiss, F.; Grace, D.; Obersteiner, M. Biomass use, production, feed efficiencies, and greenhouse gas emissions from global livestock systems. Proc. Natl. Acad. Sci. USA 2013, 110, 20888–20893. [Google Scholar]
- Herrero, M.; Fawcett, R.H.; Jessop, N.S. Predicting intake and nutrient supply of tropical and temperate diets for ruminants using a simple dynamic model of digestion. In Bioparametrics Ruminant Nutrition Reference Laboratories Monograph; Institute of Ecology and Resource Management, University of Edinburgh: Edinburgh, UK, 2002. [Google Scholar]
- Mendes, L.B.; Herrero, M.; Havlík, P.; Mosnier, A.; Balieiro, S.; Moreira, R.; Obersteiner, M. Simulation of enteric methane emissions from individual beef cattle in tropical pastures of improving quality: A case study with the model Ruminant. Adv. Anim. Biosci. 2016, 7, 233–234. [Google Scholar]
- Illius, A.; Gordon, I. Prediction of intake and digestion in ruminants by a model of rumen kinetics integrating animal size and plant characteristics. J. Agric. Sci. 1991, 116, 145–157. [Google Scholar]
- Analla, M. Model validation through the linear regression fit to actual versus predicted values. Agric. Syst. 1998, 57, 115–119. [Google Scholar]
- Tedeschi, L.O. Assessment of the adequacy of mathematical models. Agric. Syst. 2006, 89, 225–247. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar]
- Bradstreet, R.B. The Kjeldahl digestion. In The Kjeldahl Method for Organic Nitrogen; Bradstreet, R.B., Ed.; Academic Press: Cambridge, MA, USA, 1965; pp. 9–88. [Google Scholar] [CrossRef]
- Horwitz, W.; Albert, R.; Deutsch, M.J.; Thompson, N.J. Precision parameters of methods of analysis required for nutrition labeling. Part I. Major nutrients. J. Assoc. Off. Anal. Chem. 1990, 73, 661–680. [Google Scholar]
- Tilley, J.; Terry, R. A two stage for the in vitro digestion of forage crops. J. Br. Grassl. Soc 1963, 18, 104–111. [Google Scholar]
- Lockyer, D. Methane emissions from grazing sheep and calves. Agric. Ecosyst. Environ. 1997, 66, 11–18. [Google Scholar]
- Gasmet Tehcnologies, Oy. Global Solutions for Gas Monitoring. Available online: http://www.gasmet.com/ (accessed on 25 September 2018).
- Storm, I.M.; Hellwing, A.L.F.; Nielsen, N.I.; Madsen, J. Methods for measuring and estimating methane emission from ruminants. Animals 2012, 2, 160–183. [Google Scholar]
- Mayer, D.; Butler, D. Statistical validation. Ecol. Model. 1993, 68, 21–32. [Google Scholar]
- Neter, J.; Kutner, M.H.; Nachtsheim, C.J.; Wasserman, W. Applied Linear Statistical Models, 5th ed.; McGraw-Hill: Boston, MA, USA, 1996. [Google Scholar]
- Rice, J.A.; Cochran, P.A. Independent evaluation of a bioenergetics model for largemouth bass. Ecology 1984, 65, 732–739. [Google Scholar]
- Lin, L.I.-K. A concordance correlation coefficient to evaluate reproducibility. Biometrics 1989, 45, 255–268. [Google Scholar]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 11 August 2018).
- Hendricksen, R.; Minson, D. The feed intake and grazing behaviour of cattle grazing a crop of Lablab purpureus cv. Rongai. J. Agric. Sci. 1980, 95, 547–554. [Google Scholar]
- Jamieson, W.; Hodgson, J. The effects of variation in sward characteristics upon the ingestive behaviour and herbage intake of calves and lambs under a continuous stocking management. Grass Forage Sci. 1979, 34, 273–282. [Google Scholar]
- Gaviria-Uribe, X.; Castro-Montoya, J.; Bolívar, D.M.; Molina-Botero, I.; Chirinda, N.; Dickhoefer, U.; Barahona, R.; Arango, J. The effects of two species of Leucaena on in vitro rumen fermentation, methane production and post-ruminal protein supply in diets based on Urochloa hybrid cv. Cayman. Agronomy 2022, 12, 629. [Google Scholar]
- Molina, I.; Angarita, E.; Mayorga, O.; Chará, J.; Barahona-Rosales, R. Effect of Leucaena leucocephala on methane production of Lucerna heifers fed a diet based on Cynodon plectostachyus. Livest. Sci. 2016, 185, 24–29. [Google Scholar]
- Molina, I.; Donneys, G.; Montoya, S.; Rivera, J.; Villegas, G.; Chará, J.; Barahona, R. La inclusión de Leucaena leucocephala reduce la producción de metano de terneras Lucerna alimentadas con Cynodon plectostachyus y Megathyrsus maximus. Livest. Res. Rural Dev. 2015, 27, 1–8. [Google Scholar]
- Piñeiro-Vázquez, A.T.; Jiménez-Ferrer, G.O.; Chay-Canul, A.J.; Casanova-Lugo, F.; Díaz-Echeverría, V.F.; Ayala-Burgos, A.J.; Solorio-Sánchez, F.J.; Aguilar-Pérez, C.F.; Ku-Vera, J.C. Intake, digestibility, nitrogen balance and energy utilization in heifers fed low-quality forage and Leucaena leucocephala. Anim. Feed Sci. Technol. 2017, 228, 194–201. [Google Scholar]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar]
- Black, J.; Beever, D.; Faichney, G.; Howarth, B.; Graham, N.M. Simulation of the effects of rumen function on the flow of nutrients from the stomach of sheep: Part 1—Description of a computer program. Agric. Syst. 1981, 6, 195–219. [Google Scholar]
- Fagundes, G.M.; Benetel, G.; Welter, K.C.; Melo, F.A.; Muir, J.P.; Carriero, M.M.; Souza, R.L.; Meo-Filho, P.; Frighetto, R.T.; Berndt, A. Tannin as a natural rumen modifier to control methanogenesis in beef cattle in tropical systems: Friend or foe to biogas energy production? Res. Vet. Sci. 2020, 132, 88–96. [Google Scholar]
- Boadi, D.; Wittenberg, K.; McCaughey, W. Effects of grain supplementation on methane production of grazing steers using the sulphur (SF6) tracer gas technique. Can. J. Anim. Sci. 2002, 82, 151–157. [Google Scholar]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar]
- Hungate, R.E. The Rumen and its Microbes; Academic Press: New York, NY, USA, 1996. [Google Scholar]
- Jenkins, T. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar]
- Bauchop, T.; Elsden, S. The growth of micro-organisms in relation to their energy supply. Microbiology 1960, 23, 457–469. [Google Scholar]
- Hart, K.J.; Martin, P.; Foley, P.; Kenny, D.; Boland, T. Effect of sward dry matter digestibility on methane production, ruminal fermentation, and microbial populations of zero-grazed beef cattle. J. Anim. Sci. 2009, 87, 3342–3350. [Google Scholar]
- Nsahlai, I.; Apaloo, J. On the suitability of Illius and Gordon\’s model for simulating the intake and digestibility of roughage diets by ruminants. S. Afr. J. Anim. Sci. 2007, 37, 275–289. [Google Scholar]
- Chirinda, N.; Kracher, D.; Lægdsmand, M.; Porter, J.R.; Olesen, J.E.; Petersen, B.M.; Doltra, J.; Kiese, R.; Butterbach-Bahl, K. Simulating soil N2O emissions and heterotrophic CO2 respiration in arable systems using FASSET and MoBiLE-DNDC. Plant Soil 2011, 343, 139–160. [Google Scholar]
- Mauricio, R.M.; Ribeiro, R.S.; Paciullo, D.S.C.; Cangussú, M.A.; Murgueitio, E.; Chará, J.; Estrada, M.X.F. Silvopastoral systems in Latin America for biodiversity, environmental, and socioeconomic improvements. In Agroecosystem Diversity; Elsevier: Amsterdam, The Netherlands, 2019; pp. 287–297. [Google Scholar]
- Canul-Solis, J.; Campos-Navarrete, M.; Piñeiro-Vázquez, A.; Casanova-Lugo, F.; Barros-Rodríguez, M.; Chay-Canul, A.; Cárdenas-Medina, J.; Castillo-Sánchez, L. Mitigation of rumen methane emissions with foliage and pods of tropical trees. Animals 2020, 10, 843. [Google Scholar]



{kind=link}
{kind=link}
| Forage Diets | CP (%) | NDF (%) | NSC (%) | EE (%) | Ashes (%) | IVDMD (%) |
|---|---|---|---|---|---|---|
| Toledo Grass 1 | 6.5 | 69.2 | 11.4 | 2.51 | 10.5 | 64.1 |
| Cayman Grass 2 | 8.3 | 68.2 | 8.8 | 2.51 | 12.1 | 61.3 |
| Star Grass 3 plus Kudzu 4 (70:30) | 11.2 | 72.9 | 3.4 | 2.97 | 9.6 | 58.7 |
| Cayman Grass 2 plus Leucaena 5 (70:30) | 14.7 | 56.8 | 12.5 | 3.46 | 12.5 | 60.4 |
| Cayman Grass 2 plus Leucaena 6 (70:30) | 11.0 | 62.5 | 10.9 | 3.78 | 11.9 | 61.6 |
| Toledo Grass 1 plus Canavalia 7 plus Leucaena 5 (70:15:15) | 10.8 | 66.5 | 10.2 | 2.92 | 9.6 | 61.2 |
| Equation | |
|---|---|
| Accuracy | |
| Mean of S/O ratio | |
| Slope of the linear regression, β1 | |
| Mean bias, MB (%) | |
| Precision | |
| Coefficient of variation of S/O (%) | |
| Coefficient of determination, R2 | |
| Model efficiency, ME | |
| Combination of accuracy and precision | |
| Mean square prediction error, MSPE | |
| Bias, B (%) | |
| Slope, Sl (%) | |
| Random, Rd (%) | |
| Concordance correlation coefficient, CCC | |
| Accuracy component, Ca | |
| Precision component (Pearson correlation coefficient), R |
| Without Leucaena | |
|---|---|
| N | 22 1 |
| Accuracy | |
| Mean S/O ratio | 1.07 |
| Slope | 1.40 |
| Mean bias (%) | 2.23 |
| Precision | |
| Coefficient of variation of S/O ratio (%) | 17.0 |
| R2 | 0.886 |
| Model efficiency | 0.809 |
| Combined accuracy and precision | |
| Mean square prediction error | 0.426 |
| Bias (%) | 2.1 |
| Slope (%) | 38.0 |
| Random (%) | 59.8 |
| Concordance correlation coefficient | 0.869 |
| Ca | 0.923 |
| R | 0.941 |
| All Diets | Diets without Leucaena | Diets with Leucaena | |
|---|---|---|---|
| N | 22 | 12 | 10 1 |
| Accuracy | |||
| Mean S/O ratio | 0.697 | 0.637 | 0.769 |
| Slope | 0.907 | 2.559 | 0.153 |
| Mean bias (%) | −30.5 | −39.2 | −23.5 |
| Precision | |||
| Coefficient of variation of S/O ratio (%) | 21.1 | 15.6 | 21.7 |
| R2 | 0.609 | 0.922 | 0.055 |
| Model efficiency | −0.431 | −0.659 | −7.594 |
| Combined accuracy and precision | |||
| Mean square prediction error | 3259 | 3500 | 2969 |
| Bias (%) | 72.2 | 74.6 | 69.3 |
| Slope (%) | 0.4 | 20.7 | 19.7 |
| Random (%) | 27.4 | 4.7 | 11.0 |
| Concordance correlation coefficient | 0.485 | 0.303 | 0.078 |
| Ca | 0.621 | 0.316 | 0.330 |
| R | 0.780 | 0.960 | 0.235 |
| Variable | Effect on Methane Emissions |
|---|---|
| NDF | - |
| Non-structural carbohydrates | - |
| Fat | - |
| Ashes | - |
| Crude protein | + |
| IVDMD | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruden, A.; Rivera, B.; Vargas, J.E.; López, S.; Gaviria, X.; Chirinda, N.; Arango, J. Evaluation of a Model (RUMINANT) for Prediction of DMI and CH4 from Tropical Beef Cattle. Animals 2023, 13, 721. https://doi.org/10.3390/ani13040721
Ruden A, Rivera B, Vargas JE, López S, Gaviria X, Chirinda N, Arango J. Evaluation of a Model (RUMINANT) for Prediction of DMI and CH4 from Tropical Beef Cattle. Animals. 2023; 13(4):721. https://doi.org/10.3390/ani13040721
Chicago/Turabian StyleRuden, Alejandro, Bernardo Rivera, Julio Ernesto Vargas, Secundino López, Xiomara Gaviria, Ngonidzashe Chirinda, and Jacobo Arango. 2023. "Evaluation of a Model (RUMINANT) for Prediction of DMI and CH4 from Tropical Beef Cattle" Animals 13, no. 4: 721. https://doi.org/10.3390/ani13040721