

Putative Role of CFSH in the Eyestalk-AG-Testicular Endocrine Axis of the Swimming Crab Portunus trituberculatus

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Extraction of Total RNA and cDNA Synthesis
2.3. Gene Cloning and Bioinformatics Analyses
2.4. Semiquantitative PCR
2.5. RNA Interference and Recombinant Protein Preparation and Injection
2.6. Quantitative Real-Time PCR
2.7. Western Blot Analysis
2.8. Measurement of the Level of cAMP, cGMP, and NOS in Testis
2.9. Histological Observation of the Testis of RNAi and Recombinant Protein Injection
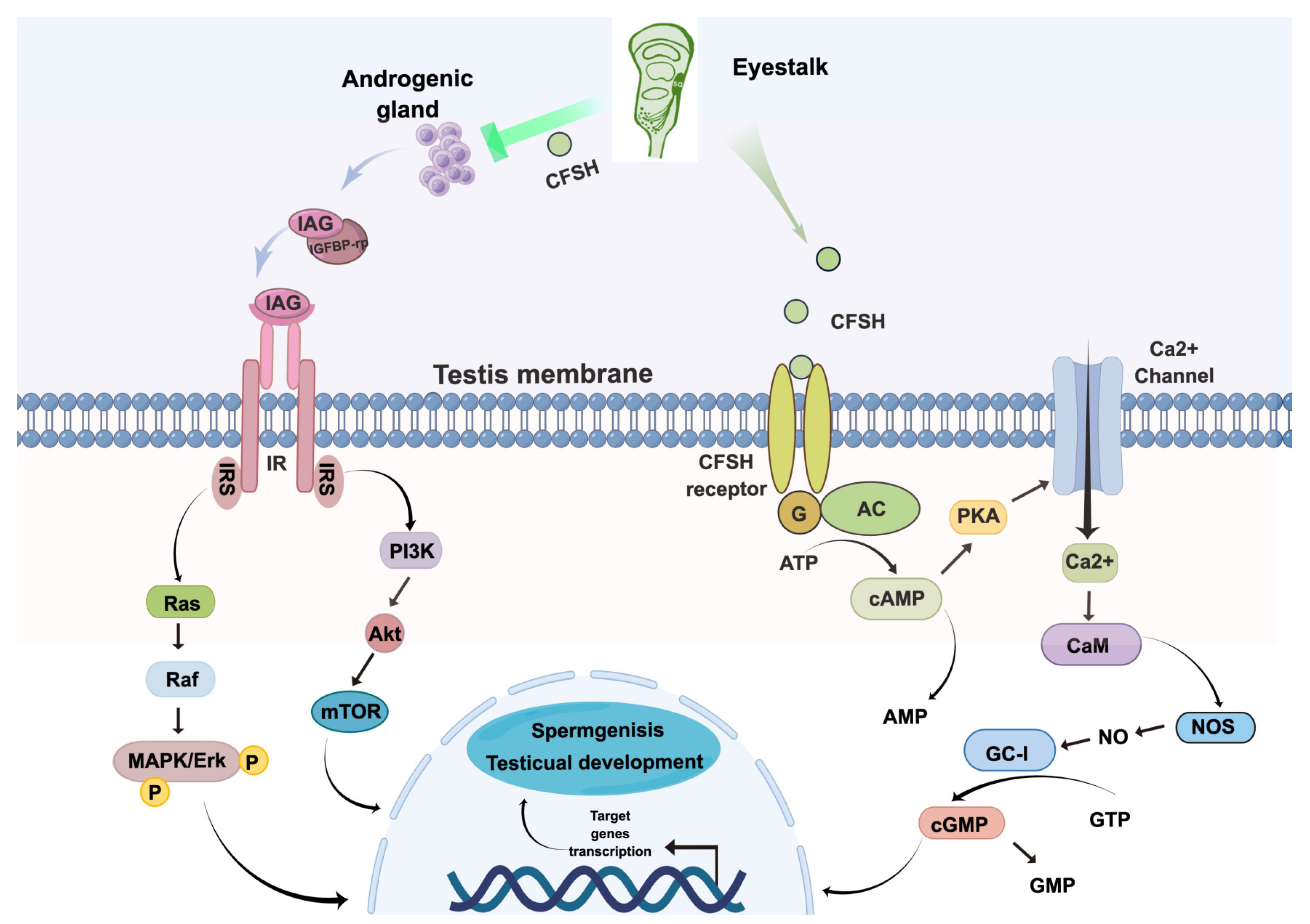
2.10. Conceptual Diagrams of Hypothetical Pathways
2.11. Statistical Analysis
3. Results
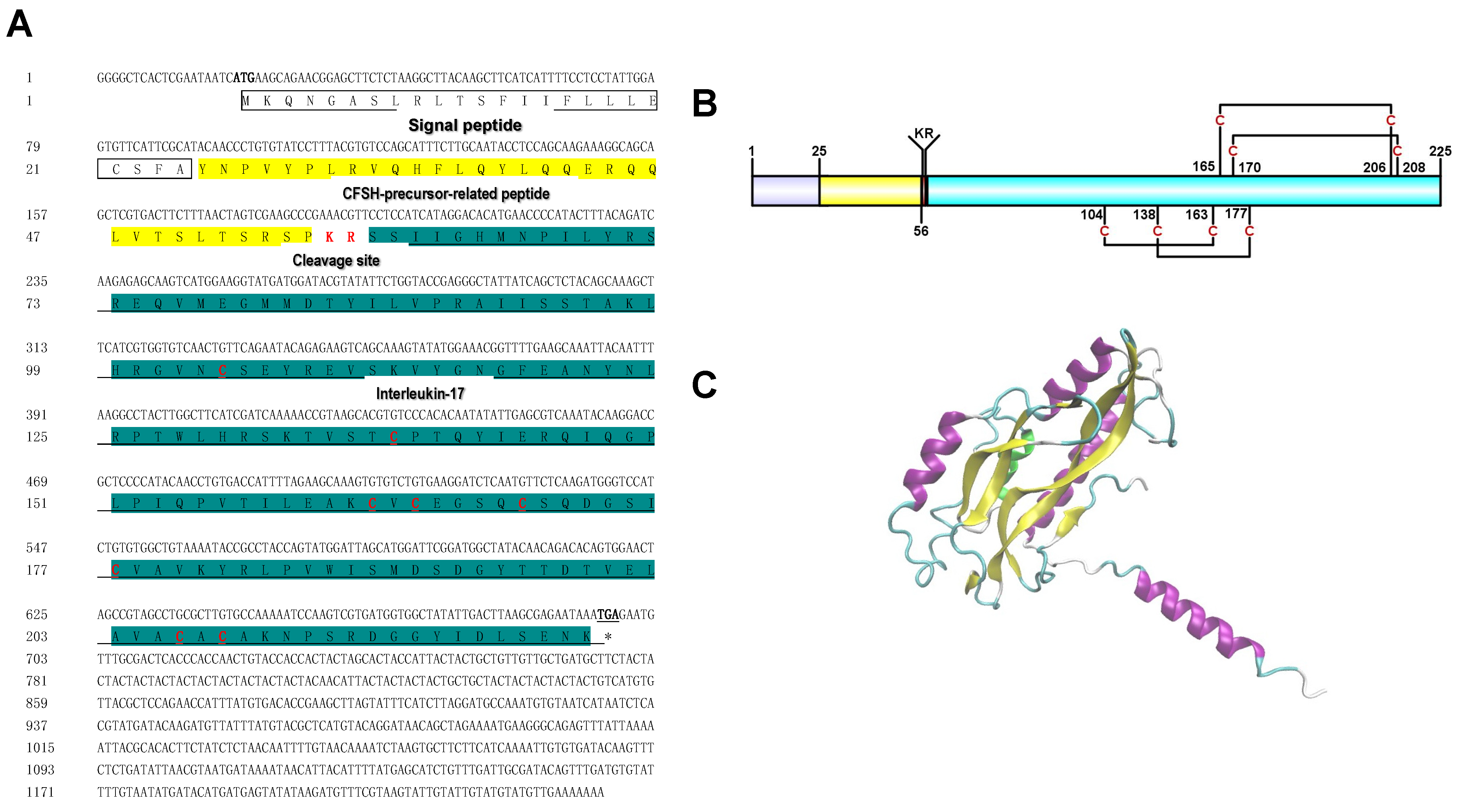
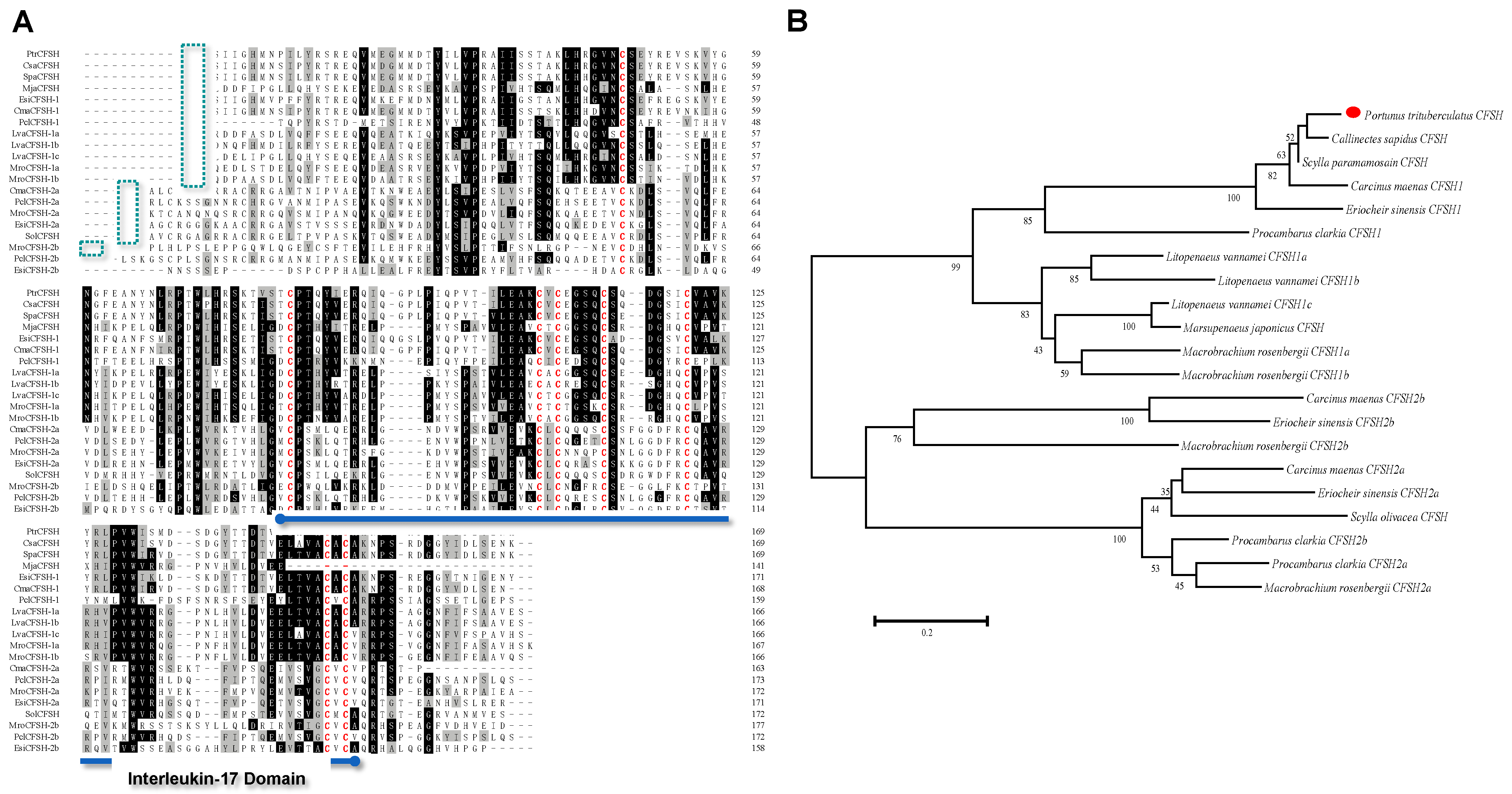
3.1. Molecular Characterization and Tissue Distribution of PtCFSH
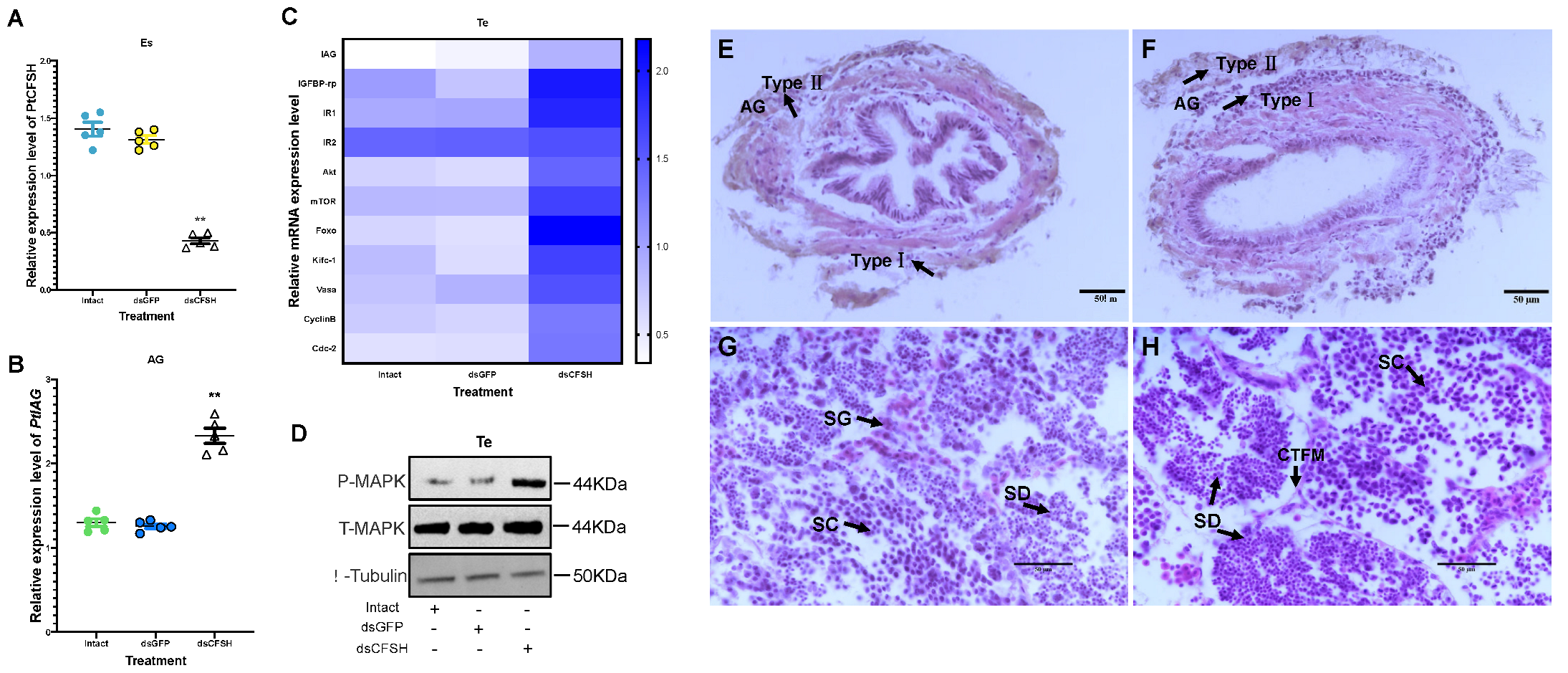
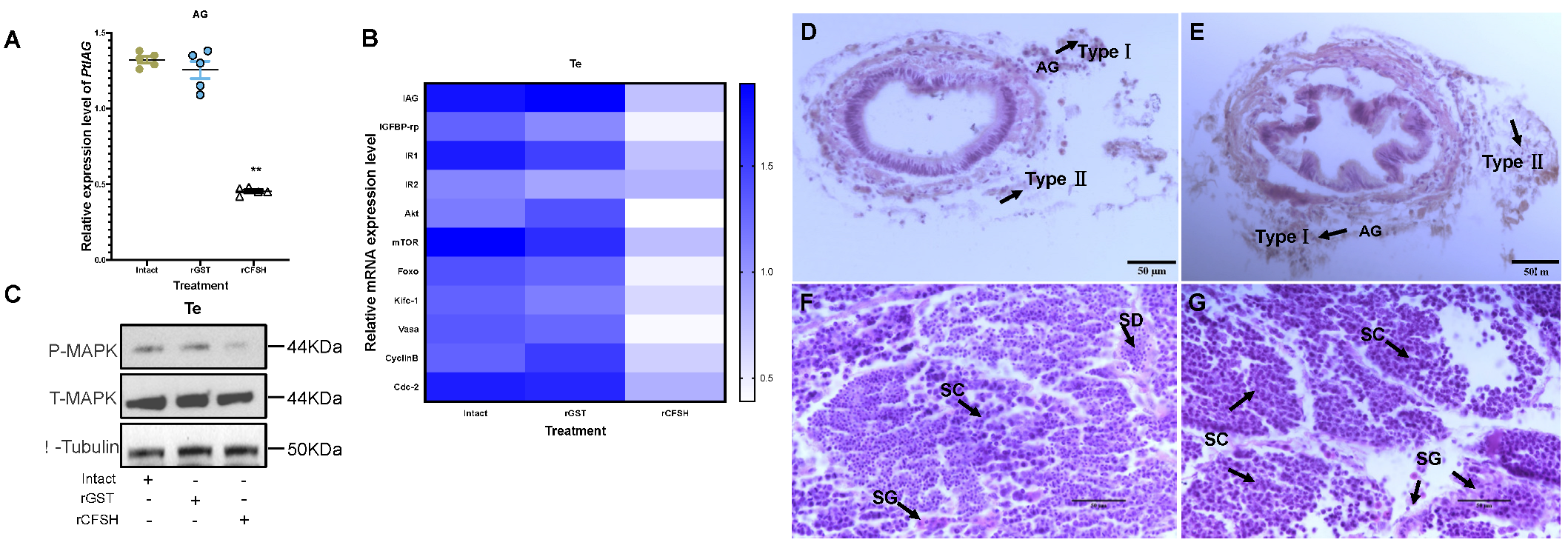
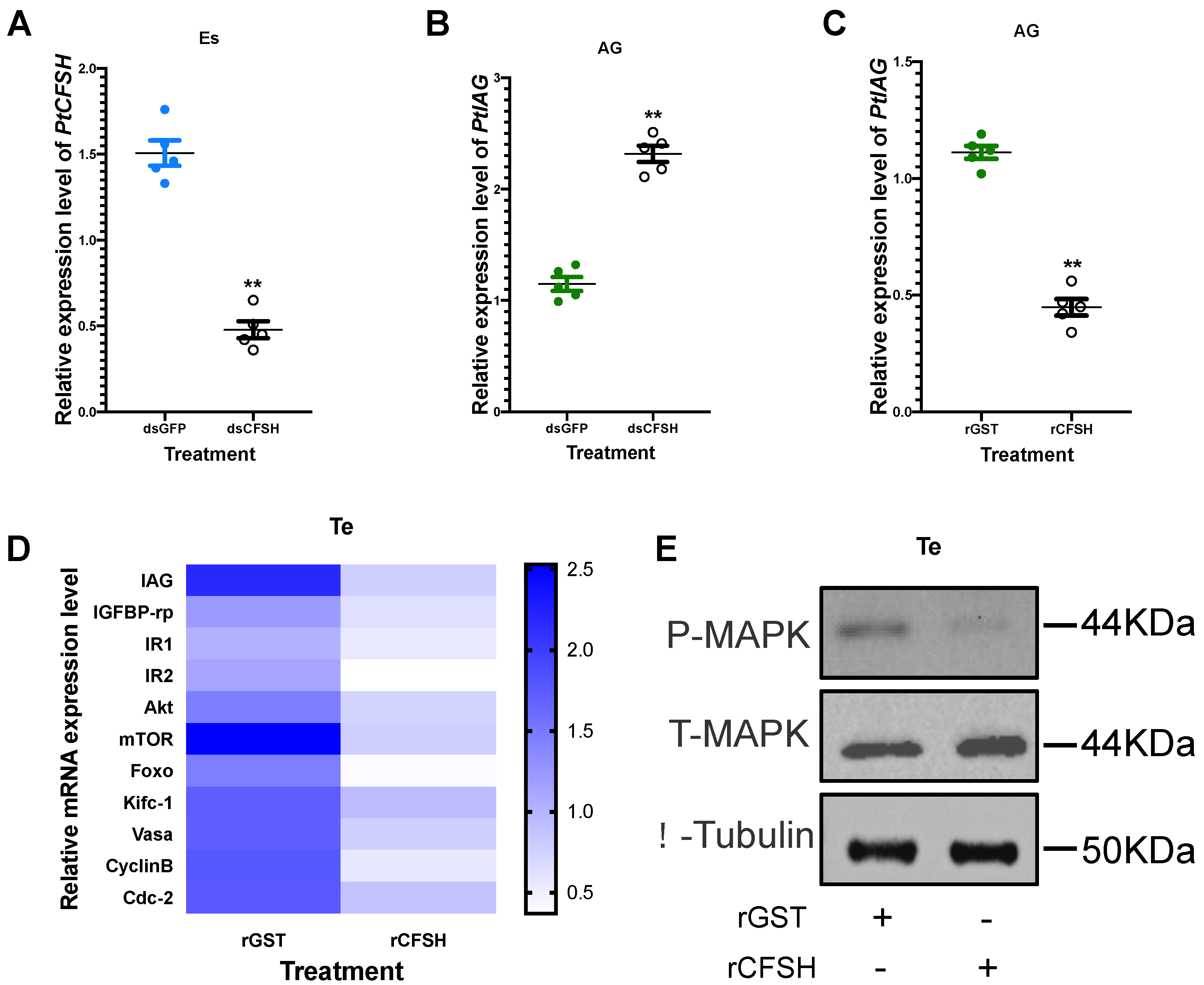
3.2. Effects of PtCFSH on AG
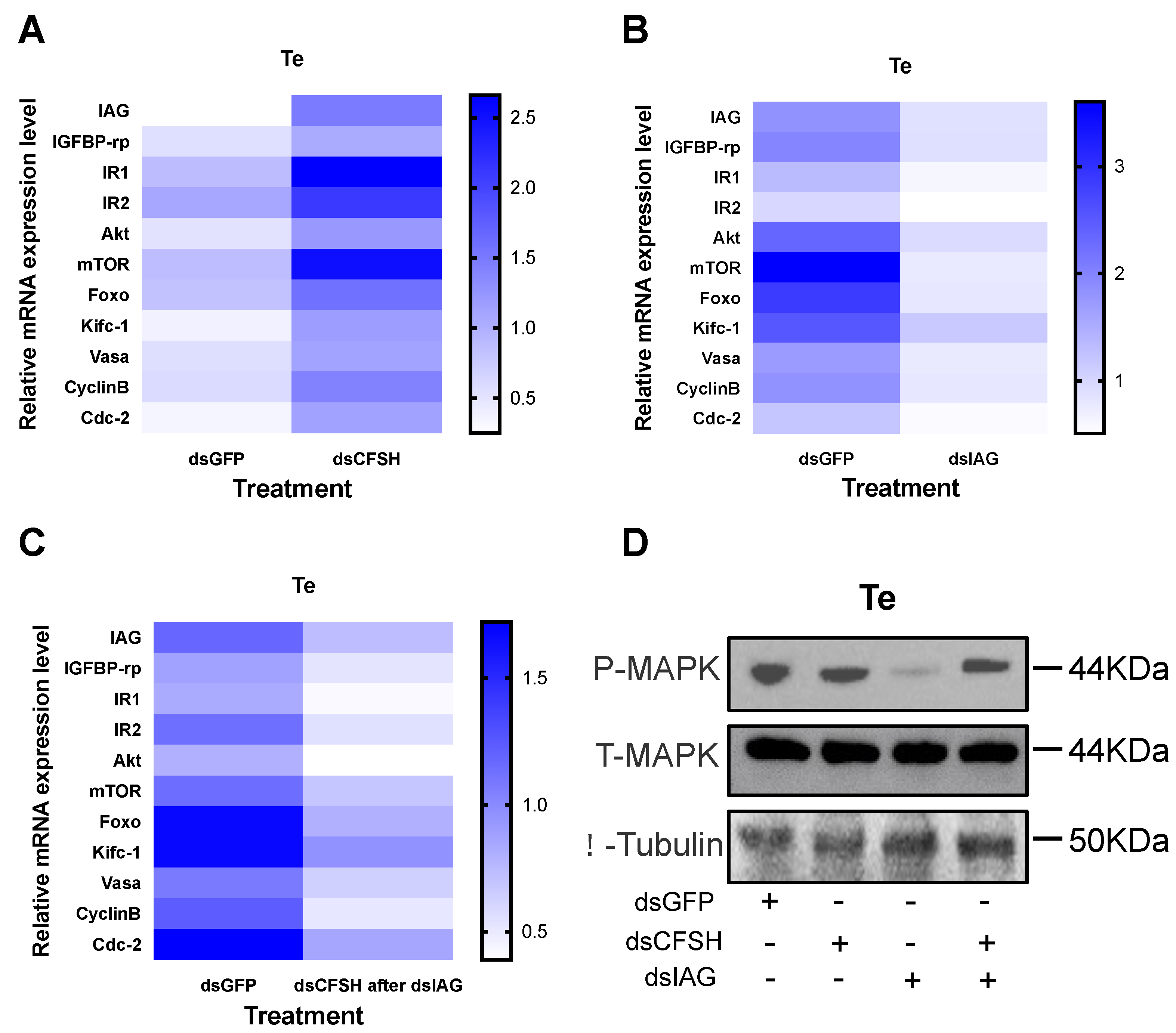
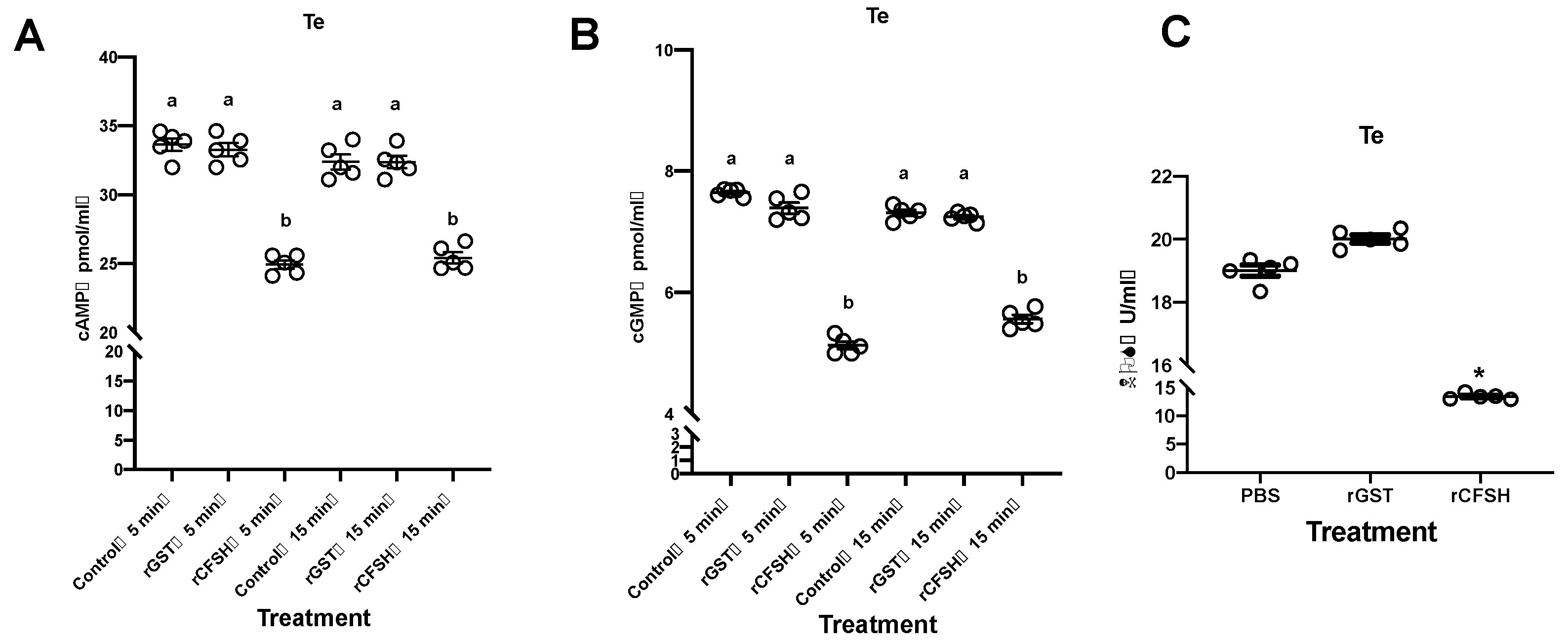
3.3. Effects of PtCFSH on Testis
3.4. Validation of the CFSH–IAG–Testis Endocrine Axis
3.5. Possible Signaling Mechanisms of PtCFSH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malecha, S.R.; Nevin, P.A.; Ha, P.; Barck, L.E.; Lamadrid-Rose, Y.; Masuno, S.; Hedgecock, D. Sex-ratios and sex-determination in progeny from crosses of surgically sex-reversed freshwater prawns, Macrobrachium rosenbergii. Aquaculture 1992, 105, 201–218. [Google Scholar] [CrossRef]
- Taketomi, Y.; Nishikawa, S. Implantation of androgenic glands into immature female crayfish, Procambarus clarkii, with masculinization of sexual characteristics. J. Crustacean Biol. 1996, 16, 232–239. [Google Scholar] [CrossRef]
- Barki, A.; Karplus, I.; Manor, R.; Sagi, A. Intersexuality and behavior in crayfish: The de-masculinization effects of androgenic gland ablation. Horm. Behav. 2006, 50, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Aflalo, E.D.; Hoang, T.T.T.; Nguyen, V.; Lam, Q.; Nguyen, D.; Trinh, Q.; Raviv, S.; Sagi, A. A novel two-step procedure for mass production of all-male populations of the giant freshwater prawn Macrobrachium rosenbergii. Aquaculture 2006, 256, 468–478. [Google Scholar] [CrossRef]
- Fu, C.; Li, F.; Wang, L.; Wu, F.; Wang, J.; Fan, X.; Liu, T. Molecular characteristics and abundance of insulin-like androgenic gland hormone and effects of RNA interference in Eriocheir sinensis. Anim. Reprod. Sci. 2020, 215, 106332. [Google Scholar] [CrossRef]
- Rosen, O.; Manor, R.; Weil, S.; Gafni, O.; Linial, A.; Aflalo, E.D.; Ventura, T.; Sagi, A. A sexual shift induced by silencing of a single insulin-like gene in crayfish: Ovarian upregulation and testicular degeneration. PLoS ONE 2010, 5, e15281. [Google Scholar] [CrossRef] [Green Version]
- Levy, T.; Sagi, A. The “IAG-switch”—A key controlling element in decapod crustacean sex differentiation. Front. Endocrinol. 2020, 11, 651. [Google Scholar] [CrossRef]
- Khalaila, I.; Manor, R.; Weil, S.; Granot, Y.; Keller, R.; Sagi, A. The eyestalk–androgenic gland–testis endocrine axis in the crayfish Cherax quadricarinatus. Gen. Comp. Endocrinol. 2002, 127, 147–156. [Google Scholar] [CrossRef]
- Charniaux-Cotton, H. Crustacean reproduction. Endocrinol. Sel. Invertebr. Types 1988, 2, 279–303. [Google Scholar]
- Payen, G. Etude descriptive des principales étapes de la morphogenèse sexuelle chez un crustace décapode à développement condense, l’Ecrevisse Pontastacus leptodactylus leptodactylus (Eschscholtz, 1823). Ann. Embryol. Morphog. 1973, 6, 179–206. [Google Scholar]
- Taketomi, Y.; Nishikawa, S.; Koga, S. Testis and androgenic gland during development of external sexual characteristics of the crayfish Procambarus clarkii. J. Crustacean Biol. 1996, 16, 24–34. [Google Scholar] [CrossRef]
- Vázquez-Islas, G.; Guerrero-Tortolero, D.A.; Garza-Torres, R.; Álvarez-Ruiz, P.; Mejía-Ruiz, H.; Campos-Ramos, R. Quantitative analysis of hypertrophy and hyperactivity in the androgenic gland of eyestalk-ablated male Pacific white shrimp Litopenaeus vannamei during molt stages. Aquaculture 2015, 439, 7–13. [Google Scholar] [CrossRef]
- Rao, K.R. Crustacean pigmentary-effector hormones: Chemistry and functions of RPCH, PDH, and related peptides. Am. Zool. 2001, 41, 364–379. [Google Scholar] [CrossRef] [Green Version]
- Webster, S.G.; Keller, R.; Dircksen, H. The CHH-superfamily of multifunctional peptide hormones controlling crustacean metabolism, osmoregulation, moulting, and reproduction. Gen. Comp. Endocrinol. 2012, 175, 217–233. [Google Scholar] [CrossRef]
- Li, F.; Bai, H.; Zhang, W.; Fu, H.; Jiang, F.; Liang, G.; Jin, S.; Sun, S.; Qiao, H. Cloning of genomic sequences of three crustacean hyperglycemic hormone superfamily genes and elucidation of their roles of regulating insulin-like androgenic gland hormone gene. Gene 2015, 561, 68–75. [Google Scholar] [CrossRef]
- Zmora, N.; Chung, J.S. A novel hormone is required for the development of reproductive phenotypes in adult female crabs. Endocrinology 2014, 155, 230–239. [Google Scholar] [CrossRef]
- Ventura, T.; Cummins, S.F.; Fitzgibbon, Q.; Battaglene, S.; Elizur, A. Analysis of the central nervous system transcriptome of the eastern rock lobster Sagmariasus verreauxi reveals its putative neuropeptidome. PLoS ONE 2014, 9, e97323. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Cummins, S.F.; Elizur, A.; Ventura, T. Transcriptomic characterization and curation of candidate neuropeptides regulating reproduction in the eyestalk ganglia of the Australian crayfish, Cherax quadricarinatus. Sci. Rep. 2016, 6, 38658. [Google Scholar] [CrossRef] [Green Version]
- Tsutsui, N.; Kotaka, S.; Ohira, T.; Sakamoto, T. Characterization of distinct ovarian isoform of crustacean female sex hormone in the kuruma prawn Marsupenaeus japonicus. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2018, 217, 7–16. [Google Scholar] [CrossRef]
- Liu, A.; Liu, J.; Liu, F.; Huang, Y.; Wang, G.; Ye, H. Crustacean female sex hormone from the mud crab Scylla paramamosain is highly expressed in prepubertal males and inhibits the development of androgenic gland. Front. Physiol. 2018, 9, 924. [Google Scholar] [CrossRef] [Green Version]
- Thongbuakaew, T.; Suwansa-ard, S.; Sretarugsa, P.; Sobhon, P.; Cummins, S.F. Identification and characterization of a crustacean female sex hormone in the giant freshwater prawn, Macrobrachium rosenbergii. Aquaculture 2019, 507, 56–68. [Google Scholar] [CrossRef]
- Liu, F.; Shi, W.; Huang, L.; Wang, G.; Zhu, Z.; Ye, H. Roles of Crustacean Female Sex Hormone 1a in a Protandric Simultaneous Hermaphrodite Shrimp. Front. Mar. Sci. 2021, 8, 95–153. [Google Scholar] [CrossRef]
- Zhu, D.; Feng, T.; Mo, N.; Han, R.; Lu, W.; Shao, S.; Cui, Z. New insights for the regulatory feedback loop between type 1 crustacean female sex hormone (CFSH-1) and insulin-like androgenic gland hormone (IAG) in the Chinese mitten crab (Eriocheir sinensis). Front. Physiol. 2022, 13, 1054773. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Liu, Z.; Liu, M.; Tao, T.; Shen, X.; Zhu, D. Role of Halloween genes in ecdysteroids biosynthesis of the swimming crab (Portunus trituberculatus): Implications from RNA interference and eyestalk ablation. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 199, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Xie, X.; Tao, T.; Jiang, Q.; Shao, J.; Zhu, D. Molecular characterization of methoprene-tolerant gene (Met) in the swimming crab Portunus trituberculatus: Its putative role in methyl farnesoate-mediated vitellogenin transcriptional activation. Anim. Reprod. Sci. 2016, 174, 132–142. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chang, E.S.; Mykles, D.L. Regulation of crustacean molting: A review and our perspectives. Gen. Comp. Endocrinol. 2011, 172, 323–330. [Google Scholar] [CrossRef]
- Sroyraya, M.; Chotwiwatthanakun, C.; Stewart, M.J.; Soonklang, N.; Kornthong, N.; Phoungpetchara, I.; Hanna, P.J.; Sobhon, P. Bilateral eyestalk ablation of the blue swimmer crab, Portunus pelagicus, produces hypertrophy of the androgenic gland and an increase of cells producing insulin-like androgenic gland hormone. Tissue Cell 2010, 42, 293–300. [Google Scholar] [CrossRef]
- Zhang, E.; Qiu, G. A novel Dmrt gene is specifically expressed in the testis of Chinese mitten crab, Eriocheir sinensis. Dev. Genes Evol. 2010, 220, 151–159. [Google Scholar] [CrossRef]
- Huang, X.; Bae, S.H.; Bachvaroff, T.R.; Schott, E.J.; Ye, H.; Chung, J.S. Does a blue crab putative insulin-like peptide binding protein (ILPBP) play a role in a virus infection? Fish Shellfish. Immunol. 2016, 58, 340–348. [Google Scholar] [CrossRef]
- Senarai, T.; Saetan, J.; Tamtin, M.; Weerachatyanukul, W.; Sobhon, P.; Sretarugsa, P. Presence of gonadotropin-releasing hormone-like peptide in the central nervous system and reproductive organs of the male blue swimming crab, Portunus pelagicus, and its effect on spermatogenesis. Cell Tissue Res. 2016, 365, 265–277. [Google Scholar] [CrossRef]
- Aizen, J.; Chandler, J.C.; Fitzgibbon, Q.P.; Sagi, A.; Battaglene, S.C.; Elizur, A.; Ventura, T. Production of recombinant insulin-like androgenic gland hormones from three decapod species: In vitro testicular phosphorylation and activation of a newly identified tyrosine kinase receptor from the Eastern spiny lobster, Sagmariasus verreauxi. Gen. Comp. Endocrinol. 2016, 229, 8–18. [Google Scholar] [CrossRef]
- Jiang, Q.; Lu, B.; Lin, D.; Huang, H.; Chen, X.; Ye, H. Role of crustacean female sex hormone (CFSH) in sex differentiation in early juvenile mud crabs, Scylla paramamosain. Gen. Comp. Endocrinol. 2020, 289, 113383. [Google Scholar] [CrossRef]
- Kotaka, S.; Ohira, T. cDNA cloning and in situ localization of a crustacean female sex hormone-like molecule in the kuruma prawn Marsupenaeus japonicus. Fish. Sci. 2018, 84, 53–60. [Google Scholar] [CrossRef]
- Veenstra, J.A. The power of next-generation sequencing as illustrated by the neuropeptidome of the crayfish Procambarus clarkii. Gen. Comp. Endocrinol. 2015, 224, 84–95. [Google Scholar] [CrossRef]
- Bulaj, G. Formation of disulfide bonds in proteins and peptides. Biotechnol. Adv. 2005, 23, 87–92. [Google Scholar] [CrossRef]
- Wojtasek, H.; Prestwich, G.D. Key disulfide bonds in an insect hormone binding protein: cDNA cloning of a juvenile hormone binding protein of Heliothis virescens and ligand binding by native and mutant forms. Biochemistry 1995, 34, 5234–5241. [Google Scholar] [CrossRef]
- Ma, D.-D.; Pan, M.-Y.; Hou, C.-C.; Tan, F.-Q.; Yang, W.-X. KIFC1 and myosin Va: Two motors for acrosomal biogenesis and nuclear shaping during spermiogenesis of Portunus trituberculatus. Cell Tissue Res. 2017, 369, 625–640. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Han, K.; Zou, Z.; Zhang, Z. A vasa gene from green mud crab Scylla paramamosain and its expression during gonadal development and gametogenesis. Mol. Biol. Rep. 2012, 39, 4327–4335. [Google Scholar] [CrossRef]
- Guo, Q.; Li, S.; Lv, X.; Xiang, J.; Sagi, A.; Manor, R.; Li, F. A putative insulin-like androgenic gland hormone receptor gene specifically expressed in male Chinese shrimp. Endocrinology 2018, 159, 2173–2185. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.-E.; Zheng, H.; Xie, X.; Xu, R.; Zhu, D. Molecular identification and putative role of insulin growth factor binding protein-related protein (IGFBP-rp) in the swimming crab Portunus trituberculatus. Gene 2022, 833, 146551. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.; Li, C.; Wu, W.; Li, B. Identification and evolution of two insulin receptor genes involved in Tribolium castaneum development and reproduction. Gene 2016, 585, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zheng, H.; Zheng, L.; Wang, Y.; Wang, M.; Xie, X.; Zhu, D. Molecular characterization of the insulin-like androgenic gland hormone in the swimming crab, Portunus trituberculatus, and its involvement in the insulin signaling system. Front. Endocrinol. 2020, 11, 585. [Google Scholar] [CrossRef] [PubMed]
- Bao, C.; Yang, Y.; Huang, H.; Ye, H. Inhibitory role of the mud crab short neuropeptide F in vitellogenesis and oocyte maturation via autocrine/paracrine signaling. Front. Endocrinol. 2018, 9, 390. [Google Scholar] [CrossRef] [Green Version]
- Christie, A.E.; Stemmler, E.A.; Dickinson, P.S. Crustacean neuropeptides. Cell. Mol. Life Sci. 2010, 67, 4135–4169. [Google Scholar] [CrossRef]
- Jayasankar, V.; Tomy, S.; Wilder, M.N. Insights on molecular mechanisms of ovarian development in decapod crustacea: Focus on vitellogenesis-stimulating factors and pathways. Front. Endocrinol. 2020, 11, 577925. [Google Scholar] [CrossRef]
- Shimizu, H.; Morio, T.; Shimizu, H.D.; Urushihara, H. A mutation in the cAMP signaling pathway affects sexual development of Dictyostelium discoideum. Dev. Growth Differ. 1997, 39, 227–234. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | PCR Objective |
|---|---|---|
| PtCFSH-F1 | CGTCAAATACAAGGACCGCTC | 3′ RACE |
| PtCFSH-F2 | CAATGTTCTCAAGATGGGTCC | 3′ RACE |
| PtCFSH-R1 | CCTCGCAGTCCAGAACCAA | 5′ RACE |
| PtCFSH-R2 | GTGATAGAGGGCGTGGG | 5′ RACE |
| PtCFSH-F | TCCTATTGGAGTGTTCATTCGC | cDNA clone |
| PtCFSH-R | GTGGTGGTACAGTTGGTGGG | cDNA clone |
| PtCFSH-RT-F | ACCGCCTACCAGTATGGATTAG | RT-PCR |
| PtCFSH-RT-R | GCATCAGCAACAACAGCAGTA | RT-PCR |
| 5′outer | CTAATACGACTCACTATAGGGC | 5′ RACE |
| 5′inner | AAGCAGTGGTATCAACGCAGAGT | 5′ RACE |
| 3′inner | TCCACTAGTGATTTCACTATAGG | 3′ RACE |
| 3′outer | CTAATACGACTCACTATAGGGC | 3′ RACE |
| AP | TACCGTCGTTCCACTAGTGATTTCACTATAGG(T)17 | 3′ RACE |
| dsGFP-F | TAATACGACTCACTATAGGGCGACGTAAACGGCCACAAGT | RNAi |
| dsGFP-R | TAATACGACTCACTATAGGGCTTGTACAGCTCGTCCATGC | RNAi |
| dsPtCFSH-F | TAATACGACTCACTATAGGGAGATCCTATTGGAGTGTTCATTCG | RNAi |
| dsPtCFSH-R | TAATACGACTCACTATAGGGAGATACAGTTGGTGGGTGAGTCG | RNAi |
| dsPtIAG-F | TAATACGACTCACTATAGGGAAACGAAGACCCAATGCTACC | RNAi |
| dsPtIAG-R | TAATACGACTCACTATAGGGTTACTGCCTATTTCGGGAAGC | RNAi |
| PtCFSH-QF | GTATTTCATCTTAGGATGCCAA | qRT-PCR |
| PtCFSH-QR | TAAACTCTGCCCTTCATTTTCT | qRT-PCR |
| β-actin-QF | CGAAACCTTCAACACTCCCG | qRT-PCR |
| β-actin-QR | GATAGCGTGAGGAAGGGCATA | qRT-PCR |
| PtIGFBP-rp-QF | TTACCACTATTGACGGCACCT | qRT-PCR |
| PtIGFBP-rp-QR | TCATTATC TGTACCCATCCTGTT | qRT-PCR |
| PtIAG-QF | TCTTATTAGCGACTTCTCCG | qRT-PCR |
| PtIAG-QR | CCTCTGTCCCTCGTTTATGT | qRT-PCR |
| PtIR1-QF | CTGATGCGTTTGTCGTATTT | qRT-PCR |
| PtIR1-QR | GAAGCGTGGTGCCTATTT | qRT-PCR |
| PtIR2-QF | ACCAGCTAGTGGGAACCG | qRT-PCR |
| PtIR2-QR | GGGAGGGACTCTTTGACG | qRT-PCR |
| PtAkt-QF | CTCAACCAGGAACGCTTCTTC | qRT-PCR |
| PtAkt-QR | TGTGTCCATCAGCATCCAGTAA | qRT-PCR |
| PtmTOR-QF | TCTCCTGGCTGTTGCTGTC | qRT-PCR |
| PtmTOR-QR | GCTTCTTGCTTGGTGTATCCTT | qRT-PCR |
| PtAkt-QF | CTCAACCAGGAACGCTTCTTC | qRT-PCR |
| PtAkt-QR | TGTGTCCATCAGCATCCAGTAA | qRT-PCR |
| Ptcdc2-QF | CCGTCAAGCAGATGGACAGTG | qRT-PCR |
| Ptcdc2-QR | CCAGGTCGTCAAAGTAAGGGTG | qRT-PCR |
| PtCyclinB-QF | ATGTGCCACTACAAGGCGTCT | qRT-PCR |
| PtCyclinB-QR | ATCAGCGTGTCATTCCAATCC | qRT-PCR |
| PtFoxo-QF | CGGAGGTGAAGCACATCAAC | qRT-PCR |
| PtFoxo-QR | TCATTGGTGGAGGCAGAGTG | qRT-PCR |
| PtKifc1-QF | TCCAATCGCCATCTACCTCAG | qRT-PCR |
| PtKifc1-QR | CGTCTTCAGCATCTCCAGAATG | qRT-PCR |
| PtVasa-QF | GCTTGCCATCCAGATATTCCAT | qRT-PCR |
| PtVasa-QR | TGCTCCTTCATACGCCTCAA | qRT-PCR |
| PtCFSH-GST-SmaI-F | ggatccccaggaattcccgggATGAAGCAGAACGGAGCTTCTC | In-Fusion clone |
| PtCFSH-GST-XhoI-R | gtcacgatgcggccgctcgagTCATTTATTCTCGCTTAAGTCAATATAGC | In-Fusion clone |
| Species | Protein Name | GenBank Accession Number |
|---|---|---|
| Portunus trituberculatus | CFSH | ON929327 |
| Scylla paramamosain | CFSH | MN938502.1 |
| Callinectes sapidus | CFSH | GU016328.1 |
| Carcinus maenas | CFSH-1 | AEI72264.1 |
| Marsupenaeus japonicus | CFSH | BBA53799.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Xu, R.; Tu, S.; Yu, Q.; Xie, X.; Zhu, D. Putative Role of CFSH in the Eyestalk-AG-Testicular Endocrine Axis of the Swimming Crab Portunus trituberculatus. Animals 2023, 13, 690. https://doi.org/10.3390/ani13040690
Wang M, Xu R, Tu S, Yu Q, Xie X, Zhu D. Putative Role of CFSH in the Eyestalk-AG-Testicular Endocrine Axis of the Swimming Crab Portunus trituberculatus. Animals. 2023; 13(4):690. https://doi.org/10.3390/ani13040690
Chicago/Turabian StyleWang, Mengen, Rui Xu, Shisheng Tu, Qiaoling Yu, Xi Xie, and Dongfa Zhu. 2023. "Putative Role of CFSH in the Eyestalk-AG-Testicular Endocrine Axis of the Swimming Crab Portunus trituberculatus" Animals 13, no. 4: 690. https://doi.org/10.3390/ani13040690