
Expression and Characterization of the Spats1 Gene and Its Response to E2/MT Treatment in the Chinese Soft-Shelled Turtle (Pelodiscus sinensis)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethics Statement
2.2. Collection of Samples
2.2.1. Cloning and Tissue Distribution Expression
2.2.2. Seasonal Expression and In Situ Hybridization
2.2.3. Hormone Treatment in Embryonic Development
2.2.4. Hormone Treatment in Adults
2.3. RNA Extraction and cDNA Synthesis
2.4. Cloning and Sequencing
2.5. Structural Analysis
2.6. Cryostat Sections of the Gonad
2.7. In Situ Hybridization
2.8. Semi-Quantitative PCR and Real-Time Quantitative PCR
2.9. Statistical Analysis
3. Results
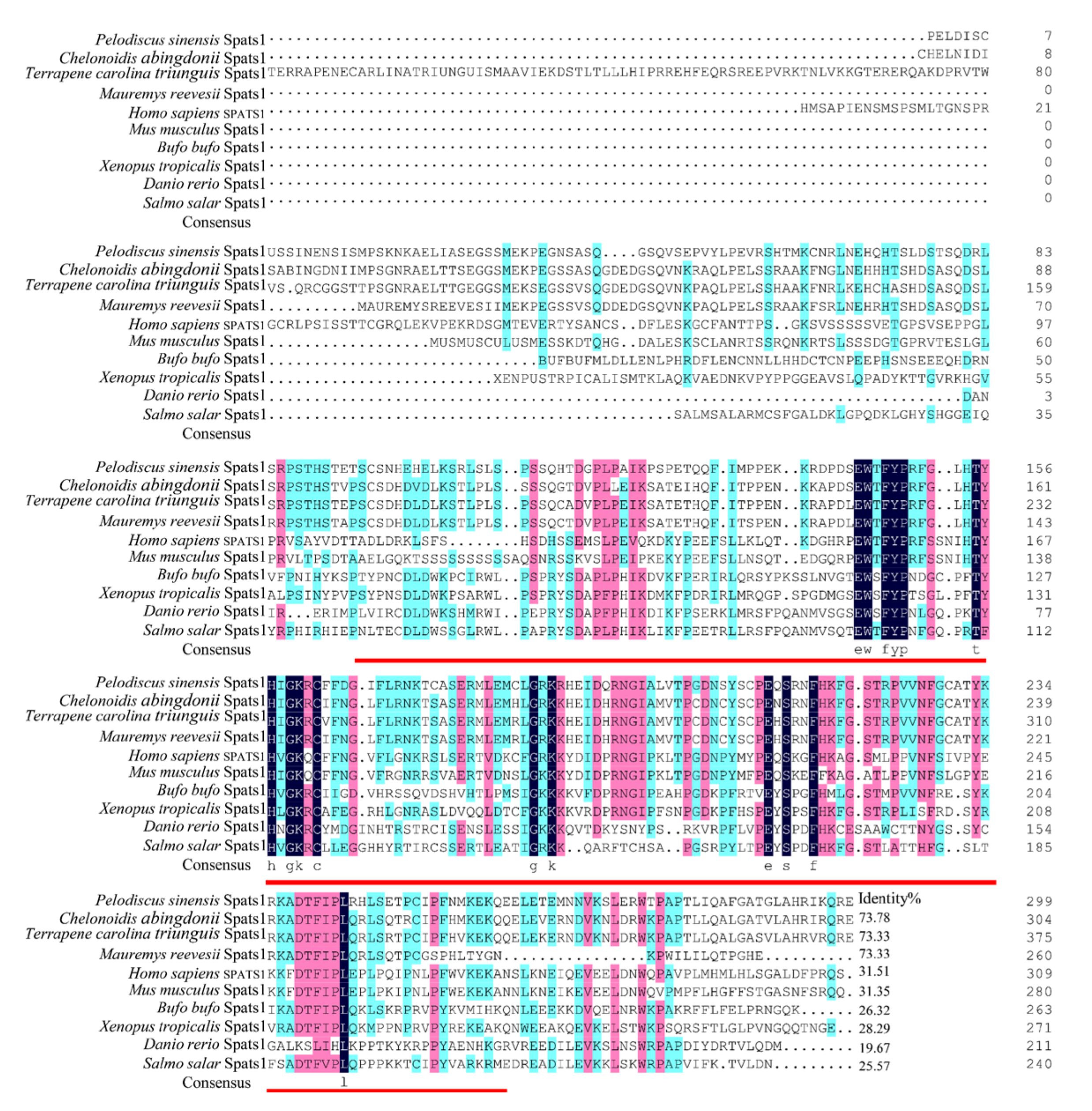
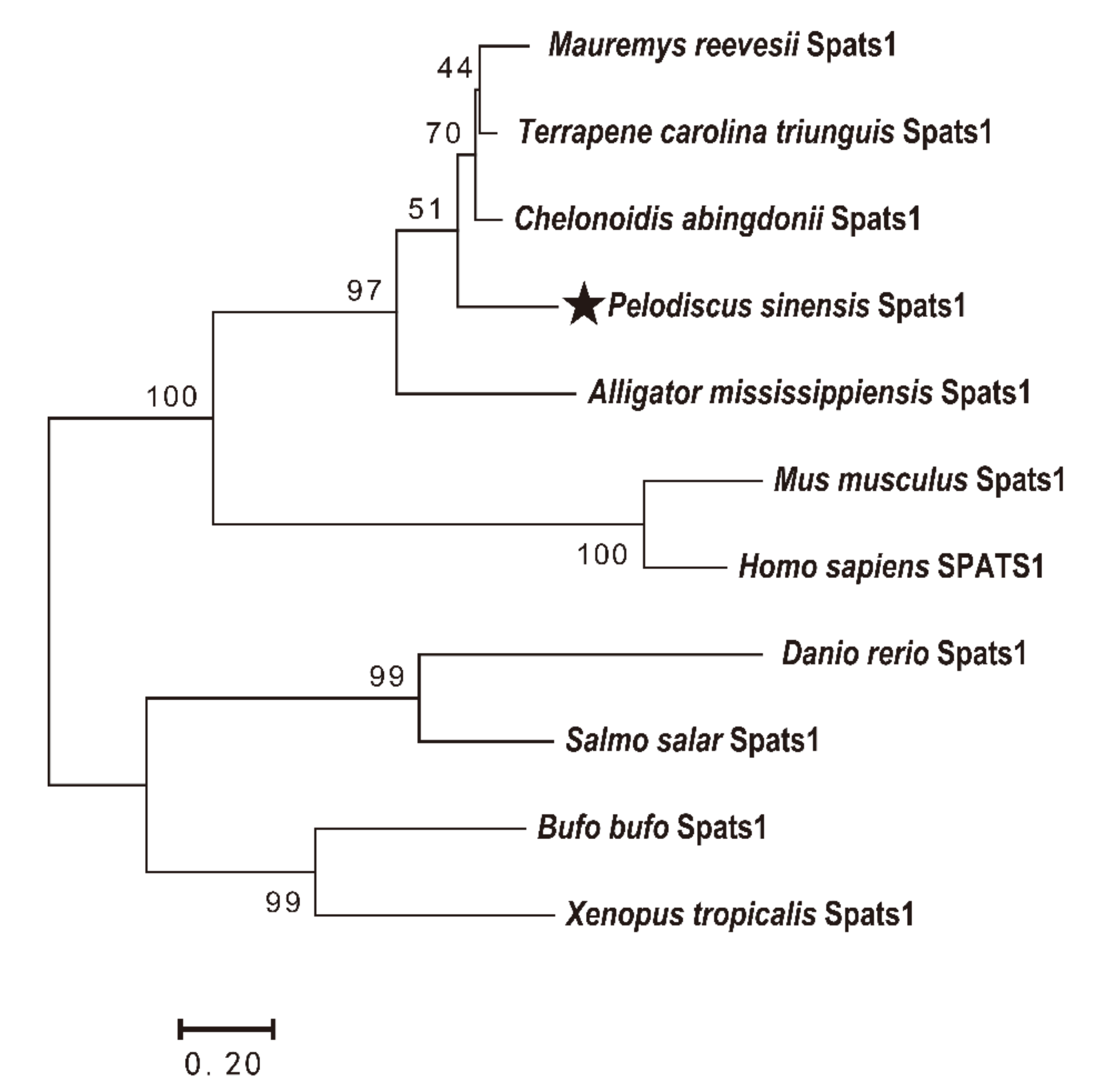
3.1. Cloning and Sequence Analyses
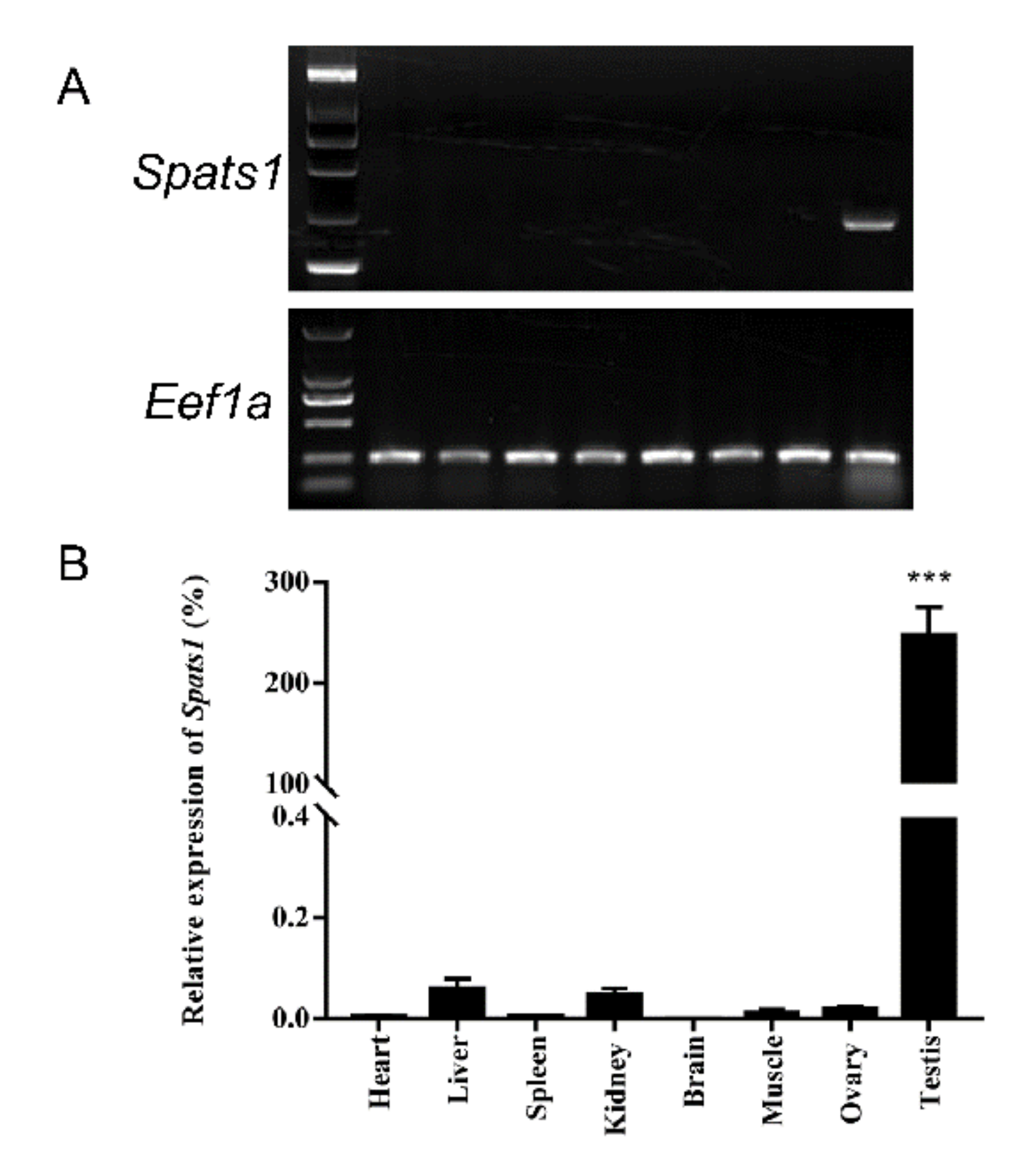
3.2. Tissue Distribution
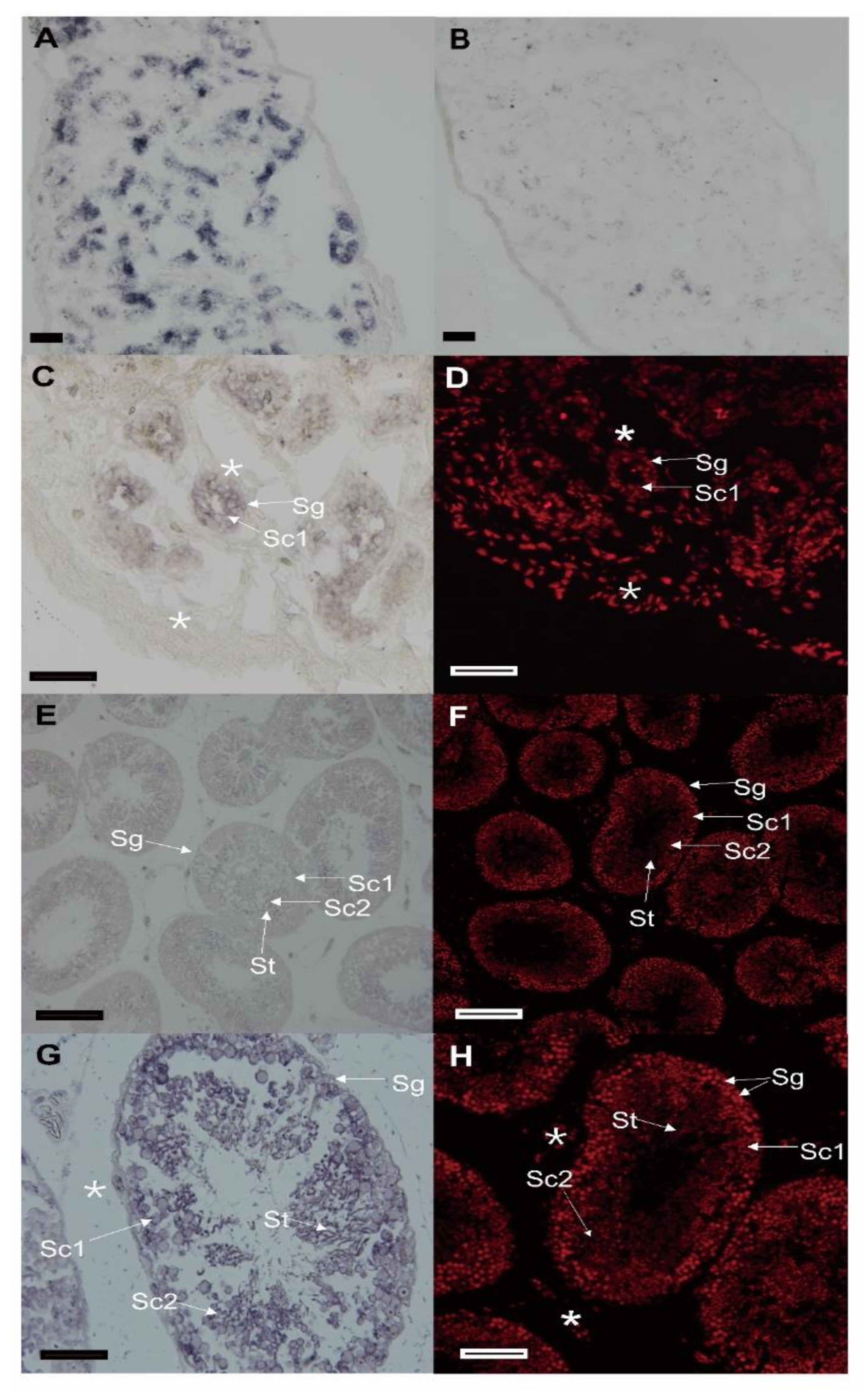
3.3. The Cellular Distribution of Spats1 mRNA in Testes
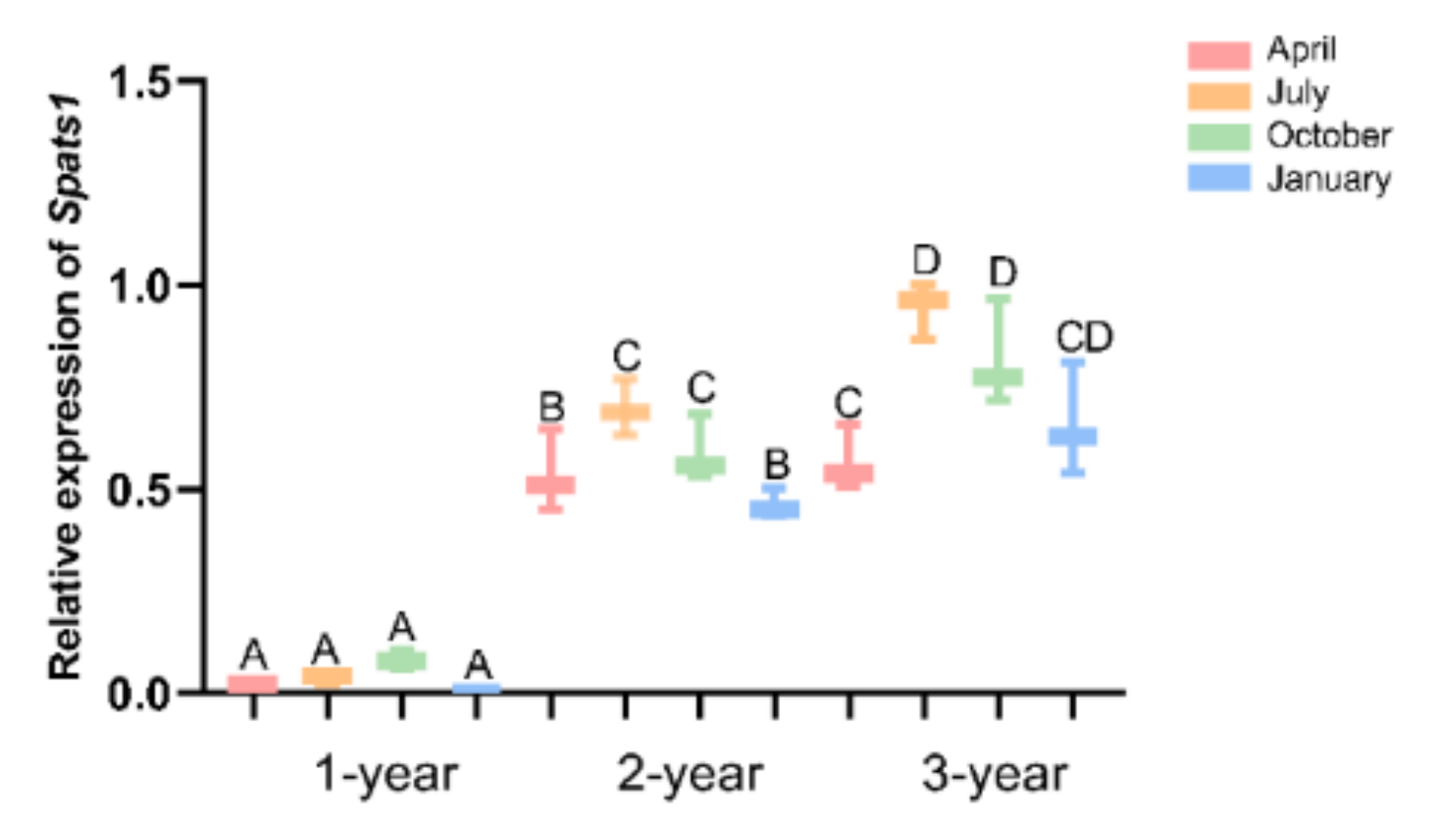
3.4. Expression Pattern of Spats1 mRNA in Testes of Different Seasons
3.5. Effect of E2 and MT Treatment on Embryos
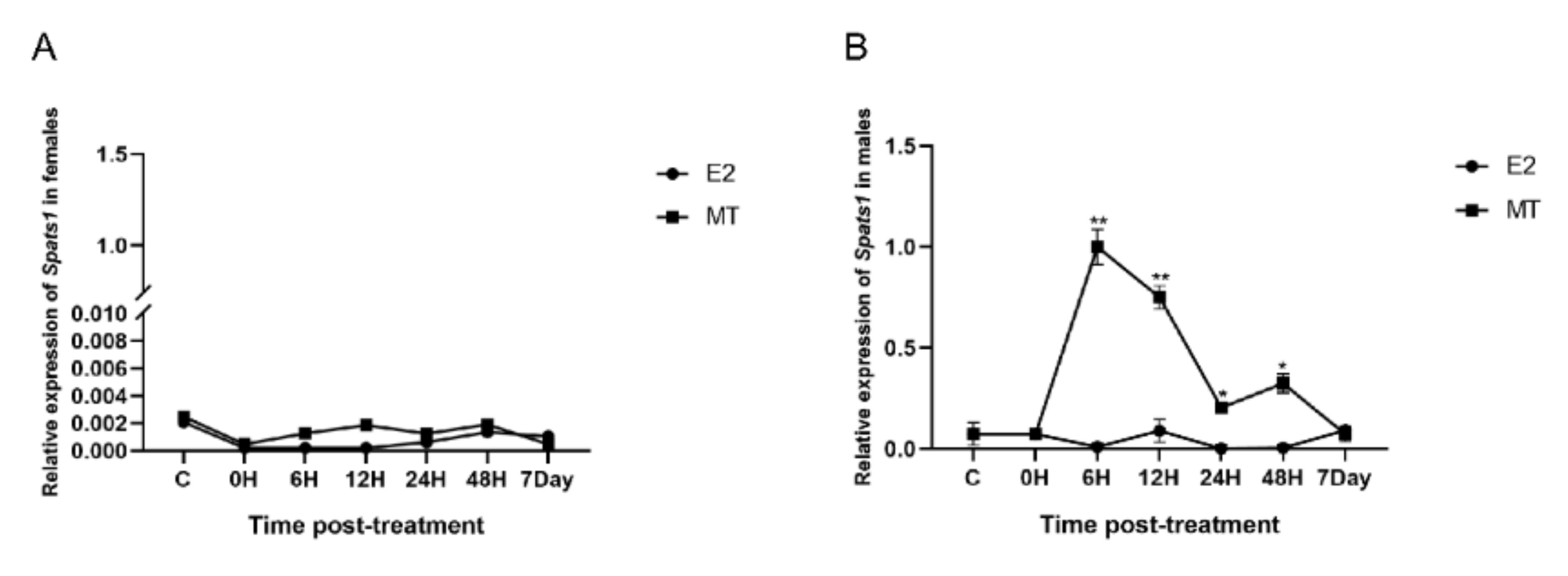
3.6. E2 and MT Treatment Were Performed on Adult Male and Female P. sinensis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, T.; Yang, W.-X. The dynamics and regulation of microfilament during spermatogenesis. Gene 2020, 744, 144635. [Google Scholar] [CrossRef] [PubMed]
- Gribbins, K. Reptilian spermatogenesis: A histological and ultrastructural perspective. Spermatogenesis 2011, 1, 250–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, L.; Agnese, M.; Lorenzo, M.; Barra, T.; Valiante, S.; Prisco, M. Spermatogenesis and regulatory factors in the wall lizard Podarcis sicula. Gen. Comp. Endocrinol. 2020, 298, 113579. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Li, W.; Zhao, J.; Liu, H.; Yu, L.; Zhu, X. Reproductive performance is associated with seasonal plasma reproductive hormone levels, steroidogenic enzymes and sex hormone receptor expression levels in cultured Asian yellow pond turtles (Mauremys mutica). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 254, 110566. [Google Scholar] [CrossRef] [PubMed]
- Resende, F.C.d.; Avelar, G.F.d. The sexual segment of the kidney of a tropical rattlesnake, Crotalus durissus (Reptilia, Squamata, Viperidae), and its relationship to seasonal testicular and androgen cycles. J. Morphol. 2021, 282, 1402–1414. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, X.; Wang, X.; Peng, N.; Luo, Z. Effects of dietary xanthophyll supplementation on growth performance, body color, carotenoids, and blood chemistry indices of chinese soft-shelled turtle Pelodiscus sinensis. N. Am. J. Aquac. 2020, 82, 394–404. [Google Scholar] [CrossRef]
- Gauthier, J.; Cannatella, D.C.; De Queiroz, K.; Kluge, A.G.; Rowe, T. Tetrapod phylogeny. Hierarchy Life 1989, 25, 337–353. [Google Scholar]
- Uno, Y.; Nishida, C.; Tarui, H.; Ishishita, S.; Takagi, C.; Nishimura, O.; Ishijima, J.; Ota, H.; Kosaka, A.; Matsubara, K.; et al. Inference of the protokaryotypes of amniotes and tetrapods and the evolutionary processes of microchromosomes from comparative gene mapping. PLoS ONE 2012, 7, e53027. [Google Scholar] [CrossRef]
- Chen, H.; Huang, Y.; Liu, T.; Haseeb, A.; Ahmed, N.; Zhang, L.; Bian, X.; Chen, Q. Characteristics of seasonal spermatogenesis in the soft-shelled turtle. Anim. Reprod. Sci. 2020, 214, 106307. [Google Scholar] [CrossRef]
- Zhang, L.; Han, X.; Li, M.; Bao, H.; Chen, Q. Spermiogenesis in soft-shelled turtle, Pelodiscus sinensis. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2007, 290, 1213–1222. [Google Scholar] [CrossRef]
- Bian, X.; Zhang, L.; Yang, L.; Yang, P.; Ullah, S.; Zhang, Q.; Chen, Q. Ultrastructure of epididymal epithelium and its interaction with the sperm in the soft-shelled turtle Pelodiscus sinensis. Micron 2013, 54, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Viets, B.; Ewert, M.; Talent, L.; Nelson, C. Sex-determining mechanisms in squamate reptiles. J. Exp. Zool. 1994, 270, 45–56. [Google Scholar] [CrossRef]
- İnanan, B.E.; Acar, Ü. Comparison of milt and blood parameters including testosterone and gonadotropins levels of sex-reversed female rainbow trout (Oncorhynchus mykiss) by different steroid hormones. Aquac. Res. 2021, 52, 1876–1884. [Google Scholar] [CrossRef]
- Ruiz-García, A.; Roco, Á.S.; Bullejos, M. Sex differentiation in amphibians: Effect of temperature and its influence on sex reversal. Sex. Dev. 2021, 15, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, T.; Mi, M.; Yang, K.; Qian, G.; Ge, C. The effects of estrogen on gonadal differentiation and expressions of DMRT1 and SOX9 in Pelodiscus sinensis. Acta Hydrobiol. Sin. 2014, 38, 467–473. [Google Scholar]
- Liang, H.; Meng, Y.; Cao, L.; Li, X.; Zou, G. Effect of exogenous hormones on R-spondin 1 (RSPO1) gene expression and embryo development in Pelodiscus sinensis. Reprod. Fertil. Dev. 2019, 31, 1425–1433. [Google Scholar] [CrossRef] [Green Version]
- Kopania, E.; Larson, E.; Callahan, C.; Keeble, S.; Good, J. Molecular evolution across mouse spermatogenesis. Mol. Biol. Evol. 2022, 39, msac023. [Google Scholar] [CrossRef]
- Geisinger, A.; Alsheimer, M.; Baier, A.; Benavente, R.; Wettstein, R. The mammalian gene pecanex 1 is differentially expressed during spermatogenesis. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 2005, 1728, 34–43. [Google Scholar] [CrossRef]
- Geisinger, A.; Dos Santos, A.; Benavente, R.; Wettstein, R. Identification and characterization of Srsp1, a rat gene differentially expressed during spermatogenesis and coding for a serine stretch-containing protein. Cytogenet. Genome Res. 2002, 98, 249–254. [Google Scholar] [CrossRef]
- Capoano, C.A.; Wettstein, R.; Kun, A.; Geisinger, A. Spats 1 (Srsp1) is differentially expressed during testis development of the rat. Gene Expr. Patterns 2010, 10, 1–8. [Google Scholar] [CrossRef]
- da Cruz, I.; Rodríguez-Casuriaga, R.; Santiñaque, F.F.; Farías, J.; Curti, G.; Capoano, C.A.; Folle, G.A.; Benavente, R.; Sotelo-Silveira, J.R.; Geisinger, A. Transcriptome analysis of highly purified mouse spermatogenic cell populations: Gene expression signatures switch from meiotic-to postmeiotic-related processes at pachytene stage. BMC Genom. 2016, 17, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouawad, N. SPATS1: New Molecular Target in Colorectal Cancer. Ph.D. Thesis, University of Pisa, Pisa, Italy, 2021. [Google Scholar]
- Zhu, J.; Lei, L.; Chen, C.; Wang, Y.; Liu, X.; Geng, L.; Li, R.; Chen, H.; Hong, X.; Yu, L.; et al. Whole-Transcriptome Analysis Identifies Gender Dimorphic Expressions of Mrnas and Non-Coding Rnas in Chinese Soft-Shell Turtle (Pelodiscus sinensis). Biology 2022, 11, 834. [Google Scholar] [CrossRef]
- Li, H.; Zhou, Z.; Wu, T.; Wu, Y.; Ji, X. Do fluctuations in incubation temperature affect hatchling quality in the Chinese soft-shelled turtle Pelodiscus sinensis? Aquaculture 2013, 406, 91–96. [Google Scholar] [CrossRef]
- Du, W.G.; Ji, X. The effects of incubation thermal environments on size, locomotor performance and early growth of hatchling soft-shelled turtles, Pelodiscus sinensis. J. Therm. Biol. 2003, 28, 279–286. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, L.; Sun, W.; Li, P.; Zhou, Y.; Qian, G.; Ge, C. Knockdown of R-spondin1 leads to partial sex reversal in genetic female Chinese soft-shelled turtle Pelodiscus sinensis. Gen. Comp. Endocrinol. 2021, 309, 113788. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, L.; Sun, W.; Li, P.; Zhou, Y.; Qian, G.; Ge, C. Characterization of the Pelodiscus sinensis polymeric immunoglobulin receptor (P. sinensis pIgR) and its response to LPS and Aeromonas sobria. Dev. Comp. Immunol. 2021, 121, 104072. [Google Scholar]
- Cai, H.; Sun, W.; Liu, H.; Bao, H.; Zhang, H.; Qian, G.; Ge, C. Molecular cloning and expression pattern of Amh and its preliminary functional analysis in male sexual differentiation in Pelodiscus sinensis. Sci. Sin. Vitae 2016, 46, 1423–1433. [Google Scholar]
- Tokita, M.; Kuratani, S. Normal embryonic stages of the Chinese softshelled turtle Pelodiscus sinensis (Trionychidae). Zool. Sci. 2001, 18, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhu, X.; Xu, H.; Zhao, J.; Chen, C. A PCR Amplification Primer, Method and Kit for Rapid Identification of Genetic Traits in the Chinese Soft-shelled Turtle. China Patent CN108841945B, 5 May 2020. [Google Scholar]
- Sun, W.; Cai, H.; Zhang, G.; Zhang, H.; Bao, H.; Wang, L.; Ye, J.; Qian, G.; Ge, C. Dmrt1 is required for primary male sexual differentiation in Chinese soft-shelled turtle Pelodiscus sinensis. Sci. Rep. 2017, 7, 4433. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.Y.; Mruk, D.D. The biology of spermatogenesis: The past, present and future. Philos. Trans. R Soc. Lond. B Biol. Sci. 2020, 365, 1459–1463. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Agarwal, A. Spermatogenesis: An overview. Sperm Chromatin 2011, 19–44. [Google Scholar] [CrossRef]
- Johnson, L. Efficiency of spermatogenesis. Microsc. Res. Technol. 1995, 32, 385–422. [Google Scholar] [CrossRef] [PubMed]
- Saunders, P. Germ cell-somatic cell interactions during spermatogenesis. Reprod. Suppl. 2003, 61, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Capoano, C.A.; Ortiz-Laquintana, L.A.; Rodríguez-Casuriaga, R.; Schlapp, G.; Meikle, M.N.; Mulet, A.P.; Crispo, M.; Benavente, R.; Geisinger, A. SPATS1 (spermatogenesis-associated, serine-rich 1) is not essential for spermatogenesis and fertility in mouse. PLoS ONE 2021, 16, e0251028. [Google Scholar] [CrossRef]
- Jiménez, R.; Burgos, M.; Barrionuevo, F.J. Circannual testis changes in seasonally breeding mammals. Sex. Dev. 2015, 9, 205–215. [Google Scholar] [CrossRef]
- Mahmoud, I.; Licht, P. Seasonal changes in gonadal activity and the effects of stress on reproductive hormones in the common snapping turtle, Chelydra serpentina. Gen. Comp. Endocrinol. 1997, 107, 359–372. [Google Scholar] [CrossRef]
- Ribeiro, Y.M.; de Matos, S.A.; Domingos, F.F.T.; Dos Santos, H.B.; Vieira, A.B.C.; Bazzoli, N.; Rizzo, E. Germ cell proliferation and apoptosis during testicular regression in a seasonal breeding fish kept in captivity. Tissue Cell 2017, 49, 664–671. [Google Scholar] [CrossRef]
- Lofts, B. Patterns of spermatogenesis and steroidogenesis in male reptiles. In Reproduction and Evolution; Calaby, J.H., Tyndale-Boscoe, C.H., Eds.; Australian Academic Science: Canberra, Australia, 1977; pp. 127–136. [Google Scholar]
- Radder, R.; Shanbhag, B.; Saidapur, S. Pattern of plasma sex steroid hormone levels during reproductive cycles of male and female tropical lizard, Calotes versicolor. Gen. Comp. Endocrinol. 2001, 124, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Li, H.; Zhang, G.; Kang, X.; Li, Q.; Cao, J.; Xu, E. Serum sex hormonal levels and gonadal development in broodstock soft-shelled turtle Trionyx sinensis in a green house. Fish. Sci. 2012, 31, 264–269. [Google Scholar]
- Zhang, L.; Han, X.K.; Qi, Y.Y.; Liu, Y.; Chen, Q.S. Seasonal effects on apoptosis and proliferation of germ cells in the testes of the Chinese soft-shelled turtle, Pelodiscus sinensis. Theriogenology 2008, 69, 1148–1158. [Google Scholar] [CrossRef]
- Li, W.; Zhu, J.; Lei, L.; Chen, C.; Liu, X.; Wang, Y.; Hong, X.; Xu, H.; Zhu, X. The Seasonal and Stage-Specific Expression Patterns of HMGB2 Suggest Its Key Role in Spermatogenesis in the Chinese Soft-Shelled Turtle (Pelodiscus sinensis). Biochem. Genet. 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sukhan, Z.P.; Cho, Y.; Sharker, M.R.; Hossen, S.; Rha, S.J.; Kho, K.H. Effective accumulative temperature affects gonadal maturation by controlling expression of GnRH, GnRH receptor, serotonin receptor and APGWamide gene in Pacific abalone, Haliotis discus hannai during broodstock conditioning in hatcheries. J. Therm. Biol. 2021, 100, 103037. [Google Scholar] [CrossRef] [PubMed]
- Breitenbach, A.T.; Bowden, R.M.; Paitz, R.T. Effects of Constant and Fluctuating Temperatures on Gene Expression During Gonadal Development. Integr. Comp. Biol. 2022, icac011. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, M.; Mileo, A.; Laforgia, V.; De Falco, M.; Rosati, L. Alkyphenol Exposure Alters Steroidogenesis in Male Lizard Podarcis siculus. Animals 2021, 11, 1003. [Google Scholar] [CrossRef] [PubMed]
- Othman, R.; Ron, X.J.; Yao, H.; O’Bryant, P.; Rapp, D.; Pei, J.C.; Pei, J.C.; Wang, H.P. The effect of methyltestosterone (MT) on sex differentiation and growth in juvenile yellow perch (Perca flavescens). Fish Physiol. Biochem. 2022, 48, 161–171. [Google Scholar] [CrossRef]
- Cai, Y.; He, Z.; Zhang, Q.; Wang, X.; Zhang, S.; He, L.; Yang, D.; Zhang, M.; Yan, T. Oestradiol upregulates the expression of Cyp19a1a and the feminization ratio in Schizothorax prenanti. Aquac. Rep. 2022, 24, 101138. [Google Scholar] [CrossRef]
- Gardner, L.; Anderson, T.; Place, A.R.; Dixon, B.; Elizur, A. Sex change strategy and the aromatase genes. J. Steroid Biochem. Mol. Biol. 2005, 94, 395–404. [Google Scholar] [CrossRef]
- Jiang, Y.; Luo, H.; Hou, M.; Chen, J.; Tao, B.; Zhu, Z.; Song, Y.; Hu, W. Aromatase inhibitor induces sex reversal in the protogynous hermaphroditic rice field eel (Monopterus albus). Aquaculture 2022, 551, 737960. [Google Scholar] [CrossRef]
- Zhu, Q.; Han, C.; Liu, S.; Ouyang, H.; Liu, D.; Zhang, Z.; Huang, J.; Han, L.; Li, S.; Li, G. Development and gene expression analysis of gonad during 17α-methyltestosterone-induced sex reversal in mandarin fish (Siniperca chuatsi). Aquac. Rep. 2022, 23, 101049. [Google Scholar] [CrossRef]
- Li, P.; Guo, Y.; Jin, L.; Liang, X.; Chen, G.; Sun, W.; Xiao, L.; Qian, G.; Ge, C. ESR1 mediates estrogen-induced feminization of genetic male Chinese soft-shelled turtle. Biol. Reprod. 2022, ioac088. [Google Scholar] [CrossRef]
- Banh, Q.Q.; Guppy, J.L.; Domingos, J.A.; Budd, A.M.; Pinto, R.C.; Marc, A.F.; Jerry, D.R. Induction of precocious females in the protandrous barramundi (Lates calcarifer) through implants containing 17β-estradiol-effects on gonadal morphology, gene expression and DNA methylation of key sex genes. Aquaculture 2021, 539, 736601. [Google Scholar] [CrossRef]
- Weber, G.M.; Leeds, T.D.; Schneider, R.P. Sex reversal of female rainbow trout by immersion in 17α-methyltestosterone. Aquaculture 2020, 528, 735535. [Google Scholar] [CrossRef]
- Delbes, G.; Blázquez, M.; Fernandino, J.I.; Grigorova, P.; Hales, B.F.; Metcalfe, C.; Navarro-Martín, L.; Parent, L.; Robaire, B.; Rwigemera, A.; et al. Effects of endocrine disrupting chemicals on gonad development: Mechanistic insights from fish and mammals. Environ. Res. 2022, 204, 112040. [Google Scholar] [CrossRef] [PubMed]
- Spaan, K.; Haigis, A.C.; Weiss, J.; Legradi, J. Effects of 25 thyroid hormone disruptors on zebrafish embryos: A literature review of potential biomarkers. Sci. Total Environ. 2019, 656, 1238–1249. [Google Scholar] [CrossRef]
- Darras, V.M. The role of maternal thyroid hormones in avian embryonic development. Front. Endocrinol. 2019, 10, 66. [Google Scholar] [CrossRef]
- Liang, H.W.; Meng, Y.; Cao, L.H.; Li, X.; Zou, G.W. Expression and characterization of the cyp19a gene and its responses to estradiol/letrozole exposure in Chinese soft-shelled turtle (Pelodiscus sinensis). Mol. Reprod. Dev. 2019, 86, 480–490. [Google Scholar] [CrossRef]
- Dzyuba, V.; Cosson, J.; Papadaki, M.; Mylonas, C.C.; Steinbach, C.; Rodina, M.; Tučkova, V.; Linhart, O.; Shelton, W.; Gela, G. Influence of Environmental Temperature and Hormonal Stimulation on the In Vitro Sperm Maturation in Sterlet Acipenser ruthenus in Advance of the Spawning Season. Animals 2021, 11, 1417. [Google Scholar] [CrossRef]
- Dostalova, P.; Zatecka, E.; Dvorakova-Hortova, K. Of oestrogens and sperm: A review of the roles of oestrogens and oestrogen receptors in male reproduction. Int. J. Mol. Sci. 2017, 18, 904. [Google Scholar] [CrossRef] [Green Version]
- Komrsková, K. The Physiology and Reproductive Potential of Sperm. Ph.D. Thesis, Charles University, Prague, Czech Republic, 2021. [Google Scholar]
- Gao, L.L.; Diao, X.M.; Li, Y.; Zhai, X.L.; Zhou, C.L. Molecular cloning and expression of foxl2 gene induced by exogenous hormone in the pelodiscus sinensis. Acta Hydrobiol. Sin. 2019, 43, 7. [Google Scholar]


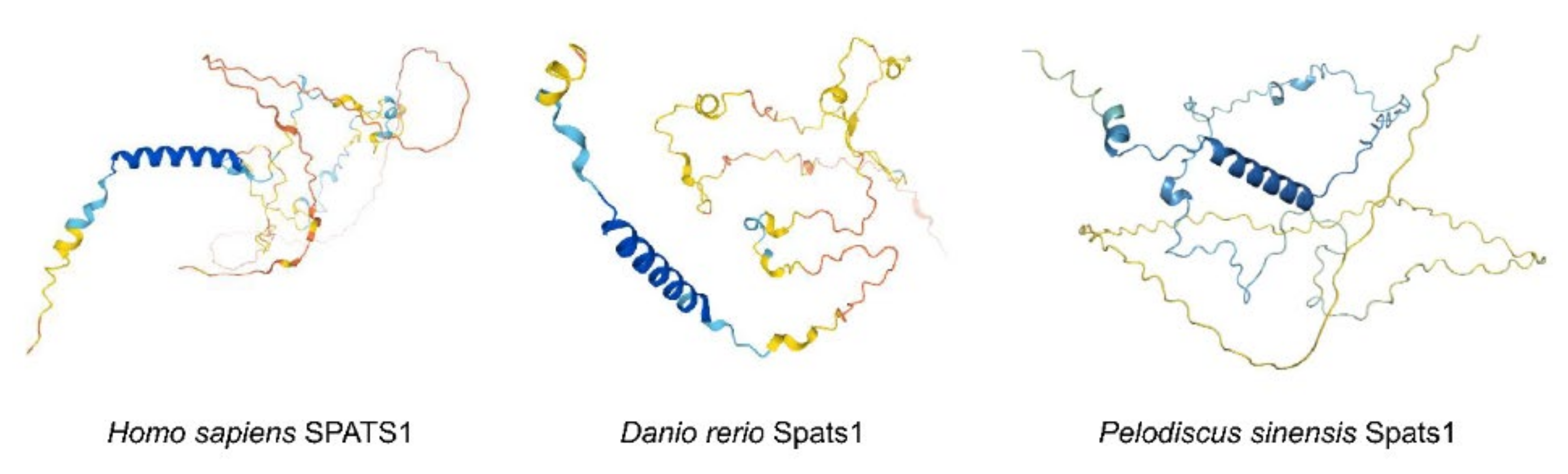
 : Very high (Plddt > 90);
: Very high (Plddt > 90);  : Confident (90 > Plddt > 70);
: Confident (90 > Plddt > 70);  : Low (70 > Plddt > 50);
: Low (70 > Plddt > 50);  : Very low (Plddt < 50).
: Very high (Plddt > 90); : Confident (90 > Plddt > 70); : Low (70 > Plddt > 50); : Very low (Plddt < 50).
: Very low (Plddt < 50).
: Very high (Plddt > 90); : Confident (90 > Plddt > 70); : Low (70 > Plddt > 50); : Very low (Plddt < 50).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Application | Sequences (5′—3′) | Amplification Efficiency | Product Size (bp) |
|---|---|---|---|---|
| PsSpats1-1F | Clone | CCCTGAATCATGACCCCTCA | 1201 | |
| PsSpats1-1R | Clone | TCCATGGAAACAACCACTGTGAA | ||
| PsSpats1-2F | Semi-Quantitative/qRT-PCR | AGCGAGGGAAGTAGCAT | 98.25% | 251 |
| Ps Spats1-2R | Semi-Quantitative/qRT-PCR | GGAAGGTGAAAGGGAAAA | ||
| Ps Spats1-3F | CISH | TAATACGACTCACTATAG GGCGAGCCGACCTTCTACCCA | 869 | |
| Ps Spats1-3R | CISH | ATTTAGGTGACACTATAGAA TACTCTCTGTCCTTTCGCATTA | ||
| Eef1a-F | Semi-Quantitative/qRT-PCR | ACTCGTCCAACTGACAAGCCTC | 100.73% | 337 |
| Eef1a-R | Semi-Quantitative/qRT-PCR | CACGGCGAACATCTTTCACAG | ||
| ZHB1-F | Sex identification PCR | CTGGAAACAATATCATCGCCGAG | 590 | |
| ZHB1-R | Sex identification PCR | TGTGTGCCGTGCCTGCGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, L.; Zhu, J.; Chen, C.; Wang, Y.; Hong, X.; Liu, X.; Yu, L.; Wei, C.; Chen, H.; Liu, Y.; et al. Expression and Characterization of the Spats1 Gene and Its Response to E2/MT Treatment in the Chinese Soft-Shelled Turtle (Pelodiscus sinensis). Animals 2022, 12, 1858. https://doi.org/10.3390/ani12141858
Lei L, Zhu J, Chen C, Wang Y, Hong X, Liu X, Yu L, Wei C, Chen H, Liu Y, et al. Expression and Characterization of the Spats1 Gene and Its Response to E2/MT Treatment in the Chinese Soft-Shelled Turtle (Pelodiscus sinensis). Animals. 2022; 12(14):1858. https://doi.org/10.3390/ani12141858
Chicago/Turabian StyleLei, Luo, Junxian Zhu, Chen Chen, Yakun Wang, Xiaoyou Hong, Xiaoli Liu, Lingyun Yu, Chengqing Wei, Haigang Chen, Yihui Liu, and et al. 2022. "Expression and Characterization of the Spats1 Gene and Its Response to E2/MT Treatment in the Chinese Soft-Shelled Turtle (Pelodiscus sinensis)" Animals 12, no. 14: 1858. https://doi.org/10.3390/ani12141858