
Effects of Birthweight of Piglets in a Multi-Suckling System on Mortality, Growth Rate, Catch-Up Growth, Feed Intake and Behaviour
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
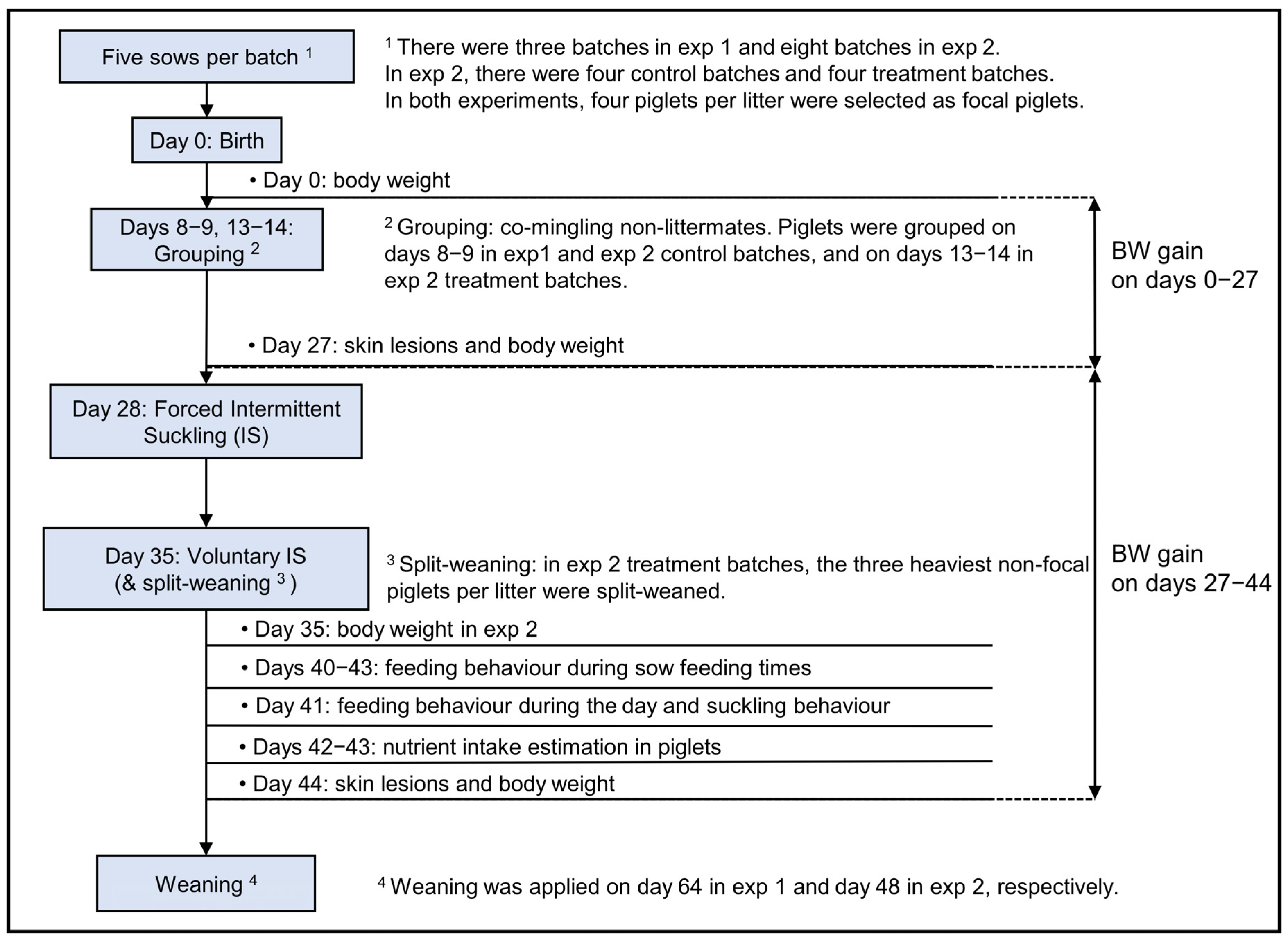
2. Materials and Methods
2.1. Animals and Experimental Design
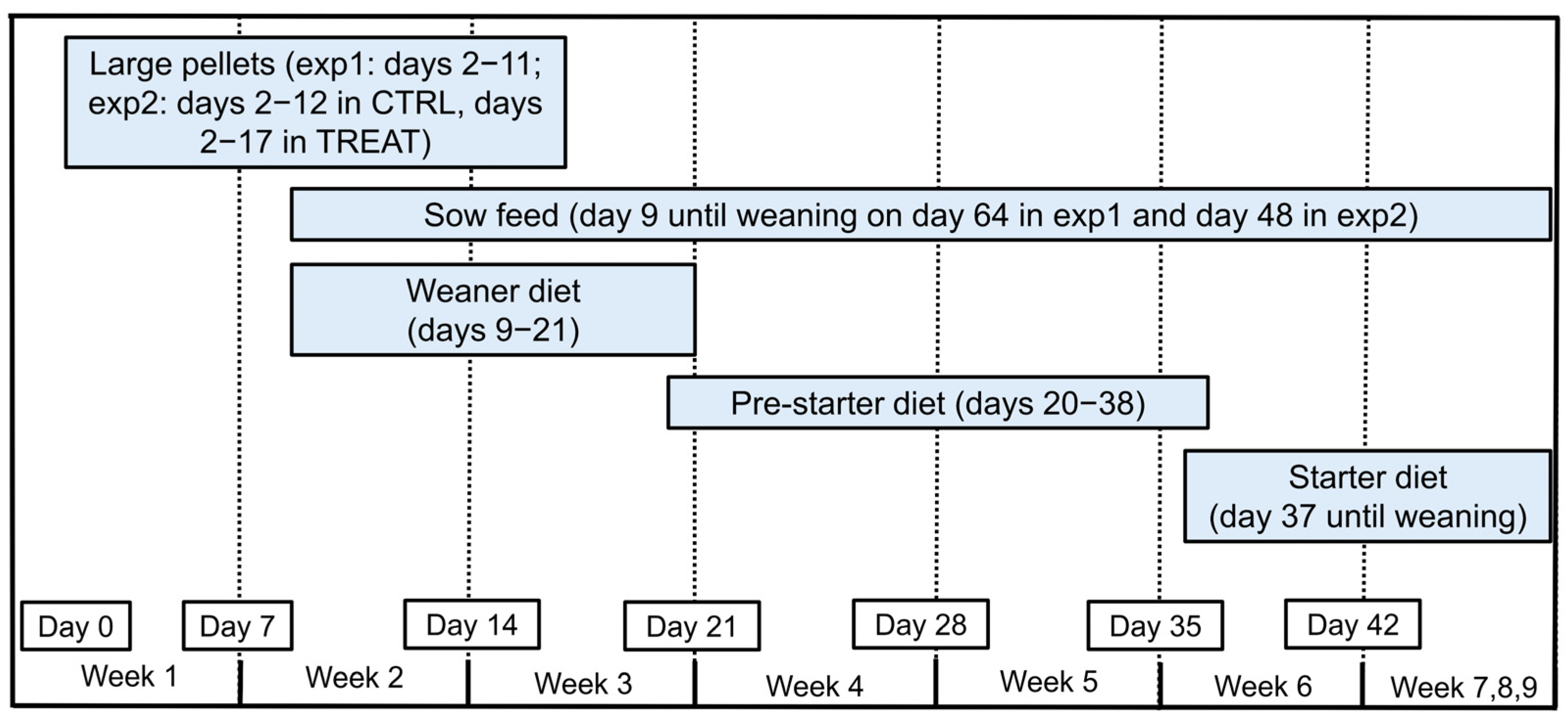
2.2. General Housing and Feeding Management
2.3. Data Collection from Exp 1 and Exp 2
2.3.1. Body Weight
2.3.2. Feeding Behaviour during Sow Feeding Times and during the Day
2.3.3. Suckling Behaviour
2.3.4. Skin Lesions
2.3.5. Individual Nutrient Intake
2.3.6. Piglet Mortality
2.4. The Definition of Fast and Slow-Growing Piglets
2.5. Statistics
3. Results
3.1. Birthweight and Mortality
3.2. Birthweight and Piglet Traits
3.3. The Effect of Growth Rate Class and Birthweight Class on Piglet Traits
4. Discussion
4.1. Birthweight and Mortality
4.2. Birthweight and Piglet Traits
4.3. The Effect of Growth Rate Class and Birthweight Class on Piglet Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moreira, R.H.R.; Pérez Palencia, J.Y.; Moita, V.H.C.; Caputo, L.S.S.; Saraiva, A.; Andretta, I.; Ferreira, R.A.; de Abreu, M.L.T. Variability of piglet birth weights: A systematic review and meta-analysis. J. Anim. Physiol. Anim. 2020, 104, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Knol, E.F.; van der Spek, D.; Zak, L.J. Genetic aspects of piglet survival and related traits: A review. J. Anim. Sci. 2022, 100, skac190. [Google Scholar] [CrossRef]
- Ward, S.A.; Kirkwood, R.N.; Plush, K.J. Are larger litters a concern for piglet survival or an effectively manageable trait? Animals 2020, 10, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, S.P.; Jansman, A.J.M.; Verstegen, M.W.A.; Awati, A.; Buist, W.; Den Hartog, L.A.; Van Hees, H.M.J.; Quiniou, N.; Hendriks, W.H.; Gerrits, W.J.J. Analysis of factors to predict piglet body weight at the end of the nursery phase. J. Anim. Sci. 2012, 90, 3243–3251. [Google Scholar] [CrossRef] [PubMed]
- Douglas, S.L.; Edwards, S.A.; Kyriazakis, I. Management strategies to improve the performance of low birth weight pigs to weaning and their long-term consequences. J. Anim. Sci. 2014, 92, 2280–2288. [Google Scholar] [CrossRef] [PubMed]
- Van Nieuwamerongen, S.E.; Soede, N.M.; Van der Peet-Schwering, C.M.C.; Kemp, B.; Bolhuis, J.E. Gradual weaning during an extended lactation period improves performance and behavior of pigs raised in a multi-suckling system. Appl. Anim. Behav. Sci. 2017, 194, 24–35. [Google Scholar] [CrossRef]
- Thomsson, O.; Magnusson, U.; Bergqvist, A.S.; Eliasson-Selling, L.; Sjunnesson, Y.C.B. Sow performance in multi-suckling pens with different management routines. Acta Vet. Scand. 2018, 60, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Lodge, G.A.; McDonald, I. The relative influence of birth weight, milk consumption and supplementary food consumption upon the growth rates of suckling piglets. Anim. Sci. 1959, 1, 139–144. [Google Scholar] [CrossRef]
- Thompson, B.K.; Fraser, D. Variation in piglet weights: Development of within-litter variation over a 5-week lactation and effect of farrowing crate design. Can. J. Anim. Sci. 1986, 66, 361–372. [Google Scholar] [CrossRef]
- Fraser, D.; Pajor, E.A.; Feddes, J.J.R. The relationship between creep feeding behavior of piglets and adaptation to weaning: Effect of diet quality. Can. J. Anim. Sci. 1994, 74, 1–6. [Google Scholar] [CrossRef]
- Muns, R.; Manzanilla, E.G.; Sol, C.; Manteca, X.; Gasa, J. Piglet behavior as a measure of vitality and its influence on piglet survival and growth during lactation. J. Anim. Sci. 2013, 91, 1838–1843. [Google Scholar] [CrossRef] [Green Version]
- Wiegert, J.G.; Garrison, C.; Knauer, M.T. Characterization of birth weight and colostrum intake on piglet survival and piglet quality. J. Anim. Sci. 2017, 95, 32. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Gerrits, W.J.J.; Reimert, I.; van der Peet-Schwering, C.; Soede, N.M. Variation in piglet body weight gain and feed intake during a 9-week lactation in a multi-suckling system. Animal 2022, 16, 100651. [Google Scholar] [CrossRef]
- Milligan, B.N.; Fraser, D.; Kramer, D.L. Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest. Prod. Sci. 2002, 76, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Quiniou, N.; Dagorn, J.; Gaudré, D. Variation of piglets’ birth weight and consequences on subsequent performance. Livest. Prod. Sci. 2002, 78, 63–70. [Google Scholar] [CrossRef]
- Hales, J.; Moustsen, V.A.; Nielsen, M.B.F.; Hansen, C.F. Individual physical characteristics of neonatal piglets affect preweaning survival of piglets born in a noncrated system. J. Anim. Sci. 2013, 91, 4991–5003. [Google Scholar] [CrossRef] [Green Version]
- Van Nieuwamerongen, S.E.; Soede, N.M.; Van der Peet-Schwering, C.M.C.; Kemp, B.; Bolhuis, J.E. Development of piglets raised in a new multi-litter housing system vs. conventional single-litter housing until 9 weeks of age. J. Anim. Sci. 2015, 93, 5442–5454. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Kuhn, G. Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis. J. Anim. Sci. 2006, 84, E113–E123. [Google Scholar] [CrossRef]
- Beaulieu, A.D.; Aalhus, J.L.; Williams, N.H.; Patience, J.F. Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork. J. Anim. Sci. 2010, 88, 2767–2778. [Google Scholar] [CrossRef] [PubMed]
- Douglas, S.L.; Edwards, S.A.; Sutcliffe, E.; Knap, P.W.; Kyriazakis, I. Identification of risk factors associated with poor lifetime growth performance in pigs. J. Anim. Sci. 2013, 91, 4123–4132. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.G.; Bee, G. Compensatory growth feeding strategy does not overcome negative effects on growth and carcass composition of low birth weight pigs. Animal 2015, 9, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, S.L.; Edwards, S.A.; Kyriazakis, I. Are all piglets born lightweight alike? Morphological measurements as predictors of postnatal performance. J. Anim. Sci. 2016, 94, 3510–3518. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Gerrits, W.J.J.; Soede, N.M.; van der Peet-Schwering, C.M.C.; Reimert, I. Effects of timing of grouping and split-weaning on growth performance and behaviour of piglets in a multi-suckling system. Appl. Anim. Behav. Sci. 2023; under review. [Google Scholar]
- Weary, D.M.; Pajor, E.A.; Bonenfant, M.; Fraser, D.; Kramer, D.L. Alternative housing for sows and litters: Part 4. Effects of sow-controlled housing combined with a communal piglet area on pre-and post-weaning behaviour and performance. Appl. Anim. Behav. Sci. 2002, 76, 279–290. [Google Scholar] [CrossRef]
- Turner, S.P.; Farnworth, M.J.; White, I.M.; Brotherstone, S.; Mendl, M.; Knap, P.; Penny, P.; Lawrence, A.B. The accumulation of skin lesions and their use as a predictor of individual aggressiveness in pigs. Appl. Anim. Behav. Sci. 2006, 96, 245–259. [Google Scholar] [CrossRef]
- Tang, T.; van der Peet-Schwering, C.M.C.; Soede, N.M.; Laurenssen, B.F.A.; Bruininx, E.M.A.M.; Bos, E.J.; Gerrits, W.J.J. A dual marker technique to estimate individual feed intake in young pigs. Animal 2022, 16, 100451. [Google Scholar] [CrossRef]
- Smit, H.J.; Taweel, H.Z.; Tas, B.M.; Tamminga, S.; Elgersma, A. Comparison of techniques for estimating herbage intake of grazing dairy cows. J. Dairy Sci. 2005, 88, 1827–1836. [Google Scholar] [CrossRef]
- Theil, P.K.; Nielsen, T.T.; Kristensen, N.B.; Labouriau, R.; Danielsen, V.; Lauridsen, C.; Jakobsen, K. Estimation of milk production in lactating sows by determination of deuterated water turnover in three piglets per litter. Acta Agric. Scand. A Anim. Sci. 2002, 52, 221–232. [Google Scholar] [CrossRef]
- Hurley, W.L. Composition of sow colostrum and milk. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 193–230. [Google Scholar]
- Stroup, W.W.; Milliken, G.A.; Claassen, E.A.; Wolfinger, R.D. SAS for Mixed Models: Introduction and Basic Applications; SAS Institute Inc.: Cary, NC, USA, 2018. [Google Scholar]
- Thomsson, O.; Sjunnesson, Y.; Magnusson, U.; Eliasson-Selling, L.; Wallenbeck, A.; Bergqvist, A.S. Consequences for piglet performance of group housing lactating sows at one, two, or three weeks post-farrowing. PLoS ONE 2016, 11, e0156581. [Google Scholar] [CrossRef] [Green Version]
- Kirkden, R.D.; Broom, D.M.; Andersen, I.L. Invited review: Piglet mortality: Management solutions. J. Anim. Sci. 2013, 91, 3361–3389. [Google Scholar] [CrossRef]
- Heuß, E.M.; Pröll-Cornelissen, M.J.; Neuhoff, C.; Tholen, E.; Große-Brinkhaus, C. Invited review: Piglet survival: Benefits of the immunocompetence. Animal 2019, 13, 2114–2124. [Google Scholar] [CrossRef]
- Vande Pol, K.D.; Bautista, R.O.; Harper, H.; Shull, C.M.; Brown, C.B.; Ellis, M. Effect of within-litter birth weight variation after cross-fostering on piglet preweaning growth and mortality. Transl. Anim. Sci. 2021, 5, txab039. [Google Scholar] [CrossRef] [PubMed]
- Verdon, M.; Morrison, R.S.; Rault, J.L. The welfare and productivity of sows and piglets in group lactation from 7, 10, or 14 d postpartum. J. Anim. Sci. 2020, 98, skaa037. [Google Scholar] [CrossRef] [PubMed]
- Marchant, J.N.; Rudd, A.R.; Mendl, M.T.; Broom, D.M.; Meredith, M.J.; Corning, S.; Simmins, P.H. Timing and causes of piglet mortality in alternative and conventional farrowing systems. Vet. Rec. 2000, 147, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, L.J.; Berg, P.; Jørgensen, G.; Andersen, I.L. Neonatal piglet traits of importance for survival in crates and indoor pens. J. Anim. Sci. 2011, 89, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Herpin, P.; Vincent, A.; Damon, M. Effect of breed and body weight on thermoregulatory abilities of European (Pie-train×(Landrace× Large White)) and Chinese (Meishan) piglets at birth. Livest. Prod. Sci. 2004, 88, 17–26. [Google Scholar] [CrossRef]
- Kammersgaard, T.S.; Pedersen, L.J.; Jørgensen, E. Hypothermia in neonatal piglets: Interactions and causes of individual differences. J. Anim. Sci. 2011, 89, 2073–2085. [Google Scholar] [CrossRef] [PubMed]
- Vanden Hole, C.; Aerts, P.; Prims, S.; Ayuso, M.; Van Cruchten, S.; Van Ginneken, C. Does intrauterine crowding affect locomotor development? A comparative study of motor performance, neuromotor maturation and gait variability among piglets that differ in birth weight and vitality. PLoS ONE 2018, 13, e0195961. [Google Scholar] [CrossRef] [Green Version]
- Herpin, P.; Damon, M.; Le Dividich, J. Development of thermoregulation and neonatal survival in pigs. Livest. Prod. Sci. 2002, 78, 25–45. [Google Scholar] [CrossRef]
- Weary, D.M.; Pajor, E.A.; Thompson, B.K.; Fraser, D. Risky behaviour by piglets: A trade off between feeding and risk of mortality by maternal crushing? Anim. Behav. 1996, 51, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Fraser, D. Behavioural perspectives on piglet survival. J. Reprod. Fertil. Suppl. 1990, 40, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Andersen, I.L.; Nævdal, E.; Bøe, K.E. Maternal investment, sibling competition, and offspring survival with increasing litter size and parity in pigs (Sus scrofa). Behav. Ecol. Sociobiol. 2011, 65, 1159–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oostindjer, M.; Bolhuis, J.E.; Mendl, M.; Held, S.; van den Brand, H.; Kemp, B. Learning how to eat like a pig: Effectiveness of mechanisms for vertical social learning in piglets. Anim. Behav. 2011, 82, 503–511. [Google Scholar] [CrossRef]
- Van der Peet-Schwering, C.M.C.; Troquet, L.M.P.; Binnendijk, G.P.; Knol, E. Effect of Genetic Background and Birth Weight on Performance of Piglets and Growing and Finishing Pigs; Wageningen Livestock Research: Wageningen, The Netherlands, 2013. [Google Scholar]
- Alvarenga, A.L.N.; Chiarini-Garcia, H.; Cardeal, P.C.; Moreira, L.P.; Foxcroft, G.R.; Fontes, D.O.; Almeida, F.R.C.L. Intra-uterine growth retardation affects birthweight and postnatal development in pigs, impairing muscle accretion, duodenal mucosa morphology and carcass traits. Reprod. Fertil. Dev. 2013, 25, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Van der Peet-Schwering, C.M.C.; Verschuren, L.M.; Bergsma, R.; Hedemann, M.S.; Binnendijk, G.P.; Jansman, A.J. The effects of birth weight and estimated breeding value for protein deposition on nitrogen efficiency in growing pigs. J. Anim. Sci. 2021, 99, skab101. [Google Scholar] [CrossRef]
- Michiels, J.; De Vos, M.; Missotten, J.; Ovyn, A.; De Smet, S.; Van Ginneken, C. Maturation of digestive function is retarded and plasma antioxidant capacity lowered in fully weaned low birth weight piglets. Br. J. Nutr. 2013, 109, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, J.C.; Thomson, W. Observations on the behaviour of suckling pigs. Brit. J. Anim. Behav. 1956, 4, 46–51. [Google Scholar] [CrossRef]
- Fraser, D.; Nicholls, C.; Fagan, W. A sow milking machine designed to compare the yield of different teats. J. Agric. Eng. Res. 1985, 31, 371–376. [Google Scholar] [CrossRef]
- Skok, J.; Brus, M.; Škorjanc, D. Growth of piglets in relation to milk intake and anatomical location of mammary glands. Acta Agric. Scand. A. 2007, 57, 129–135. [Google Scholar] [CrossRef]
- Ogawa, S.; Tsukahara, T.; Tsuruta, T.; Nishibayashi, R.; Okutani, M.; Nakatani, M.; Higashide, K.; Iida, S.; Nakanishi, N.; Ushida, K.; et al. The evaluation of secretion volume and immunoglobulin A and G concentrations in sow colostrum from anterior to posterior teats. Anim. Sci. J. 2014, 85, 678–682. [Google Scholar] [CrossRef]
- Hurley, W.L. Mammary gland growth in the lactating sow. Livest. Prod. Sci. 2001, 70, 149–157. [Google Scholar] [CrossRef]
- Hartsock, T.G.; Graves, H.B.; Baumgardt, B.R. Agonistic behavior and the nursing order in suckling piglets: Relationships with survival, growth and body composition. J. Anim. Sci. 1977, 44, 320–330. [Google Scholar] [CrossRef]
- Farmer, C. Mammary development in lactating sows: The importance of suckling. Animal 2019, 13, s20–s25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevara, R.D.; Pastor, J.J.; Manteca, X.; Tedo, G.; Llonch, P. Systematic review of animal-based indicators to measure thermal, social, and immune-related stress in pigs. PLoS ONE 2022, 17, e0266524. [Google Scholar] [CrossRef]
- Van Kerschaver, C.; Vandaele, M.; Degroote, J.; Van Tichelen, K.; Fremaut, D.; Van Ginneken, C.; Michiels, J. Effect of starting time of co-mingling non-littermates during lactation on performance and skin lesions of sows and piglets. Livest. Sci. 2021, 250, 104563. [Google Scholar] [CrossRef]
- Bernardino, T.; Tatemoto, P.; Morrone, B.; Mazza Rodrigues, P.H.; Zanella, A.J. Piglets born from sows fed high fibre diets during pregnancy are less aggressive prior to weaning. PLoS ONE 2016, 11, e0167363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, S.P.; Jansman, A.J.M.; Verstegen, M.W.A.; Den Hartog, L.A.; Van Hees, H.M.J.; Bolhuis, J.E.; Van Kempen, T.A.T.G.; Gerrits, W.J.J. Identifying the limitations for growth in low performing piglets from birth until 10 weeks of age. Animal 2014, 8, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Hoy, S.; Schamun, S.; Weirich, C. Investigations on feed intake and social behaviour of fattening pigs fed at an electronic feeding station. Appl. Anim. Behav. Sci. 2012, 139, 58–64. [Google Scholar] [CrossRef]
- Nielsen, B.L.; De Jong, I.C.; Vries, T.J.D. The use of feeding behaviour in the assessment of animal welfare. In Nutrition and the Welfare of Farm Animals; Phillips, C.J.C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 59–84. [Google Scholar]
- Valros, A.; Sali, V.; Hälli, O.; Saari, S.; Heinonen, M. Does weight matter? Exploring links between birth weight, growth and pig-directed manipulative behaviour in growing-finishing pigs. Appl. Anim. Behav. Sci. 2021, 245, 105506. [Google Scholar] [CrossRef]
- Algers, B.; Jensen, P.; Steinwall, L. Behaviour and weight changes at weaning and regrouping of pigs in relation to teat quality. Appl. Anim. Behav. Sci. 1990, 26, 143–155. [Google Scholar] [CrossRef]
- Yang, C.H.; Ko, H.L.; Salazar, L.C.; Llonch, L.; Manteca, X.; Camerlink, I.; Llonch, P. Pre-weaning environmental enrichment increases piglets’ object play behaviour on a large scale commercial pig farm. Appl. Anim. Behav. Sci. 2018, 202, 7–12. [Google Scholar] [CrossRef]
- Bus, J.D.; Boumans, I.J.; Webb, L.E.; Bokkers, E.A. The potential of feeding patterns to assess generic welfare in growing-finishing pigs. Appl. Anim. Behav. Sci. 2021, 241, 105383. [Google Scholar] [CrossRef]
- Olsen, A.N.W.; Dybkjaer, L.; Vestergaard, K.S. Cross-suckling and associated behaviour in piglets and sows. Appl. Anim. Behav. Sci. 1998, 61, 13–24. [Google Scholar] [CrossRef]
- Quesnel, H.; Farmer, C.; Theil, P.K. Colostrum and milk production. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 173–192. [Google Scholar]
- Berkeveld, M.; Langendijk, P.; van Beers-Schreurs, H.M.; Koets, A.P.; Taverne, M.A.; Verheijden, J.H. Postweaning growth check in pigs is markedly reduced by intermittent suckling and extended lactation. J. Anim. Sci. 2007, 85, 258–266. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mean ± SEM | βBiW 2 | p 3 | |||
|---|---|---|---|---|---|
| BiW | Sex | ||||
| Response variables 4 for all piglets (n = 621 on day 44) | |||||
| BW gain (g/day) | Days 0–27 | 214 ± 2 | 102 | <0.001 | 0.153 |
| Days 27–44 5 | 398 ± 4 | 110 | <0.001 | 0.024 | |
| Days 0–44 6 | 285 ± 2 | 102 | <0.001 | 0.027 | |
| Response variables for focal piglets (n = 214 on day 44) | |||||
| BW gain (g/day) | Days 0–27 | 216 ± 3 | 85 | <0.001 | 0.270 |
| Days 27–44 | 412 ± 6 | 103 | <0.001 | 0.904 | |
| Days 0–44 | 291 ± 4 | 91 | <0.001 | 0.549 | |
| Nutrient dry matter (DM) intake (g/day) on days 42–43 | Sow feed | 163 ± 6 | 44 | 0.065 | 0.793 |
| Piglet feed | 293 ± 12 | 5 | 0.918 | 0.832 | |
| Total feed | 456 ± 13 | 50 | 0.354 | 0.925 | |
| Milk | 137 ± 7 | 102 | <0.001 | 0.420 | |
| Feeding behaviour during sow feeding times (%) on days 40–43 | Contacting sow feed | 11.9 ± 0.7 | 1.3 | 0.609 | 0.221 |
| Contacting piglet feed | 7.1 ± 0.5 | 2.7 | 0.205 | 0.832 | |
| Contacting total feed | 18.9 ± 0.8 | 4.0 | 0.235 | 0.410 | |
| Feeding behaviour during the day (%) on day 41 | Contacting sow feed 7 | 4.7 ± 0.3 | --- | 0.278 | 0.035 |
| Contacting piglet feed 8 | 4.6 ± 0.3 | --- | 0.481 | 0.045 | |
| Contacting total feed | 9.3 ± 0.5 | 0.8 | 0.672 | 0.968 | |
| Suckling behaviour: the presence at teats (no./7.5 h) on day 41 | Front and middle teats | 4.3 ± 0.2 | 2.3 | 0.009 | 0.883 |
| Rear teats | 1.7 ± 0.2 | 0.5 | 0.524 | 0.898 | |
| Total teats | 6.0 ± 0.2 | 2.8 | <0.001 | 0.746 | |
| Alien teats | 1.4 ± 0.2 | −0.3 | 0.705 | 0.467 | |
| Skin lesions (no.) | Day 27 | 5.2 ± 0.4 | 4.3 | 0.012 | 0.105 |
| Day 44 | 8.3 ± 0.5 | 3.3 | 0.108 | 0.746 | |
| Mean ± SEM | p3 | ||||||
|---|---|---|---|---|---|---|---|
| Piglet Type 2 | |||||||
| Variables 4 | HBiW-Fast | HBiW-Slow | LBiW-Fast | LBiW-Slow | BiW Class 2 | Growth Rate Class 2 | Interaction |
| No. of alive piglets on day 44 | |||||||
| 89 | 18 | 29 | 78 | --- | --- | --- | |
| BW (kg) | |||||||
| Day 0 5 | 1.6 ± 0.0 | 1.7 ± 0.0 | 1.3 ± 0.0 | 1.3 ± 0.0 | --- | --- | --- |
| Day 27 | 7.9 ± 0.1 | 7.1 ± 0.3 | 7.5 ± 0.2 | 6.5 ± 0.1 | 0.016 | <0.001 | 0.579 |
| Day 44 | 15.6 ± 0.2 | 12.9 ± 0.4 | 14.8 ± 0.4 | 12.8 ± 0.2 | 0.151 | <0.001 | 0.693 |
| BW gain (g/day) | |||||||
| Days 0–27 | 234 ± 4 | 200 ± 12 | 228 ± 8 | 194 ± 5 | 0.416 | <0.001 | 0.868 |
| Days 27–44 | 453 ± 9 | 353 ± 16 | 431 ± 17 | 371 ± 9 | 0.609 | <0.001 | 0.529 |
| Days 0–44 | 319 ± 5 | 255 ± 8 | 307 ± 9 | 262 ± 5 | 0.604 | <0.001 | 0.537 |
| Nutrient dry matter (DM) intake (g/day) on days 42–43 | |||||||
| Sow feed | 171 ± 9 | 159 ± 25 | 175 ± 16 | 150 ± 9 | 0.800 | 0.157 | 0.382 |
| Piglet feed | 326 ± 21 | 252 ± 38 | 275 ± 32 | 272 ± 19 | 0.461 | 0.198 | 0.497 |
| Total feed | 497 ± 21 | 411 ± 33 | 450 ± 34 | 423 ± 20 | 0.436 | 0.068 | 0.764 |
| Milk 6 | 165 ± 14 | 87 ± 14 | 145 ± 19 | 113 ± 10 | 0.277 | 0.536 | 0.484 |
| Feeding behaviour during sow feeding times (%) on days 40–43 | |||||||
| Contacting sow feed | 12.1 ± 1.0 | 13.0 ± 3.0 | 10.8 ± 1.9 | 11.7 ± 1.0 | 0.469 | 0.715 | 0.625 |
| Contacting piglet feed 7 | 7.5 ± 0.8 y | 6.3 ± 1.7 xy | 6.6 ± 1.1 x | 7.1 ± 0.8 xy | 0.584 | 0.901 | 0.071 |
| Contacting total feed | 19.6 ± 1.2 | 19.2 ± 3.8 | 17.4 ± 2.2 | 18.7 ± 1.3 | 0.472 | 0.744 | 0.378 |
| Feeding behaviour during the day (%) on day 41 | |||||||
| Contacting sow feed | 4.4 ± 0.5 | 4.3 ± 1.5 | 5.0 ± 1.1 | 5.0 ± 0.6 | 0.733 | 0.095 | 0.944 |
| Contacting piglet feed | 4.7 ± 0.5 | 4.0 ± 1.0 | 4.3 ± 0.7 | 4.8 ± 0.5 | 0.875 | 0.391 | 0.550 |
| Contacting total feed | 9.0 ± 0.7 | 8.2 ± 1.9 | 9.1 ± 1.4 | 9.9 ± 0.8 | 0.723 | 0.402 | 0.827 |
| Suckling behaviour: the presence at teats (no./7.5 h) on day 41 | |||||||
| Front and middle teats | 4.7 ± 0.3 | 4.1 ± 0.6 | 4.3 ± 0.6 | 4.0 ± 0.4 | 0.679 | 0.512 | 0.826 |
| Total teats | 6.1 ± 0.2 | 6.4 ± 0.5 | 6.3 ± 0.4 | 5.8 ± 0.3 | 0.665 | 0.859 | 0.614 |
| Alien teats | 1.2 ± 0.2 | 2.0 ± 0.7 | 0.8 ± 0.3 | 1.8 ± 0.3 | 0.383 | 0.010 | 0.833 |
| Skin lesions (no.) | |||||||
| Day 27 8 | 5.7 ± 0.7 b | 3.9 ± 1.2 a | 6.8 ± 1.3 ab | 4.5 ± 0.6 a | 0.667 | 0.039 | 0.084 |
| Day 44 | 9.0 ± 0.7 | 7.1 ± 1.4 | 9.2 ± 1.4 | 7.5 ± 0.9 | 0.424 | 0.047 | 0.518 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, T.; Gerrits, W.J.J.; van der Peet-Schwering, C.M.C.; Soede, N.M.; Reimert, I. Effects of Birthweight of Piglets in a Multi-Suckling System on Mortality, Growth Rate, Catch-Up Growth, Feed Intake and Behaviour. Animals 2023, 13, 297. https://doi.org/10.3390/ani13020297
Tang T, Gerrits WJJ, van der Peet-Schwering CMC, Soede NM, Reimert I. Effects of Birthweight of Piglets in a Multi-Suckling System on Mortality, Growth Rate, Catch-Up Growth, Feed Intake and Behaviour. Animals. 2023; 13(2):297. https://doi.org/10.3390/ani13020297
Chicago/Turabian StyleTang, Tianyue, Walter J. J. Gerrits, Carola M. C. van der Peet-Schwering, Nicoline M. Soede, and Inonge Reimert. 2023. "Effects of Birthweight of Piglets in a Multi-Suckling System on Mortality, Growth Rate, Catch-Up Growth, Feed Intake and Behaviour" Animals 13, no. 2: 297. https://doi.org/10.3390/ani13020297