
Effects of the Expressions and Variants of the CAST Gene on the Fatty Acid Composition of the Longissimus Thoracis Muscle of Grazing Sonid Sheep
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Determination of Fatty Acid Composition
2.3. RNA Extraction and Real-Time Quantitative Amplification
2.4. Resequencing and Variant Detection in CAST
2.5. Polymorphism Genotyping Using iPLEX MassARRAY
2.6. Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
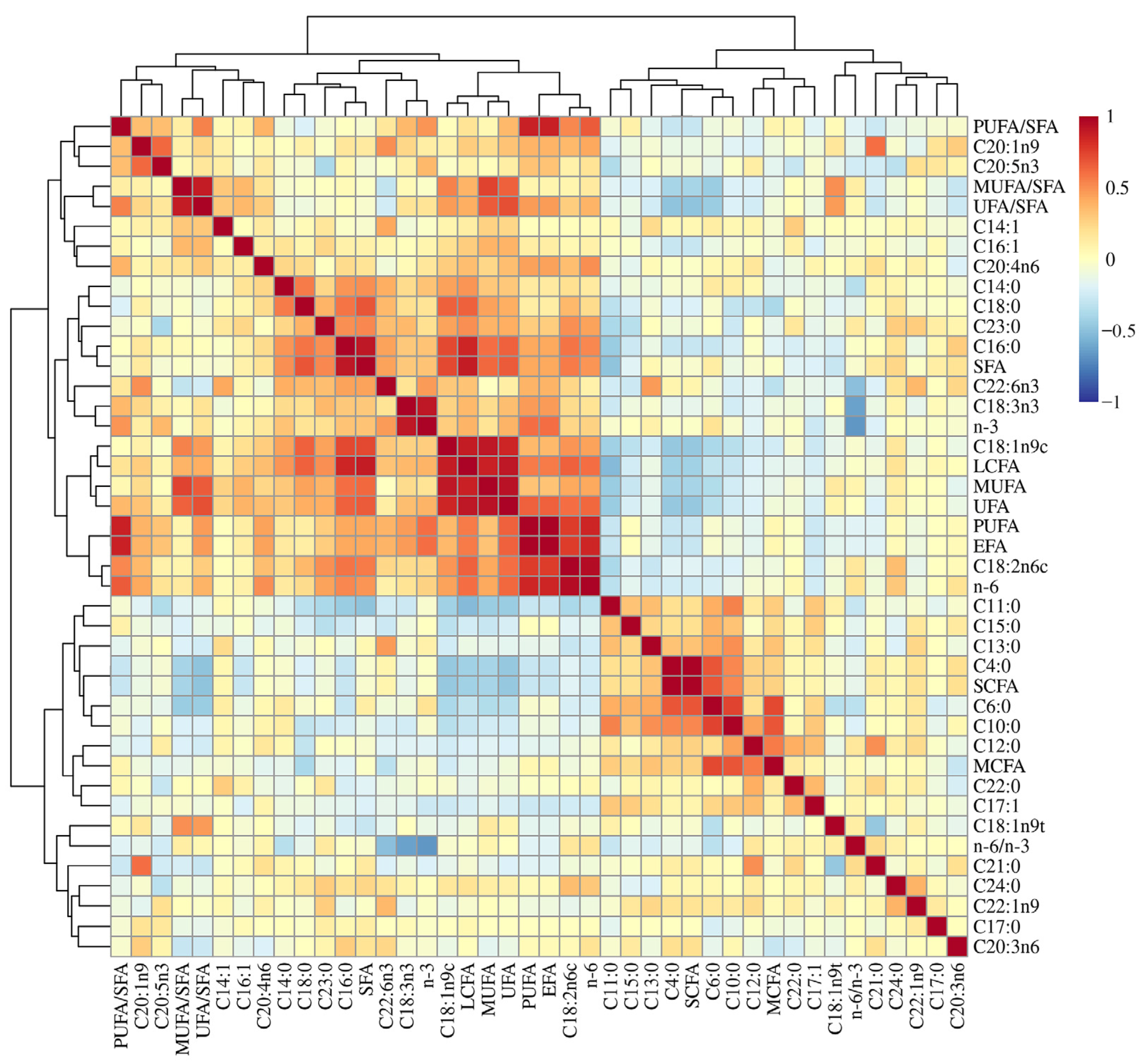
3.1. Fatty Acid Profiles of the Longissimus Thoracis Muscle
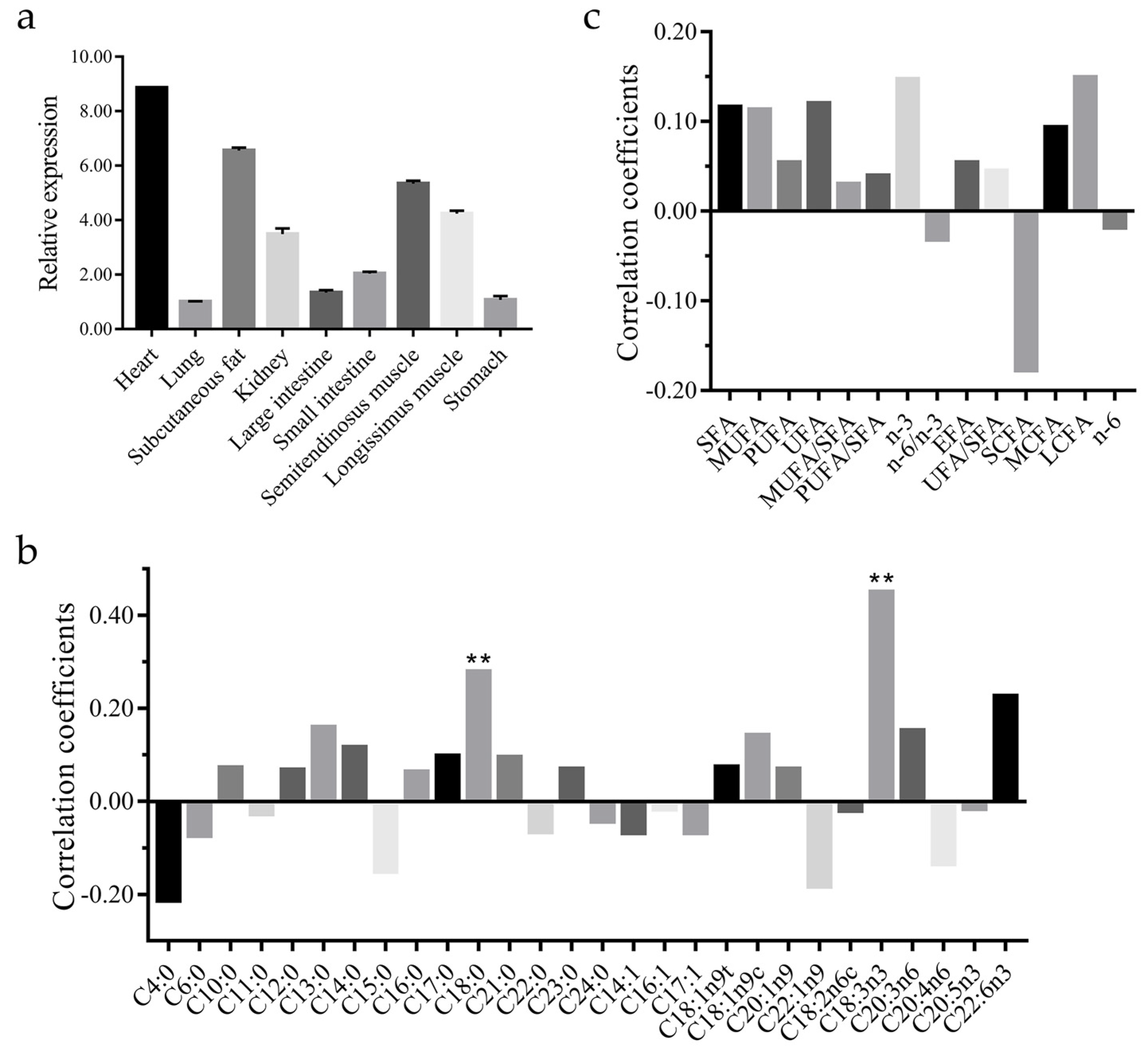
3.2. CAST Gene Expression Profiles in Sheep
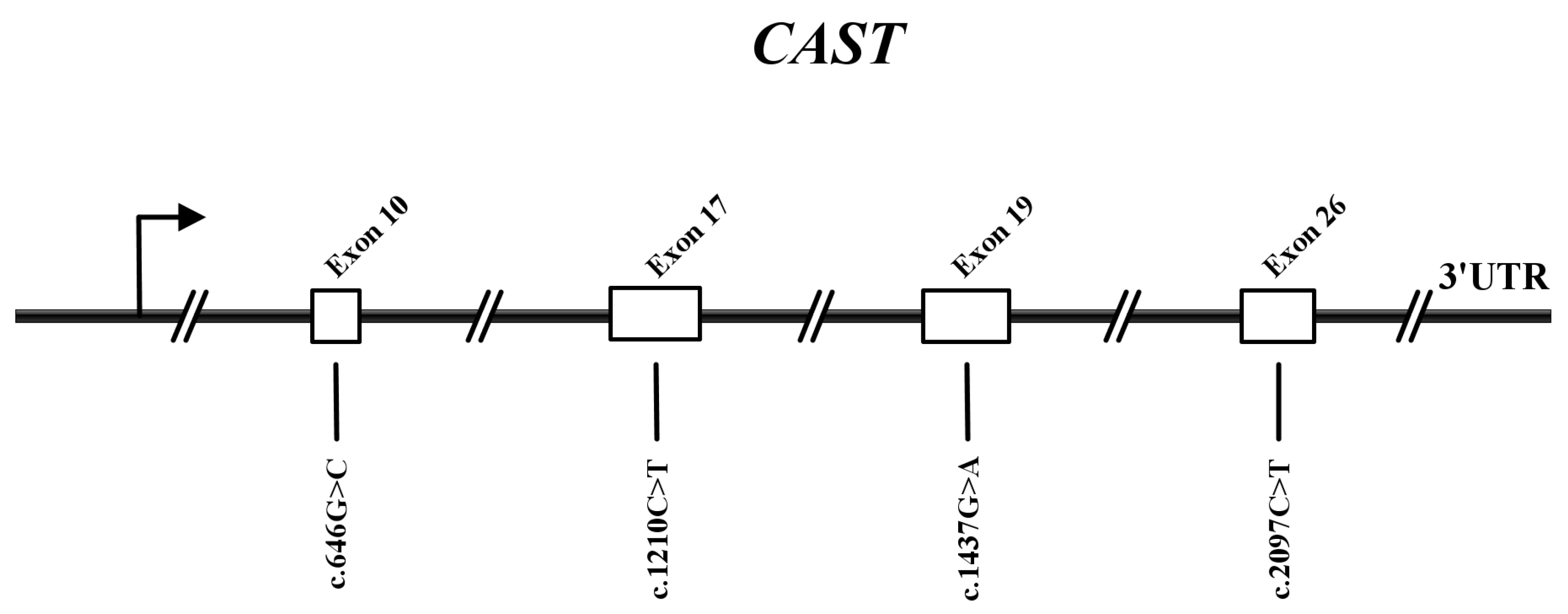
3.3. Variant Discovery in the CAST of Sonid Sheep
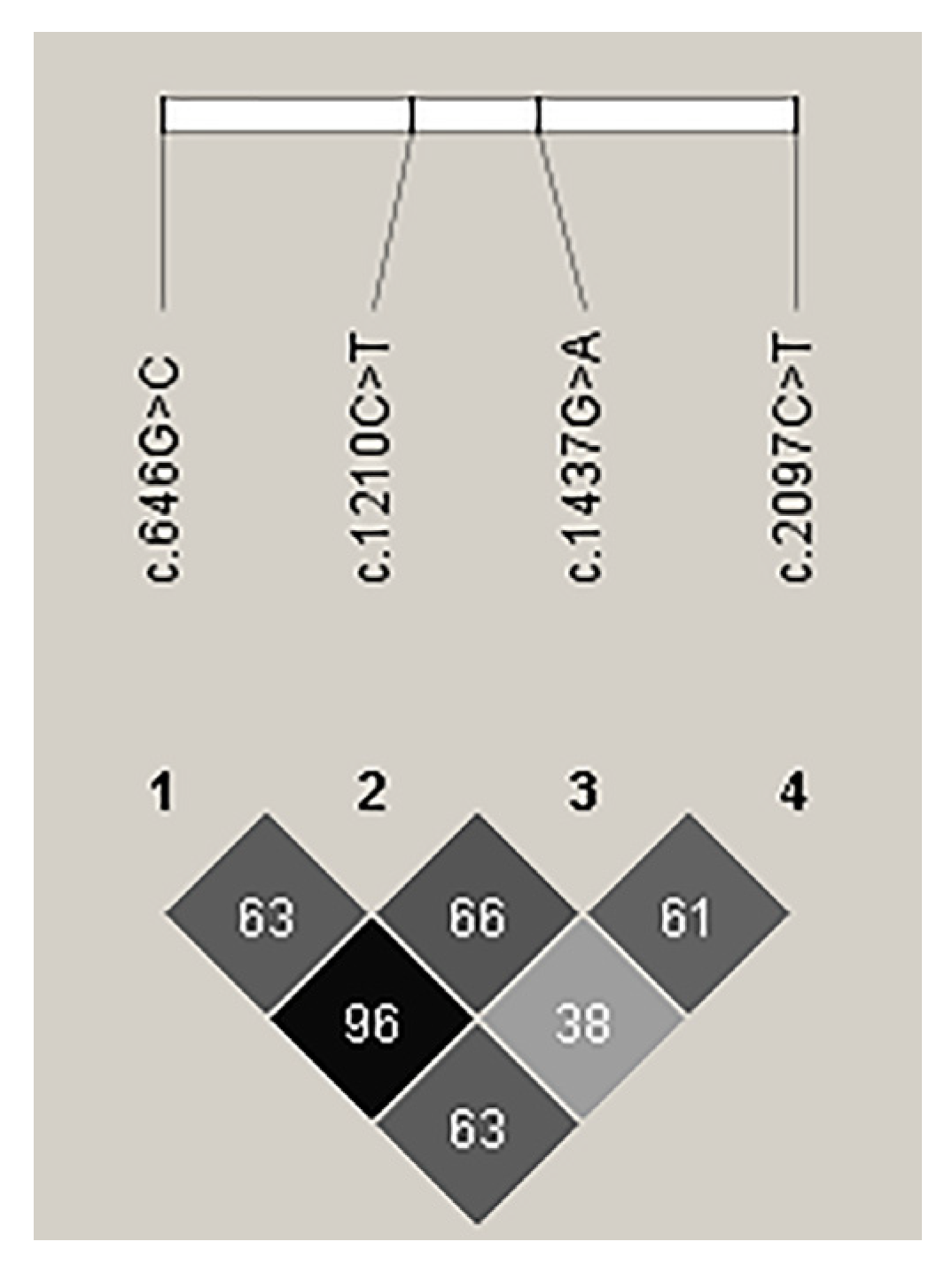
3.4. Linkage Disequilibrium Analysis of Novel Variants of CAST
3.5. Associations between Variants in CAST and Fatty Acid
3.6. Associations between Haplotypes of CAST and Fatty Acid
3.7. Bioinformatics analysis of ovine CAST
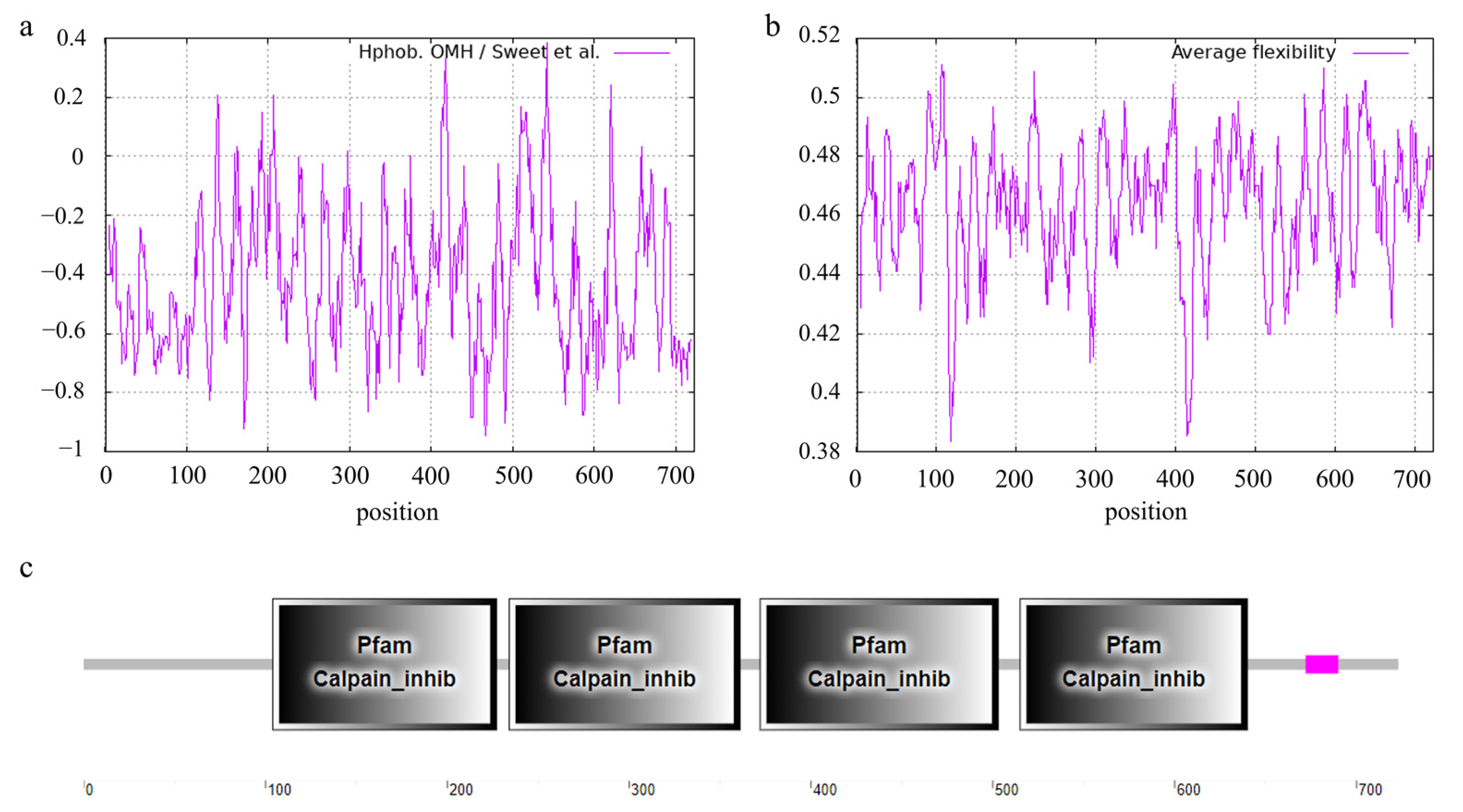
3.7.1. Feature and Structure Prediction of the Ovine CAST Protein
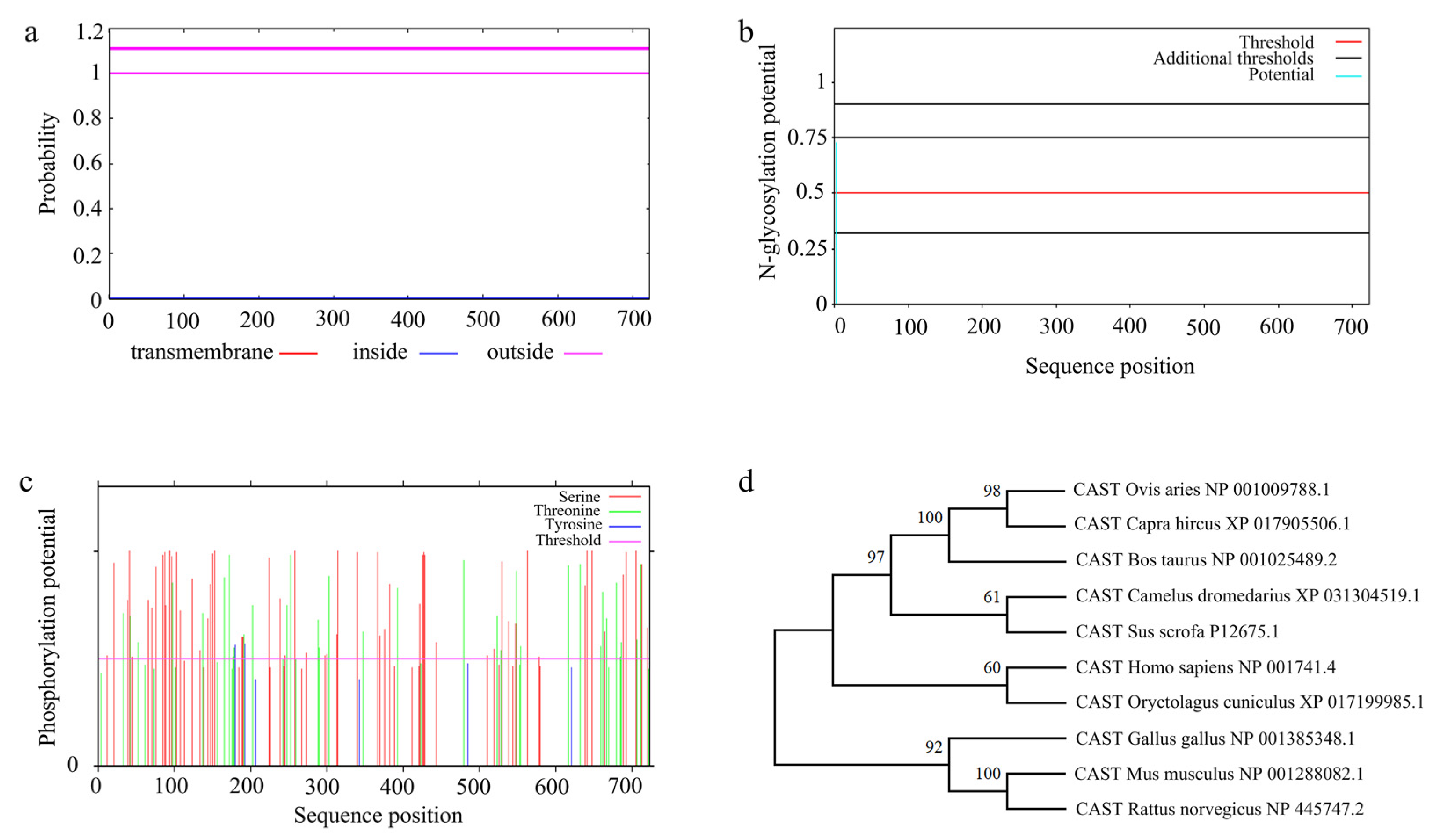
3.7.2. Amino Acid Sequence Analysis of Ovine CAST
3.7.3. Multiple Sequence Alignment and Phylogenetic
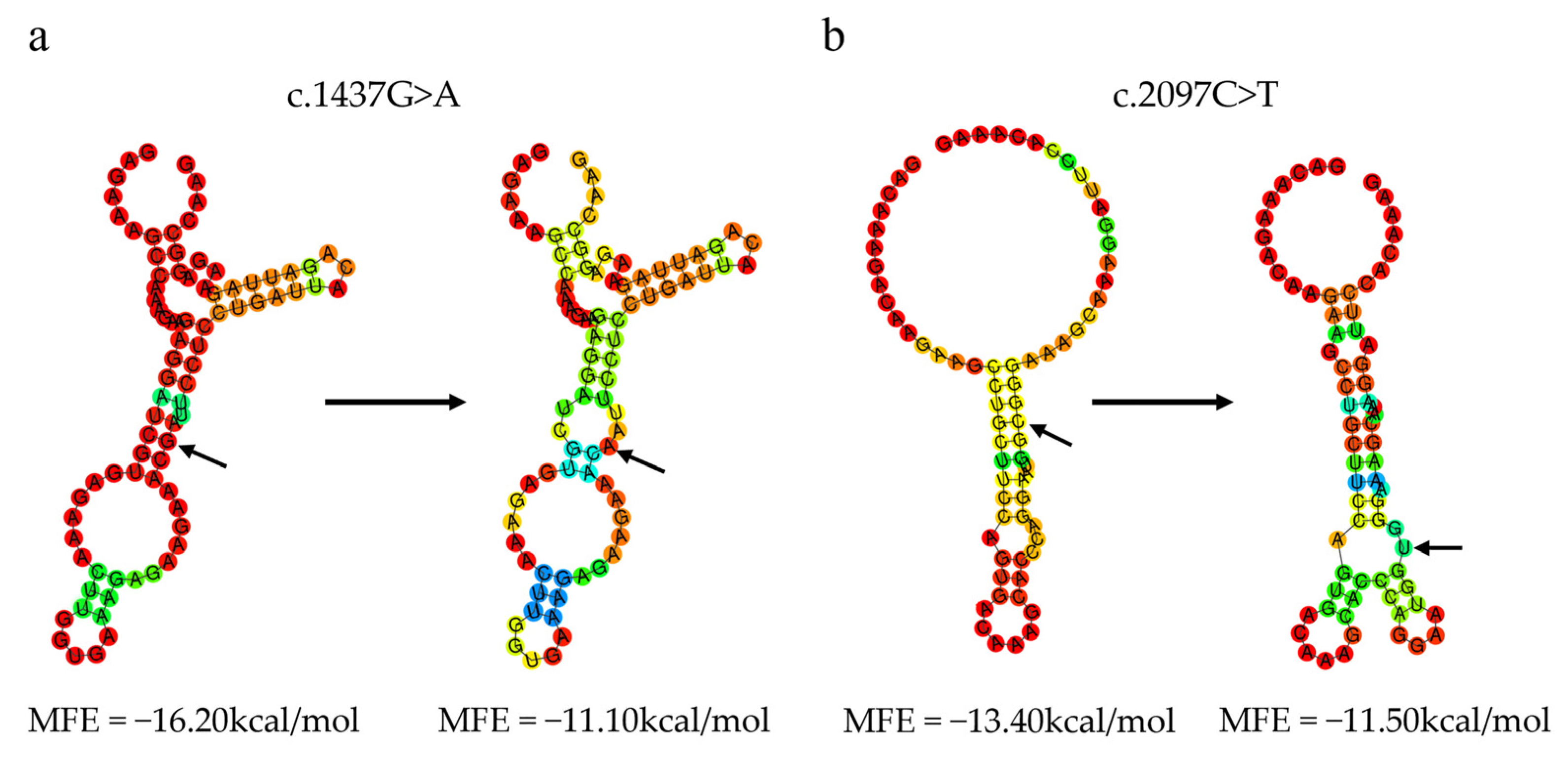
3.7.4. Effect of Variants on the mRNA Secondary Structure of the Ovine CAST Gene
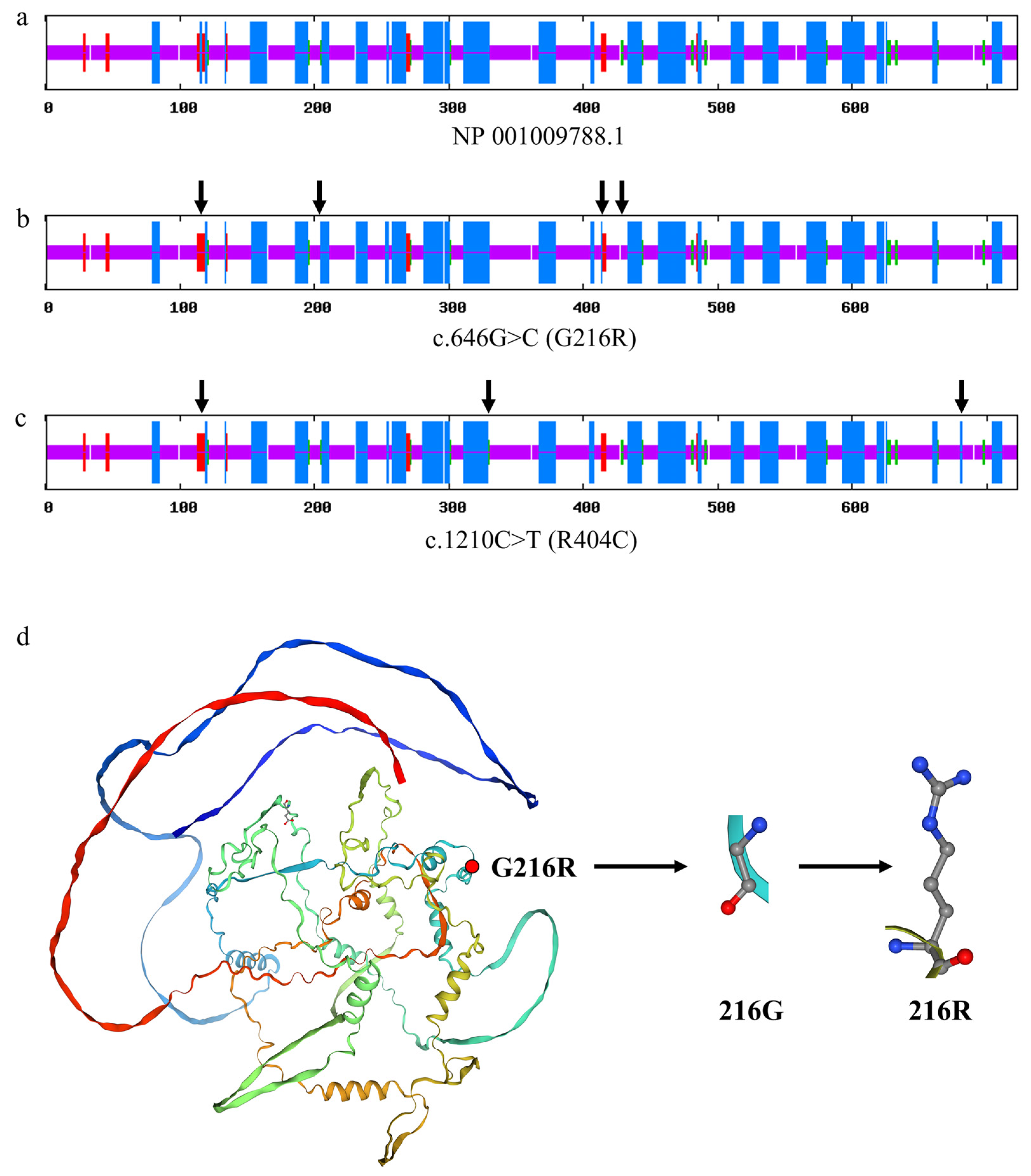
3.7.5. Effect of Variants on the Secondary and Tertiary Structure of the Ovine CAST Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- China National Commission of Animal Genetic Resources (CNCAGR). Animal Genetic Resources in China: Bovines; China Agriculture Press: Beijing, China, 2011; pp. 15–20. [Google Scholar]
- Hoy, A.J.; Nagarajan, S.R.; Butler, L.M. Tumour fatty acid metabolism in the context of therapy resistance and obesity. Nat. Rev. Cancer 2021, 21, 753–766. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Ladeira, M.M.; Schoonmaker, J.P.; Swanson, K.C.; Duckett, S.K.; Gionbelli, M.P.; Rodrigues, L.M.; Teixeira, P.D. Review: Nutrigenomics of marbling and fatty acid profile in ruminant meat. Animal 2018, 12, s282–s294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannier, L.; Gardner, G.E.; O’Reilly, R.A.; Pethick, D.W. Factors affecting lamb eating quality and the potential for their integration into an MSA sheepmeat grading model. Meat Sci. 2018, 144, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koohmaraie, M.; Geesink, G.H. Contribution of postmortem muscle biochemistry to the delivery of consistent meat quality with particular focus on the calpain system. Meat Sci. 2006, 74, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Koohmaraie, M.; Kent, M.P.; Shackelford, S.D.; Veiseth, E.; Wheeler, T.L. Meat tenderness and muscle growth: Is there any relationship? Meat Sci. 2002, 62, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, C.M.; Sensky, P.L.; Bardsley, R.G.; Buttery, P.J.; Parr, T. Tenderness--an enzymatic view. Meat Sci. 2010, 84, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Calvo, J.H.; Iguacel, L.P.; Kirinus, J.K.; Serrano, M.; Ripoll, G.; Casasus, I.; Joy, M.; Perez-Velasco, L.; Sarto, P.; Alberti, P.; et al. A new single nucleotide polymorphism in the calpastatin (CAST) gene associated with beef tenderness. Meat Sci. 2014, 96, 775–782. [Google Scholar] [CrossRef]
- Ropka-Molik, K.; Bereta, A.; Tyra, M.; Rozycki, M.; Piorkowska, K.; Szyndler-Nedza, M.; Szmatola, T. Association of calpastatin gene polymorphisms and meat quality traits in pig. Meat Sci. 2014, 97, 143–150. [Google Scholar] [CrossRef]
- Davoli, R.; Schivazappa, C.; Zambonelli, P.; Braglia, S.; Rossi, A.; Virgili, R. Association study between single nucleotide polymorphisms in porcine genes and pork quality traits for fresh consumption and processing into Italian dry-cured ham. Meat Sci. 2017, 126, 73–81. [Google Scholar] [CrossRef]
- Castro, S.; Rios, M.; Ortiz, Y.; Manrique, C.; Jimenez, A.; Ariza, F. Association of single nucleotide polymorphisms in CAPN1, CAST and MB genes with meat color of Brahman and crossbreed cattle. Meat Sci. 2016, 117, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jin, S.; Kim, H.J.; Bhuiyan, M.S.A.; Lee, D.H.; Lee, S.H.; Jang, S.B.; Han, M.H.; Lee, S.H. Validation Study of SNPs in CAPN1-CAST Genes on the Tenderness of Muscles (Longissimus thoracis and Semimembranosus) in Hanwoo (Korean Cattle). Animals 2019, 9, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadhil, M.; Zulkadir, U. Association between polymorphisms of Myf5, MSTN and CAST genes and fattening performance in Brown Swiss and Holstein cattle breeds. Anim. Biotechnol. 2021, 32, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gregula-Kania, M.; Gruszecki, T.M.; Junkuszew, A.; Juszczuk-Kubiak, E.; Florek, M. Association of CAST gene polymorphism with carcass value and meat quality in two synthetic lines of sheep. Meat Sci. 2019, 154, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Tong, B.; Gao, G.Q.; Muramatsu, Y.; Ohta, T.; Kose, H.; Li, G.P.; Fatchiyah, F.; Yamada, T. Association of the expression levels in the longissimus muscle and a SNP in the CDC10 gene with marbling in Japanese Black beef cattle. Meat Sci. 2015, 108, 28–31. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Gabriel, S.; Ziaugra, L.; Tabbaa, D. SNP genotyping using the Sequenom MassARRAY iPLEX platform. Curr. Protoc. Hum. Genet. 2009, 60, 2.12.1–2.12.18. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Nei, M.; Roychoudhury, A.K. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.Y.; Cheng, G.; Cheng, Z.X.; Bao, C.; Yamada, T.; Cao, G.F.; Bao, S.Q.; Schreurs, N.M.; Zan, L.S.; Tong, B. Association of variants in FABP4, FASN, SCD, SREBP1 and TCAP genes with intramuscular fat, carcass traits and body size in Chinese Qinchuan cattle. Meat Sci. 2022, 192, 108882. [Google Scholar] [CrossRef]
- Mathews, D.H.; Disney, M.D.; Childs, J.L.; Schroeder, S.J.; Zuker, M.; Turner, D.H. Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Proc. Natl. Acad. Sci. USA 2004, 101, 7287–7292. [Google Scholar] [CrossRef] [Green Version]
- Biesalski, H.K. Meat as a component of a healthy diet—Are there any risks or benefits if meat is avoided in the diet? Meat Sci. 2005, 70, 509–524. [Google Scholar] [CrossRef]
- McNeill, S.H. Inclusion of red meat in healthful dietary patterns. Meat Sci. 2014, 98, 452–460. [Google Scholar] [CrossRef]
- Micha, R.; Michas, G.; Lajous, M.; Mozaffarian, D. Processing of meats and cardiovascular risk: Time to focus on preservatives. BMC Med. 2013, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Stables, M.J.; Gilroy, D.W. Old and new generation lipid mediators in acute inflammation and resolution. Prog. Lipid Res. 2011, 50, 35–51. [Google Scholar] [CrossRef]
- Wood, J.D. Fat Deposition and the Quality of Fat Tissue in Meat Animals. Fats in Animal Nutrition; Wiseman, J., Ed.; Butterworths: London, UK, 1984; pp. 407–435. [Google Scholar]
- Jacob, R.H.; Pethick, D.W. Animal factors affecting the meat quality of Australian lamb meat. Meat Sci. 2014, 96, 1120–1123. [Google Scholar] [CrossRef]
- Gunawan, A.; Listyarini, K.; Harahap, R.S.; Jakaria; Roosita, K.; Sumantri, C.; Inounu, I.; Akter, S.H.; Islam, M.A.; Uddin, M.J. Hepatic transcriptome analysis identifies genes, polymorphisms and pathways involved in the fatty acids metabolism in sheep. PLoS ONE 2021, 16, e0260514. [Google Scholar] [CrossRef]
- Duan, J.; Wainwright, M.S.; Comeron, J.M.; Saitou, N.; Sanders, A.R.; Gelernter, J.; Gejman, P.V. Synonymous mutations in the human dopamine receptor D2 (DRD2) affect mRNA stability and synthesis of the receptor. Hum. Mol. Genet. 2003, 12, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Kimchi-Sarfaty, C.; Oh, J.M.; Kim, I.-W.; Sauna, Z.E.; Calcagno, A.M.; Ambudkar, S.V.; Gottesman, M.M. A "Silent" Polymorphism in the MDR1 Gene Changes Substrate Specificity. Science 2007, 315, 525–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagani, F.; Raponi, M.; Baralle, F.E. Synonymous mutations in CFTR exon 12 affect splicing and are not neutral in evolution. Proc. Natl. Acad. Sci. USA 2005, 102, 6368–6372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, A.M. α-Adducin nsSNPs affect mRNA secondary structure, protein modification and stability. Meta Gene 2018, 17, 153–162. [Google Scholar] [CrossRef]
- Soemedi, R.; Cygan, K.J.; Rhine, C.L.; Glidden, D.T.; Taggart, A.J.; Lin, C.-L.; Fredericks, A.M.; Fairbrother, W.G. The effects of structure on pre-mRNA processing and stability. Methods 2017, 125, 36–44. [Google Scholar] [CrossRef]
- Sauna, Z.E.; Kimchi-Sarfaty, C. Understanding the contribution of synonymous mutations to human disease. Nat. Rev. Genet. 2011, 12, 683–691. [Google Scholar] [CrossRef]
- Fung, K.L.; Pan, J.; Ohnuma, S.; Lund, P.E.; Pixley, J.N.; Kimchi-Sarfaty, C.; Ambudkar, S.V.; Gottesman, M.M. MDR1 synonymous polymorphisms alter transporter specificity and protein stability in a stable epithelial monolayer. Cancer Res. 2014, 74, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Bevilacqua, P.C.; Ritchey, L.E.; Su, Z.; Assmann, S.M. Genome-Wide Analysis of RNA Secondary Structure. Annu. Rev. Genet. 2016, 50, 235–266. [Google Scholar] [CrossRef] [Green Version]
- Rose, J.; Kraft, T.; Brenner, B.; Montag, J. Hypertrophic cardiomyopathy MYH7 mutation R723G alters mRNA secondary structure. Physiol. Genom. 2020, 52, 15–19. [Google Scholar] [CrossRef]
- Warner, R.D.; Greenwood, P.L.; Pethick, D.W.; Ferguson, D.M. Genetic and environmental effects on meat quality. Meat Sci. 2010, 86, 171–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff Lonergan, E.; Zhang, W.; Lonergan, S.M. Biochemistry of postmortem muscle—Lessons on mechanisms of meat tenderization. Meat Sci. 2010, 86, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Emori, Y.; Kawasaki, H.; Imajoh, S.; Minami, Y.; Suzuki, K. All four repeating domains of the endogenous inhibitor for calcium-dependent protease independently retain inhibitory activity. Expression of the cDNA fragments in Escherichia coli. J. Biol. Chem. 1988, 263, 2364–2370. [Google Scholar] [CrossRef] [PubMed]
- Fiego, D.P.L.; Santoro, P.; Macchioni, P.; De Leonibus, E. Influence of genetic type, live weight at slaughter and carcass fatness on fatty acid composition of subcutaneous adipose tissue of raw ham in the heavy pig. Meat Sci. 2005, 69, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Pena, R.N.; Ros-Freixedes, R.; Tor, M.; Estany, J. Genetic Marker Discovery in Complex Traits: A Field Example on Fat Content and Composition in Pigs. Int. J. Mol. Sci. 2016, 17, 2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Shoji, N.; Honda, T.; Oyama, K. Genetic relationships between meat quality traits and fatty acid composition in Japanese black cattle. Anim. Sci. J. 2017, 88, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Sellier, P.; Maignel, L.; Bidanel, J.P. Genetic parameters for tissue and fatty acid composition of backfat, perirenal fat and longissimus muscle in Large White and Landrace pigs. Animal 2010, 4, 497–504. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Product Length (bp) | Annealing Temperature (°C) | Accession Number |
|---|---|---|---|---|
| GAPDH | F: AATACTGAGATGTCCTTC | 140 | 53.8 | NM_001190390.1 |
| R: TTTATGGTGGTTGATTTC | ||||
| CAST | F: ATCCAGAAGACGGAAAGCCT | 144 | 61.3 | NM_001009788.1 |
| R: GCAGTGGTTTTCCGTCTTTATCCTT |
| Polymorphism | Primers (5′-3′) | Tm (°C) |
|---|---|---|
| c.646G>C (G216R) | F: ACGTTGGATGCTTGCCTTCTCAGCATCATC | 52.2 |
| R: ACGTTGGATGTAAACTCACCGAGGAGTCTG | ||
| E: GAGGCCCTGCGATCC | ||
| c.1210C>T (R404C) | F: ACGTTGGATGGTTGTGTCTGCTGCTTTCTG | 70.4 |
| R: ACGTTGGATGTGACTGCACAGAACACATGG | ||
| E: CCGGCATCCCCGGCCGCTGCCCC | ||
| c.1437G>A (479T) | F: ACGTTGGATGAGCCAAAGAAGAGGATCGTG | 45.3 |
| R: ACGTTGGATGCCTTGGCCTCTTCTAATCTG | ||
| E: ATCTGTAATCAGGAGGAAT | ||
| c.2097C>T (699G) | F: ACGTTGGATGAAGACAAGAAGCCTGCTTCC | 52.0 |
| R: ACGTTGGATGGACACAACCGAGCTTTGAAC | ||
| E: GTGGAATCCTTTGCTTTCCC |
| Fatty Acid Composition | c.1210C>T (R404C) | c.1437G>A in LD-M | c.2097C>T | ||||
|---|---|---|---|---|---|---|---|
| Genotype | Genotype | Genotype | |||||
| CC (253) | CT (114) | TT (11) | GG (288) | GA (84) | CC (287) | CT (89) | |
| C10:0 | 0.31 ± 0.01 | 0.34 ± 0.02 | 0.35 ± 0.05 | 0.31 ± 0.01 a | 0.36 ± 0.02 b | 0.31 ± 0.01 a | 0.37 ± 0.02 b |
| C12:0 | 0.44 ± 0.02 a | 0.59 ± 0.07 b | 0.52 ± 0.16 ab | 0.47 ± 0.03 | 0.56 ± 0.08 | 0.47 ± 0.03 | 0.57 ± 0.07 |
| C14:0 | 1.56 ± 0.04 A | 1.36 ± 0.05 B | 1.32 ± 0.15 AB | 1.56 ± 0.04 A | 1.30 ± 0.06 B | 1.51 ± 0.04 | 1.44 ± 0.06 |
| C18:0 | 9.24 ± 0.13 a | 8.64 ± 0.18 b | 8.95 ± 0.58 ab | 9.24 ± 0.12 A | 8.50 ± 0.21 B | 9.12 ± 0.12 | 8.84 ± 0.22 |
| SFA | 36.12 ± 0.35 | 34.79 ± 0.43 | 36.84 ± 1.41 | 36.22 ± 0.32 a | 34.45 ± 0.50 b | 35.91 ± 0.30 | 35.04 ± 0.59 |
| C18:1n9t | 1.71 ± 0.14 | 2.17 ± 0.22 | 1.80 ± 0.06 | 1.71 ± 0.12 a | 2.30 ± 0.27 b | 1.76 ± 0.12 | 2.28 ± 0.34 |
| C18:1n9c | 16.19 ± 0.28 a | 14.96 ± 0.40 b | 16.09 ± 0.94 ab | 16.14 ± 0.27 a | 14.77 ± 0.42 b | 15.99 ± 0.26 | 15.29 ± 0.46 |
| C18:3n3 | 1.76 ± 0.04 a | 1.55 ± 0.08 b | 1.51 ± 0.24 ab | 1.74 ± 0.04 | 1.53 ± 0.09 | 1.75 ± 0.04 a | 1.53 ± 0.08 b |
| n-6 | 5.45 ± 0.09 A | 5.47 ± 0.14 AB | 6.44 ± 0.40 B | 5.43 ± 0.08 | 5.61 ± 0.18 | 5.43 ± 0.08 | 5.69 ± 0.18 |
| n-3 | 2.76 ± 0.06 A | 2.51 ± 0.09 B | 2.36 ± 0.30 B | 2.73 ± 0.06 A | 2.50 ± 0.10 B | 2.76 ± 0.06 a | 2.42 ± 0.09 b |
| n-6/n-3 | 1.97 ± 0.23 A | 2.18 ± 0.59 B | 2.73 ± 1.22 B | 1.99 ± 0.24 A | 2.24 ± 0.71 B | 1.97 ± 0.28 A | 2.35 ± 0.56 B |
| Haplotype | c.646G>C | c.1210C>T | c.1437G>A | c.2097C>T | Frequency |
|---|---|---|---|---|---|
| H1 (604) | G | C | G | C | 0.798 |
| H2 (77) | C | T | A | T | 0.101 |
| H3 (40) | G | T | G | C | 0.053 |
| Fatty Acid Composition | Genotype of Combination (Number) | ||
|---|---|---|---|
| H1H1 (238) | H1H2 (64) | H1H3 (35) | |
| (GGCCGGCC) | (GCCTGACT) | (GGCTGGCC) | |
| C12:0 | 0.44 ± 0.02 a | 0.59 ± 0.09 ab | 0.69 ± 0.15 b |
| C21:0 | 0.54 ± 0.03 A | 0.52 ± 0.06 a | 1.2 ± 0.52 Bb |
| C18:3n3 | 1.77 ± 0.05 a | 1.52 ± 0.11 b | 1.59 ± 0.14 ab |
| n-3 | 2.78 ± 0.06 A | 2.44 ± 0.11 B | 2.51 ± 0.17 AB |
| n-6/n-3 | 1.96 ± 0.24 a | 2.28 ± 0.73 b | 2.12 ± 0.96 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Li, T.; Lu, D.; Yamada, T.; Li, X.; Bao, S.; Liu, J.; Borjigin, G.; Cang, M.; Tong, B. Effects of the Expressions and Variants of the CAST Gene on the Fatty Acid Composition of the Longissimus Thoracis Muscle of Grazing Sonid Sheep. Animals 2023, 13, 195. https://doi.org/10.3390/ani13020195
Guo X, Li T, Lu D, Yamada T, Li X, Bao S, Liu J, Borjigin G, Cang M, Tong B. Effects of the Expressions and Variants of the CAST Gene on the Fatty Acid Composition of the Longissimus Thoracis Muscle of Grazing Sonid Sheep. Animals. 2023; 13(2):195. https://doi.org/10.3390/ani13020195
Chicago/Turabian StyleGuo, Xin, Terigele Li, Datong Lu, Takahisa Yamada, Xihe Li, Siqin Bao, Jiasen Liu, Gerelt Borjigin, Ming Cang, and Bin Tong. 2023. "Effects of the Expressions and Variants of the CAST Gene on the Fatty Acid Composition of the Longissimus Thoracis Muscle of Grazing Sonid Sheep" Animals 13, no. 2: 195. https://doi.org/10.3390/ani13020195