
SpPdp11 Administration in Diet Modified the Transcriptomic Response and Its Microbiota Associated in Mechanically Induced Wound Sparus aurata Skin
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Bacterial Growth
2.2. Animals, Experimental Design, and Sampling
2.3. DNA Extraction and Sequencing of the 16S rRNA Gene from the Skin-Associated Microbiota
2.4. RNA Isolation, Quality Control and Sequencing
2.5. Differential Expression Analysis and Functional Enrichment
2.6. Microbiota and Genes Correlation
2.7. Ethics Approval and Consent to Participate
3. Results
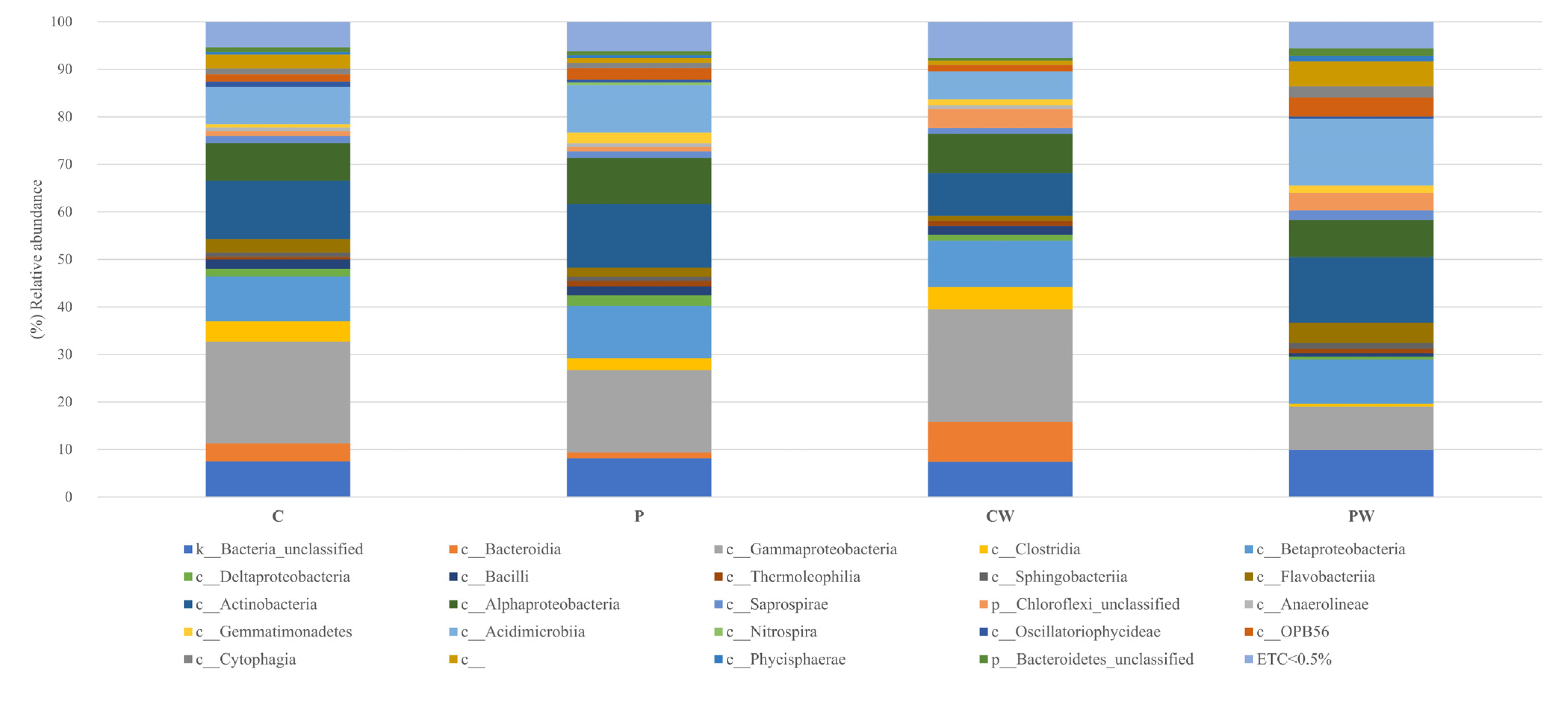
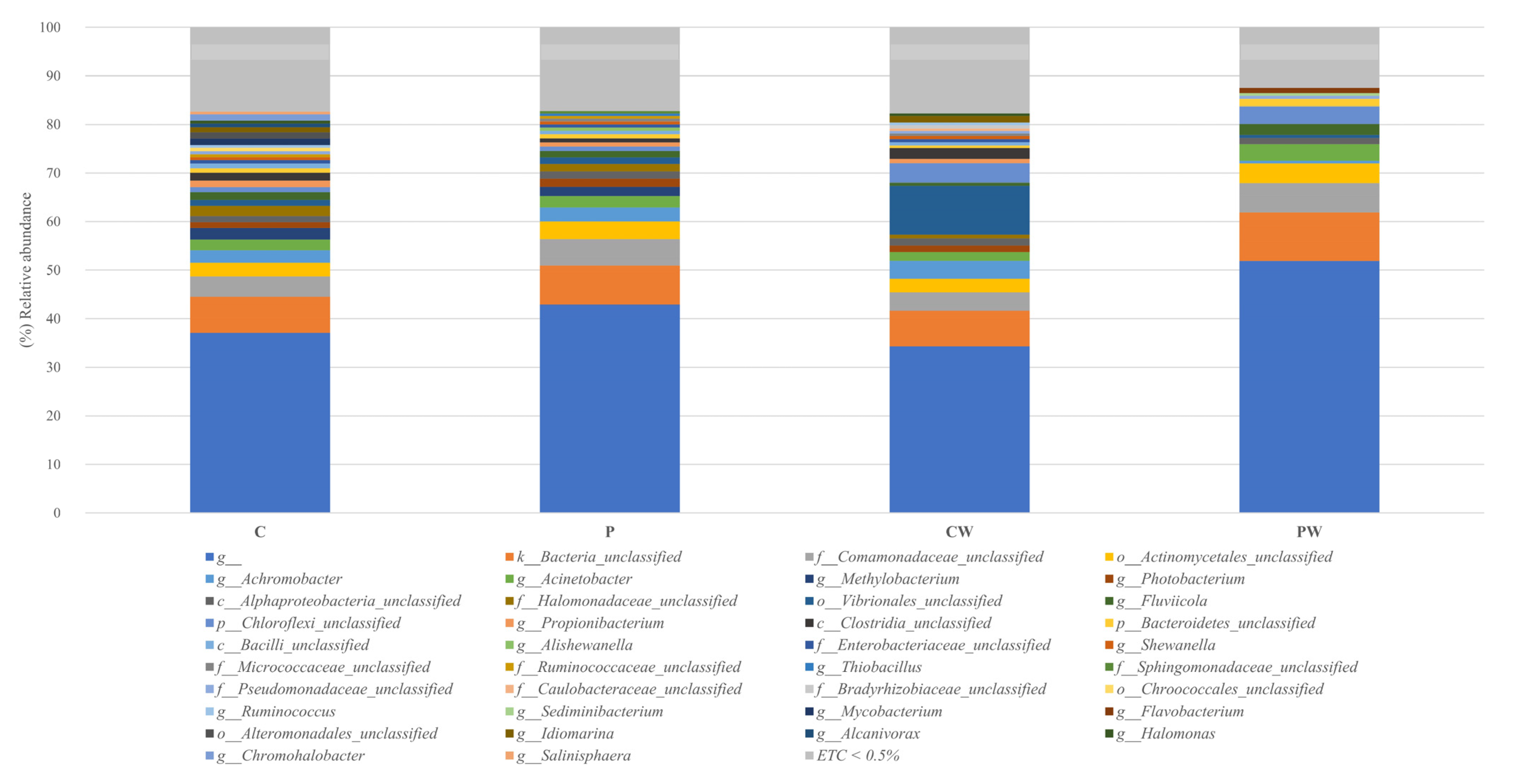
3.1. Skin Microbiota Analysis
3.2. Skin Transcriptome Sequencing and Analysis of Differentially Expressed Genes (DEGs)
3.3. Analysis of DEGs with the Highest Expression Change Value
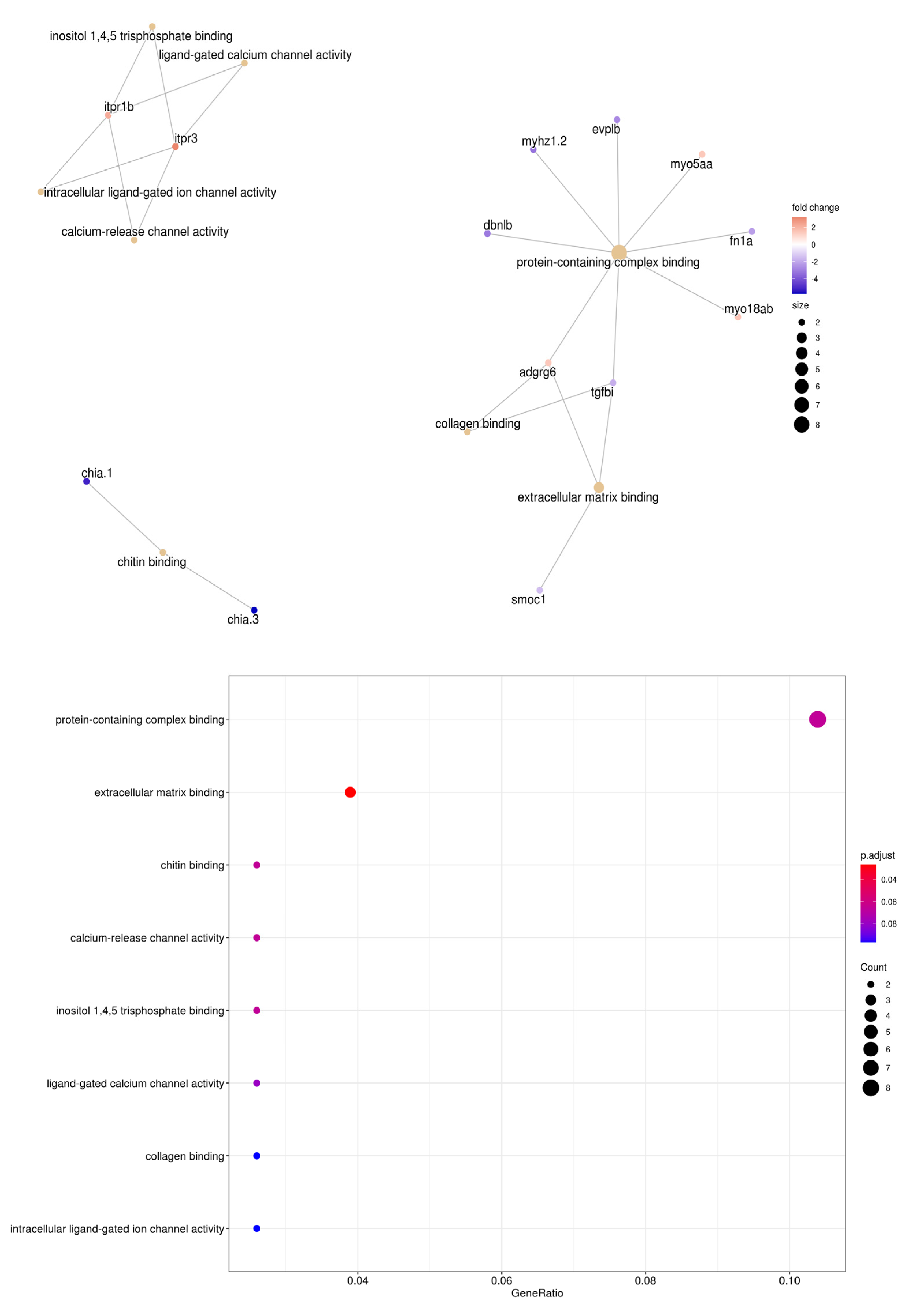
3.4. Enrichment Analysis of S. aurata Transcriptome and Its Differentially Expressed Genes
3.5. Correlation between the Skin Mucus-Associated Microbiota and the Most Up- or Down-Regulated Genes in Wounded S. aurata
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karlsen, C.; Ottem, K.F.; Brevik, Ø.J.; Davey, M.; Sørum, H.; Winther-Larsen, H.C. The Environmental and Host-Associated Bacterial Microbiota of Arctic Seawater-Farmed Atlantic Salmon with Ulcerative Disorders. J. Fish Dis. 2017, 40, 1645–1663. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, F.A.; Mabrok, M.; Machado, M.; Azeredo, R.; Afonso, A.; Esteban, M.A.; Costas, B. Mucosal and Systemic Immune Responses in Senegalese Sole (Solea senegalensis Kaup) Bath Challenged with Tenacibaculum maritimum: A Time-Course Study. Fish Shellfish Immunol. 2019, 87, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Angeles Esteban, M.; Editors, A.; Bensussan, A.; Flaño, E.; Hayball, J.D.; Puccetti, P. An Overview of the Immunological Defenses in Fish Skin. Int. Sch. Res. Netw. ISRN Immunol. 2012, 2012, 853470. [Google Scholar] [CrossRef] [Green Version]
- Fast, M.D.; Sims, D.E.; Burka, J.F.; Mustafa, A.; Ross, N.W. Skin Morphology and Humoral Non-Specific Defence Parameters of Mucus and Plasma in Rainbow Trout, Coho and Atlantic Salmon. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.E. Immunity to Bacteria in Fish. Fish Shellfish Immunol. 1999, 9, 291–308. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The Skin Microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Vendrell, D.; de Blas, I.; Ruiz-Zarzuela, I.; Gironés, O.; Múzquiz, J.L. In Vitro Competitive Adhesion and Production of Antagonistic Compounds by Lactic Acid Bacteria against Fish Pathogens. Vet. Microbiol. 2007, 122, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Peatman, E.; Lange, M.; Zhao, H.; Beck, B.H. Physiology and Immunology of Mucosal Barriers in Catfish (Ictalurus spp.). Tissue Barriers 2015, 3, e1068907. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.; Salinas, I. Under Pressure: Interactions between Commensal Microbiota and the Teleost Immune System. Front. Immunol. 2017, 8, 559. [Google Scholar] [CrossRef] [Green Version]
- Hoseinifar, S.H.; Shakouri, M.; Yousefi, S.; Van Doan, H.; Shafiei, S.; Yousefi, M.; Mazandarani, M.; Torfi Mozanzadeh, M.; Tulino, M.G.; Faggio, C. Humoral and Skin Mucosal Immune Parameters, Intestinal Immune Related Genes Expression and Antioxidant Defense in Rainbow Trout (Oncorhynchus mykiss) Fed Olive (Olea europea L.) Waste. Fish Shellfish Immunol. 2020, 100, 171–178. [Google Scholar] [CrossRef]
- Sylvain, F.É.; Derome, N. Vertically and Horizontally Transmitted Microbial Symbionts Shape the Gut Microbiota Ontogenesis of a Skin-Mucus Feeding Discus Fish Progeny. Sci. Rep. 2017, 7, 5263. [Google Scholar] [CrossRef] [Green Version]
- Gajardo, K.; Jaramillo-Torres, A.; Kortner, T.M.; Merrifield, D.L.; Tinsley, J.; Bakke, A.M.; Krogdahl, Å. Alternative Protein Sources in the Diet Modulate Microbiota and Functionality in the Distal Intestine of Atlantic Salmon (Salmo salar). Appl. Environ. Microbiol. 2017, 83, e02615-16. [Google Scholar] [CrossRef] [Green Version]
- Landeira-Dabarca, A.; Sieiro, C.; Álvarez, M. Change in Food Ingestion Induces Rapid Shifts in the Diversity of Microbiota Associated with Cutaneous Mucus of Atlantic Salmon Salmo salar. J. Fish Biol. 2013, 82, 893–906. [Google Scholar] [CrossRef]
- Wu, H.J.; Wu, E. The Role of Gut Microbiota in Immune Homeostasis and Autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Legrand, T.P.R.A.; Catalano, S.R.; Wos-Oxley, M.L.; Stephens, F.; Landos, M.; Bansemer, M.S.; Stone, D.A.J.; Qin, J.G.; Oxley, A.P.A. The Inner Workings of the Outer Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail Kingfish. Front. Microbiol. 2018, 8, 2664. [Google Scholar] [CrossRef]
- Mohammed, H.H.; Arias, C.R. Potassium Permanganate Elicits a Shift of the External Fish Microbiome and Increases Host Susceptibility to Columnaris Disease. Vet. Res. 2015, 46, 82. [Google Scholar] [CrossRef] [Green Version]
- Lokesh, J.; Kiron, V. Transition from Freshwater to Seawater Reshapes the Skin-Associated Microbiota of Atlantic Salmon. Sci. Rep. 2016, 6, 19707. [Google Scholar] [CrossRef] [Green Version]
- Mateus, A.P.; Mourad, M.; Power, D.M. Skin Damage Caused by Scale Loss Modifies the Intestine of Chronically Stressed Gilthead Sea Bream (Sparus aurata, L.). Dev. Comp. Immunol. 2021, 118, 103989. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Rodiles, A. The Fish Microbiome and Its Interactions with Mucosal Tissues. Mucosal Health Aquac. 2015, 10, 273–295. [Google Scholar] [CrossRef]
- Reverter, M.; Tapissier-Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Biological and Ecological Roles of External Fish Mucus: A Review. Fishes 2018, 3, 41. [Google Scholar] [CrossRef]
- Cámara-Ruiz, M.; Cerezo, I.M.; Guardiola, F.A.; García-Beltrán, J.M.; Balebona, M.C.; Moriñigo, M.Á.; Esteban, M.Á. Alteration of the Immune Response and the Microbiota of the Skin during a Natural Infection by Vibrio harveyi in European Seabass (Dicentrarchus labrax). Microorganisms 2021, 9, 964. [Google Scholar] [CrossRef] [PubMed]
- Segner, H.; Sundh, H.; Buchmann, K.; Douxfils, J.; Sundell, K.S.; Mathieu, C.; Ruane, N.; Jutfelt, F.; Toften, H.; Vaughan, L. Health of Farmed Fish: Its Relation to Fish Welfare and Its Utility as Welfare Indicator. Fish Physiol. Biochem. 2012, 38, 85–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Ceballos-Francisco, D.; Guardiola, F.A.; Esteban, M.Á. Dietary Administration of the Probiotic Shewanella putrefaciens to Experimentally Wounded Gilthead Seabream (Sparus aurata L.) Facilitates the Skin Wound Healing. Sci. Rep. 2020, 10, 11029. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Liu, M.; Tang, L.; Liu, H.; Sun, B.; Chen, L. Probiotic Intervention Mitigates the Metabolic Disturbances of Perfluorobutanesulfonate along the Gut-Liver Axis of Zebrafish. Chemosphere 2021, 284, 131374. [Google Scholar] [CrossRef] [PubMed]
- Simón, R.; Docando, F.; Nuñez-Ortiz, N.; Tafalla, C.; Díaz-Rosales, P. Mechanisms Used by Probiotics to Confer Pathogen Resistance to Teleost Fish. Front. Immunol. 2021, 12, 653025. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, S.; Schroeder, A.; Wanka, K.M. Probiotics in Fish Nutrition—Long-Standing Household Remedy or Native Nutraceuticals? Water 2021, 13, 1348. [Google Scholar] [CrossRef]
- Díaz-Rosales, P.; Salinas, I.; Rodríguez, A.; Cuesta, A.; Chabrillón, M.; Balebona, M.C.; Moriñigo, M.Á.; Esteban, M.Á.; Meseguer, J. Gilthead Seabream (Sparus aurata L.) Innate Immune Response after Dietary Administration of Heat-Inactivated Potential Probiotics. Fish Shellfish Immunol. 2006, 20, 482–492. [Google Scholar] [CrossRef]
- Domínguez-Maqueda, M.; Cerezo, I.M.; Tapia-Paniagua, S.T.; De La Banda, I.G.; Moreno-Ventas, X.; Moriñigo, M.Á.; Balebona, M.C. A Tentative Study of the Effects of Heat-Inactivation of the Probiotic Strain Shewanella putrefaciens Ppd11 on Senegalese Sole (Solea senegalensis) Intestinal Microbiota and Immune Response. Microorganisms 2021, 9, 808. [Google Scholar] [CrossRef]
- Cámara-Ruiz, M.; Balebona, M.C.; Esteban, M.Á.; Moriñigo, M.Á. Probiotic Shewanella putrefaciens (SpPdp11) as a Fish Health Modulator: A Review. Microorganisms 2020, 8, 1990. [Google Scholar] [CrossRef]
- Tapia-Paniagua, S.T.; Vidal, S.; Lobo, C.; Prieto-Álamo, M.J.; Jurado, J.; Cordero, H.; Cerezuela, R.; García de la Banda, I.; Esteban, M.A.; Balebona, M.C.; et al. The Treatment with the Probiotic Shewanella putrefaciens Pdp11 of Specimens of Solea senegalensis Exposed to High Stocking Densities to Enhance Their Resistance to Disease. Fish Shellfish Immunol. 2014, 41, 209–221. [Google Scholar] [CrossRef]
- Tapia-Paniagua, S.T.; Chabrillón, M.; Díaz-Rosales, P.; de la Banda, I.G.; Lobo, C.; Balebona, M.C.; Moriñigo, M.A. Intestinal Microbiota Diversity of the Flat Fish Solea senegalensis (Kaup, 1858) Following Probiotic Administration. Microb. Ecol. 2010, 60, 310–319. [Google Scholar] [CrossRef]
- Martínez, G.; Shaw, E.M.; Carrillo, M.; Zanuy, S. Protein Salting-out Method Applied to Genomic DNA Isolation from Fish Whole Blood. Biotechniques 1998, 24, 238–239. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Thijs, S.; De Beeck, M.O.; Beckers, B.; Truyens, S.; Stevens, V.; Van Hamme, J.D.; Weyens, N.; Vangronsveld, J. Comparative Evaluation of Four Bacteria-Specific Primer Pairs for 16S RRNA Gene Surveys. Front. Microbiol. 2017, 8, 494. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.2-0. 2014. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 20 January 2020).
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.Y.; Dillies, M.A. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [Green Version]
- Córdoba-Caballero, J.; Seoane, P.; Jabato, F.M.; Perkins, J.R.; Manchado, M.; Claros, M.G. An Improved de Novo Assembling and Polishing of Solea senegalensis Transcriptome Shed Light on Retinoic Acid Signalling in Larvae. Sci. Rep. 2020, 10, 20654. [Google Scholar] [CrossRef]
- Jabato, F.M.; Córdoba-Caballero, J.; Rojano, E.; Romá-Mateo, C.; Sanz, P.; Pérez, B.; Gallego, D.; Seoane, P.; Ranea, J.A.G.; Perkins, J.R. Gene Expression Analysis Method Integration and Co-Expression Module Detection Applied to Rare Glucide Metabolism Disorders Using ExpHunterSuite. Sci. Rep. 2021, 11, 15062. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Van Treuren, W.; Lozupone, C.; Faust, K.; Friedman, J.; Deng, Y.; Xia, L.C.; Xu, Z.Z.; Ursell, L.; Alm, E.J.; et al. Correlation Detection Strategies in Microbial Data Sets Vary Widely in Sensitivity and Precision. ISME J. 2016, 10, 1669–1681. [Google Scholar] [CrossRef] [PubMed]
- Alzola, C.F.; Harrell, F.E. An Introduction to S and the Hmisc and Design Libraries. 2004. Available online: https://hbiostat.org/R/doc/sintro.pdf (accessed on 10 October 2020).
- Tapia-Paniagua, S.T.; Ceballos-Francisco, D.; Balebona, M.C.; Esteban, M.Á.; Moriñigo, M.Á. Mucus Glycosylation, Immunity and Bacterial Microbiota Associated to the Skin of Experimentally Ulcered Gilthead Seabream (Sparus aurata). Fish Shellfish Immunol. 2018, 75, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Verbanic, S.; Shen, Y.; Lee, J.; Deacon, J.M.; Chen, I.A. Microbial Predictors of Healing and Short-Term Effect of Debridement on the Microbiome of Chronic Wounds. npj Biofilms Microbiomes 2020, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Tax, G.; Urbán, E.; Palotás, Z.; Puskás, R.; Kónya, Z.; Bíró, T.; Kemény, L.; Szabó, K. Propionic Acid Produced by Propionibacterium Acnes Strains Contributes to Their Pathogenicity. Acta Derm. Venereol. 2016, 96, 43–49. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, M.F.P.; Pereira, C.S.B.; Fregnani, J.H.; Ribeiro, F.d.A.Q. Comparative Histological Study on Wound Healing on Rat’s Skin Treated with Mitomycin C or Clobetasol Propionate. Acta Cirúrgica Bras. 2015, 30, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Tomic-Canic, M.; Burgess, J.L.; O’Neill, K.E.; Strbo, N.; Pastar, I. Skin Microbiota and Its Interplay with Wound Healing. Am. J. Clin. Dermatol. 2020, 21, 36–43. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Chapartegui-González, I.; Fernández-Martínez, M.; González-Rico, C.; Fortún, J.; Escudero, R.; Marco, F.; Linares, L.; Montejo, M.; Aranzamendi, M.; et al. Biofilm Formation by Multidrug Resistant Enterobacteriaceae Strains Isolated from Solid Organ Transplant Recipients. Sci. Rep. 2019, 9, 8928. [Google Scholar] [CrossRef] [Green Version]
- Khouadja, S.; Lamari, F.; Bakhrouf, A.; Gaddour, K. Virulence Properties, Biofilm Formation and Random Amplified Polymorphic DNA Analysis of Photobacterium Damselae Subsp. Damselae Isolates from Cultured Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax). Microb. Pathog. 2014, 69–70, 13–19. [Google Scholar] [CrossRef]
- Remuzgo-Martínez, S.; Lázaro-Díez, M.; Padilla, D.; Vega, B.; El Aamri, F.; Icardo, J.M.; Acosta, F.; Ramos-Vivas, J. New Aspects in the Biology of Photobacterium damselae subsp. piscicida: Pili, Motility and Adherence to Solid Surfaces. Vet. Microbiol. 2014, 174, 247–254. [Google Scholar] [CrossRef]
- Cameron, L.C.; Bonis, B.; Phan, C.Q.; Kent, L.A.; Lee, A.K.; Hunter, R.C. A Putative Enoyl-CoA Hydratase Contributes to Biofilm Formation and the Antibiotic Tolerance of Achromobacter Xylosoxidans. npj Biofilms Microbiomes 2019, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Gajula, B.; Munnamgi, S.; Basu, S. How Bacterial Biofilms Affect Chronic Wound Healing: A Narrative Review. Int. J. Surg. Glob. Health 2020, 3, e16. [Google Scholar] [CrossRef]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci Transl. Med. 2014, 6, 265sr6. [Google Scholar] [CrossRef] [Green Version]
- Abreu-Blanco, M.T.; Verboon, J.M.; Liu, R.; Watts, J.J.; Parkhurst, S.M. Drosophila Embryos Close Epithelial Wounds Using a Combination of Cellular Protrusions and an Actomyosin Purse String. J. Cell Sci. 2012, 125, 5984–5997. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Schultz, G.S.; Wysocki, A. Interactions between Extracellular Matrix and Growth Factors in Wound Healing. Wound Repair Regen. 2009, 17, 153–162. [Google Scholar] [CrossRef]
- Lansdown, A.B.G. Calcium: A Potential Central Regulator in Wound Healing in the Skin. Wound Repair Regen. 2002, 10, 271–285. [Google Scholar] [CrossRef]
- Caterina, M.J.; Pang, Z.; Szallasi, A.; Huang, S.M. TRP Channels in Skin Biology and Pathophysiology. Pharmaceuticals 2016, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Yuspa, S.H.; Hennings, H.; Tucker, O.W.; Jaken, S.; Kilkenny, A.E.; Roop, D.R. Signal Transduction for Proliferation and Differentiation in Keratinocytes. Ann. N. Y. Acad. Sci. 1988, 548, 191–196. [Google Scholar] [CrossRef]
- Sero, J.E.; Thodeti, C.K.; Mammoto, A.; Bakal, C.; Thomas, S.; Ingber, D.E. Paxillin Mediates Sensing of Physical Cues and Regulates Directional Cell Motility by Controlling Lamellipodia Positioning. PLoS ONE 2011, 6, e28303. [Google Scholar] [CrossRef]
- Than, U.T.T.; Guanzon, D.; Leavesley, D.; Parker, T. Association of Extracellular Membrane Vesicles with Cutaneous Wound Healing. Int. J. Mol. Sci. 2017, 18, 956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Colomé, A.M.; Lee-Rivera, I.; Benavides-Hidalgo, R.; López, E. Paxillin: A Crossroad in Pathological Cell Migration. J. Hematol. Oncol. 2017, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borodzicz, S.; Rudnicka, L.; Mirowska-Guzel, D.; Cudnoch-Jedrzejewska, A. The Role of Epidermal Sphingolipids in Dermatologic Diseases. Lipids Health Dis. 2016, 15, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, D.R.; Handsley, M.M.; Pennington, C.J. The ADAM Metalloproteinases. Mol. Aspects Med. 2008, 29, 258–289. [Google Scholar] [CrossRef]
- Zuko, A.; Oguro-Ando, A.; Post, H.; Taggenbrock, R.L.R.E.; van Dijk, R.E.; Altelaar, A.F.M.; Heck, A.J.R.; Petrenko, A.G.; van der Zwaag, B.; Shimoda, Y.; et al. Association of Cell Adhesion Molecules Contactin-6 and Latrophilin-1 Regulates Neuronal Apoptosis. Front. Mol. Neurosci. 2016, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Klein, K. Bromodomain Protein Inhibition: A Novel Therapeutic Strategy in Rheumatic Diseases. RMD Open 2018, 4, e000744. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Wu, R.; Tang, D.; Kang, R. The BET Family in Immunity and Disease. Signal Transduct. Target. Ther. 2021, 6, 23. [Google Scholar] [CrossRef]
- Sen, G.L.; Webster, D.E.; Barragan, D.I.; Chang, H.Y.; Khavari, P.A. Control of Differentiation in a Self-Renewing Mammalian Tissue by the Histone Demethylase JMJD3. Genes Dev. 2008, 22, 1865–1870. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Zhang, J.; Shen, W.; Wang, X.; Wu, J.; Wu, J.; Shi, Y. Solution Structure of the Second Bromodomain of Brd2 and Its Specific Interaction with Acetylated Histone Tails. BMC Struct. Biol. 2007, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Nogales, E.; Ciferri, C. Structure and Function of SWI/SNF Chromatin Remodeling Complexes and Mechanistic Implications for Transcription. Prog. Biophys. Mol. Biol. 2010, 102, 122–128. [Google Scholar] [CrossRef]
- Gordts, S.C.; Muthuramu, I.; Amin, R.; Jacobs, F.; De Geest, B. The Impact of Lipoproteins on Wound Healing: Topical HDL Therapy Corrects Delayed Wound Healing in Apolipoprotein E Deficient Mice. Pharmaceuticals 2014, 7, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Bo, Q.; Shao, R.; Liang, J.; Zhai, Y.; Yang, S.; Wang, F.; Sun, X. Chitinase-3-Like-1 Promotes M2 Macrophage Differentiation and Induces Choroidal Neovascularization in Neovascular Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4596–4605. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Pasos, J.; Valentín-Tirado, G.; García-Arrarás, J.E. Melanotransferrin: New Homolog Genes and Their Differential Expression during Intestinal Regeneration in the Sea Cucumber Holothuria Glaberrima. J. Exp. Zool. Part B Mol. Dev. Evol. 2017, 328, 259–274. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.M.; Sousounis, K.; Payzin-Dogru, D.; Bryant, S.; Sandoval, A.G.W.; Martinez Fernandez, J.; Mariano, R.; Oshiro, R.; Wong, A.Y.; Leigh, N.D.; et al. Identification of Regenerative Roadblocks via Repeat Deployment of Limb Regeneration in Axolotls. npj Regen. Med. 2017, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Berasain, C.; García-Trevijano, E.R.; Castillo, J.; Erroba, E.; Lee, D.C.; Prieto, J.; Avila, M.A. Amphiregulin: An Early Trigger of Liver Regeneration in Mice. Gastroenterology 2005, 128, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Seo, S.K.; Choi, I. Endogenous VSIG4 Negatively Regulates the Helper T Cell-Mediated Antibody Response. Immunol. Lett. 2015, 165, 78–83. [Google Scholar] [CrossRef]
- Xu, S.; Sun, Z.; Li, L.; Liu, J.; He, J.; Song, D.; Shan, G.; Liu, H.; Wu, X. Induction of T Cells Suppression by Dendritic Cells Transfected with VSIG4 Recombinant Adenovirus. Immunol. Lett. 2010, 128, 46–50. [Google Scholar] [CrossRef]
- Schneider, M.; Zimmermann, A.G.; Roberts, R.A.; Zhang, L.; Swanson, K.V.; Wen, H.; Davis, B.K.; Allen, I.C.; Holl, E.K.; Ye, Z.; et al. The Innate Immune Sensor NLRC3 Attenuates Toll-like Receptor Signaling via Modification of the Signaling Adaptor TRAF6 and Transcription Factor NF-ΚB. Nat. Immunol. 2012, 13, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Mo, J.; Swanson, K.V.; Wen, H.; Petrucelli, A.; Gregory, S.M.; Zhang, Z.; Schneider, M.; Jiang, Y.; Fitzgerald, K.A.; et al. NLRC3, a Member of the NLR Family of Proteins, Is a Negative Regulator of Innate Immune Signaling Induced by the DNA Sensor STING. Immunity 2014, 40, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Naya-Català, F.; do Vale Pereira, G.; Piazzon, M.C.; Fernandes, A.M.; Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Conceição, L.E.C.; Pérez-Sánchez, J. Cross-Talk Between Intestinal Microbiota and Host Gene Expression in Gilthead Sea Bream (Sparus aurata) Juveniles: Insights in Fish Feeds for Increased Circularity and Resource Utilization. Front. Physiol. 2021, 12, 748265. [Google Scholar] [CrossRef]
- Breuzard, G.; Pagano, A.; Bastonero, S.; Malesinski, S.; Parat, F.; Barbier, P.; Peyrot, V.; Kovacic, H. Tau Regulates the Microtubule-Dependent Migration of Glioblastoma Cells via the Rho-ROCK Signaling Pathway. J. Cell Sci. 2019, 132, jcs222851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilles, A.-M.; Keil, B. Cleavage of β-Casein by Collagenases from Achromobacter Iophagus and Clostridium histolyticum. FEBS Lett. 1976, 65, 369–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, N.T.; Tsugita, A.; Keil-Dlouha, V. Purification and Characterization of Two High-Molecular-Mass Forms of Achromobacter Collagenase. Biochim. Biophys. Acta—Protein Struct. Mol. Enzymol. 1986, 874, 296–304. [Google Scholar] [CrossRef]
- Shanks, R.M.Q.; Al Higaylan, M.; Stella, N.; Brothers, K.M.; Thibodeau, P.H. Bacterial Metalloproteases Inhibit Epithelial Cell Migration and Wound Healing. Investig. Ophthalmol. Vis. Sci. 2021, 62, 416. [Google Scholar]
- Mei, F.; Liu, J.; Wu, J.; Duan, Z.; Chen, M.; Meng, K.; Chen, S.; Shen, X.; Xia, G.; Zhao, M. Collagen Peptides Isolated from Salmo salar and Tilapia nilotica Skin Accelerate Wound Healing by Altering Cutaneous Microbiome Colonization via Upregulated NOD2 and BD14. J. Agric. Food Chem. 2020, 68, 1621–1633. [Google Scholar] [CrossRef]
- Bousbaine, D.; Fisch, L.I.; London, M.; Bhagchandani, P.; Rezende de Castro, T.B.; Mimee, M.; Olesen, S.; Reis, B.S.; VanInsberghe, D.; Bortolatto, J.; et al. A Conserved Bacteroidetes Antigen Induces Anti-Inflammatory Intestinal T Lymphocytes. Science 2022, 377, 660–666. [Google Scholar] [CrossRef]
- Hansen, C.R.; Pressler, T.; Nielsen, K.G.; Jensen, P.; Bjarnsholt, T.; Høiby, N. Inflammation in Achromobacter Xylosoxidans Infected Cystic Fibrosis Patients. J. Cyst. Fibros. 2010, 9, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Usui, M.L.; Lippman, S.I.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Biofilms and Inflammation in Chronic Wounds. Adv. Wound Care 2013, 2, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Tran Van Nhieu, G.; Dupont, G.; Combettes, L. Ca2+ Signals Triggered by Bacterial Pathogens and Microdomains. Biochim. Biophys. Acta—Mol. Cell Res. 2018, 1865, 1838–1845. [Google Scholar] [CrossRef]
- Dumetz, F.; Duchaud, E.; Claverol, S.; Orieux, N.; Papillon, S.; Lapaillerie, D.; Le Hénaff, M. Analysis of the Flavobacterium psychrophilum Outer-Membrane Subproteome and Identification of New Antigenic Targets for Vaccine by Immunomics. Microbiology 2008, 154, 1793–1801. [Google Scholar] [CrossRef] [Green Version]
- De Pessemier, B.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut–Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chao1 | Shannon | Simpson | |
|---|---|---|---|
| C | 743.69 ± 56.03 | 4.48 ± 0.28 | 0.97 ± 0.01 |
| P | 685.44 ± 73.15 | 3.87 ± 1.02 | 0.88 ± 0.18 |
| CW | 717.13 ± 93.15 | 4.02 ± 0.61 | 0.93 ± 0.07 |
| PW | 689.67 ± 109.17 | 3.87 ± 0.27 | 0.95 ± 0.01 |
| S.aurata IDs | D. renio Ensembl IDs | logFC | Symbol Gene | Description | |
|---|---|---|---|---|---|
| Inmune represor | LOC115567352 | ENSDARG00000105739 | 4.535 | LOC101883187 | Like NLRC3, negative regulator of the innate immune response |
| Replication | LOC115579326 | ENSDARG00000022280 | 4.847 | brd2a | Bromodomain containing 2a |
| LOC115569844 | ENSDARG00000075560 | 2.548 | kmt2cb | Lysine (K)-specific methyltransferase 2Cb | |
| LOC115575230 | ENSDARG00000089292 | 2.361 | adgrl1a | Adhesion G protein-coupled receptor L1a | |
| Celular signal | adamtsl5 | ENSDARG00000052118 | 3.74 | adamtsl5 | ADAMTS like 5 |
| itpr3 | ENSDARG00000061741 | 3.227 | itpr3 | Inositol 1,4,5-trisphosphate receptor, type 3 | |
| Celular Migration | smpd3 | ENSDARG00000098226 | 3.658 | smpd3 | Sphingomyelin phosphodiesterase 3 |
| LOC115582399 | ENSDARG00000068262 | 3.237 | vamp5 | Vesicle-associated membrane protein 5 | |
| LOC115581334 | ENSDARG00000109620 | 3.061 | pxna | Paxillin a | |
| ttbk1 | ENSDARG00000056019 | 2.96 | ttbk1a | Tau tubulin kinase 1a | |
| Apoliprotein | LOC115578371 | ENSDARG00000020866 | −6.008 | apoa4b.2 | Apolipoprotein A-IV b, tandem duplicate 2 |
| LOC115579335 | ENSDARG00000040295 | −5.883 | apoeb | Apolipoprotein Eb | |
| LOC115574970 | ENSDARG00000073718 | −4.828 | apol | Apolipoprotein L | |
| Quinases | LOC115583589 | ENSDARG00000009612 | −5.773 | chia.3 | Chitinase, acidic.3 |
| LOC115583106 | ENSDARG00000100635 | −5.216 | chia.1 | Chitinase, acidic.1 | |
| Cell proliferation | LOC115581553 | ENSDARG00000076853 | −4.973 | areg | Amphiregulin |
| meltf | ENSDARG00000075159 | −5.649 | meltf | Melanotransferrin | |
| LOC115590067 | ENSDARG00000010154 | −4.216 | vstm4a | V-set and transmembrane domain containing 4a | |
| Mucin biosynthesis | gcnt3 | ENSDARG00000060471 | −4.399 | gcnt3 | Glucosaminyl (N-acetyl) transferase 3, mucin type |
| Cell structure | LOC115570587 | ENSDARG00000075441 | −4.33 | vwa2 | Von Willebrand factor A domain containing 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerezo, I.M.; Pérez-Gómez, O.; Bautista, R.; Seoane, P.; Esteban, M.Á.; Balebona, M.C.; Moriñigo, M.A.; Tapia-Paniagua, S.T. SpPdp11 Administration in Diet Modified the Transcriptomic Response and Its Microbiota Associated in Mechanically Induced Wound Sparus aurata Skin. Animals 2023, 13, 193. https://doi.org/10.3390/ani13020193
Cerezo IM, Pérez-Gómez O, Bautista R, Seoane P, Esteban MÁ, Balebona MC, Moriñigo MA, Tapia-Paniagua ST. SpPdp11 Administration in Diet Modified the Transcriptomic Response and Its Microbiota Associated in Mechanically Induced Wound Sparus aurata Skin. Animals. 2023; 13(2):193. https://doi.org/10.3390/ani13020193
Chicago/Turabian StyleCerezo, Isabel M., Olivia Pérez-Gómez, Rocio Bautista, Pedro Seoane, M. Ángeles Esteban, M. Carmen Balebona, Miguel A. Moriñigo, and Silvana T. Tapia-Paniagua. 2023. "SpPdp11 Administration in Diet Modified the Transcriptomic Response and Its Microbiota Associated in Mechanically Induced Wound Sparus aurata Skin" Animals 13, no. 2: 193. https://doi.org/10.3390/ani13020193