
Effect of Dietary Soy Protein Source on Effluent Water Quality and Growth Performance of Rainbow Trout Reared in a Serial Reuse Water System
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Tank System and Fish Stocking
2.2. Experimental Feeds and Feeding
2.3. Fish Sampling and Physiological Analyses
2.4. Statistical Analysis
3. Results
3.1. Water Quality
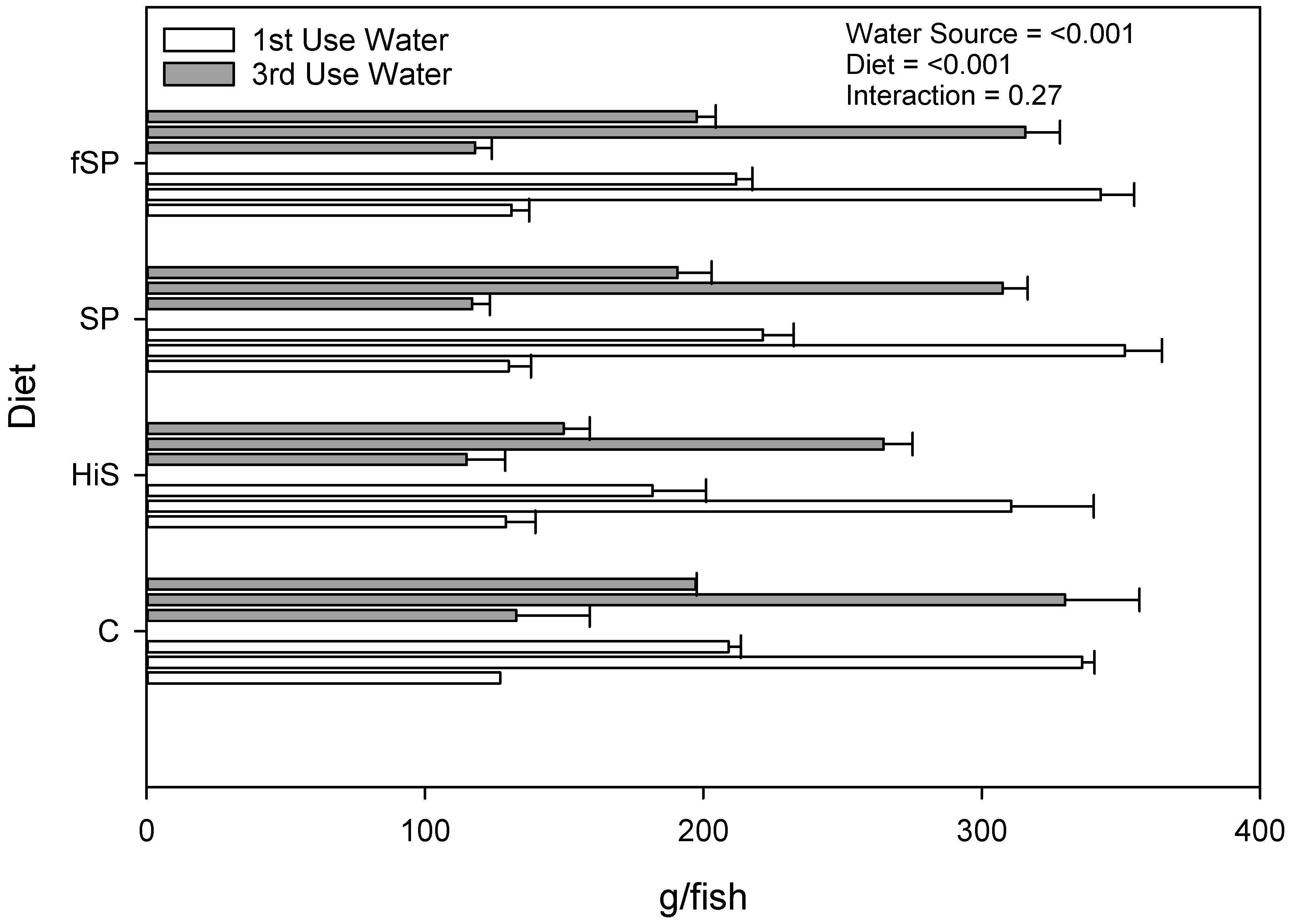
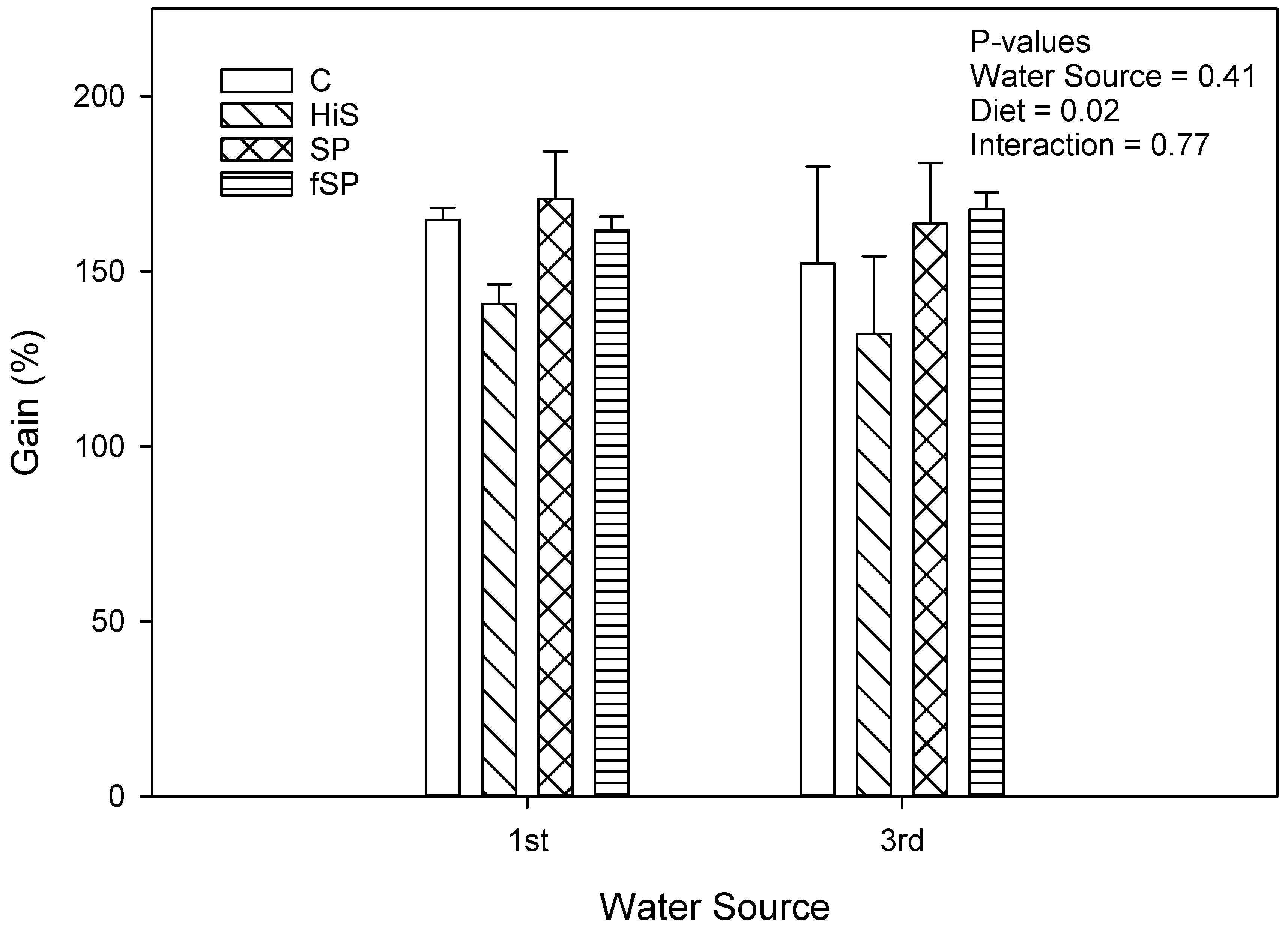
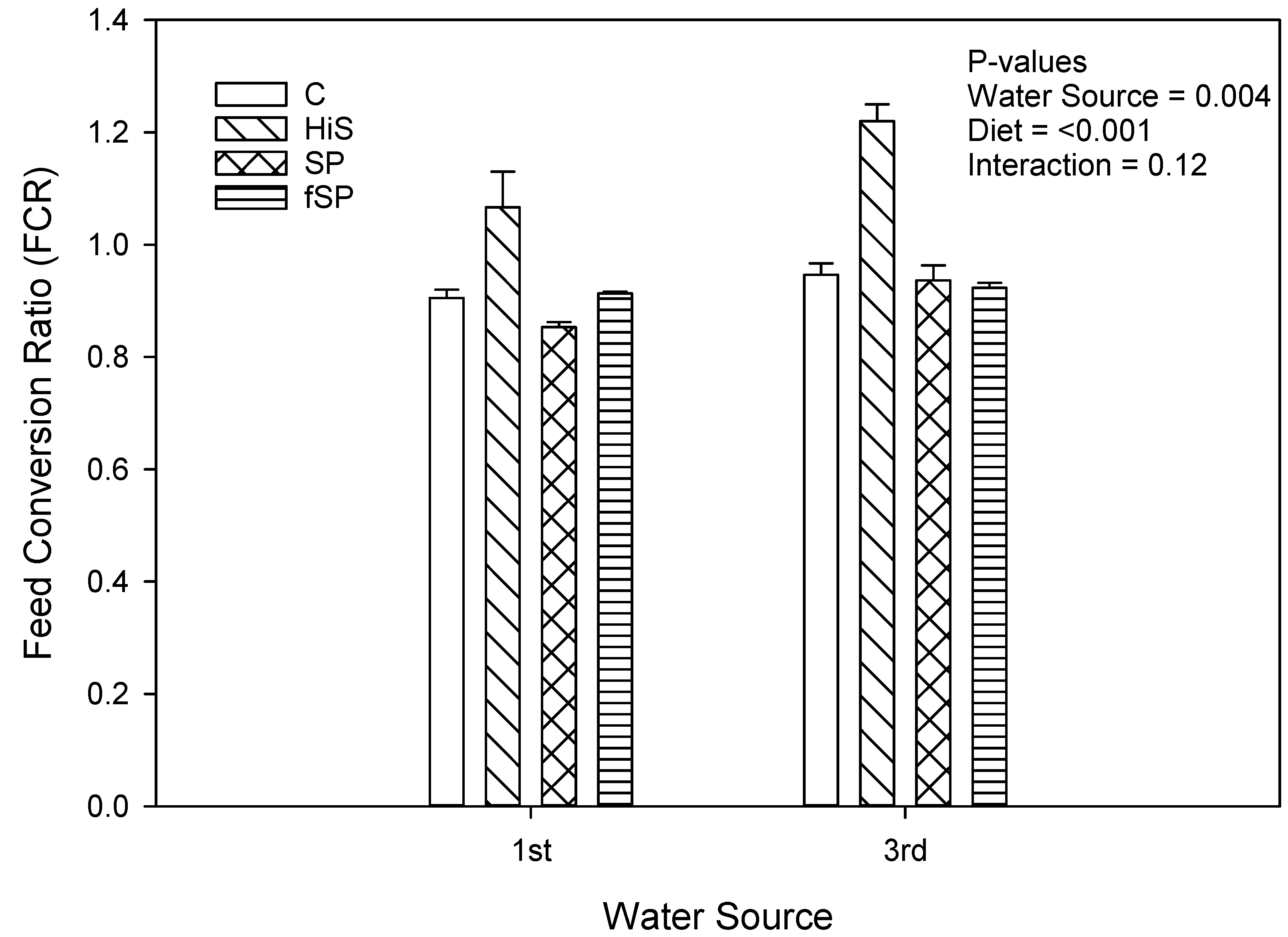
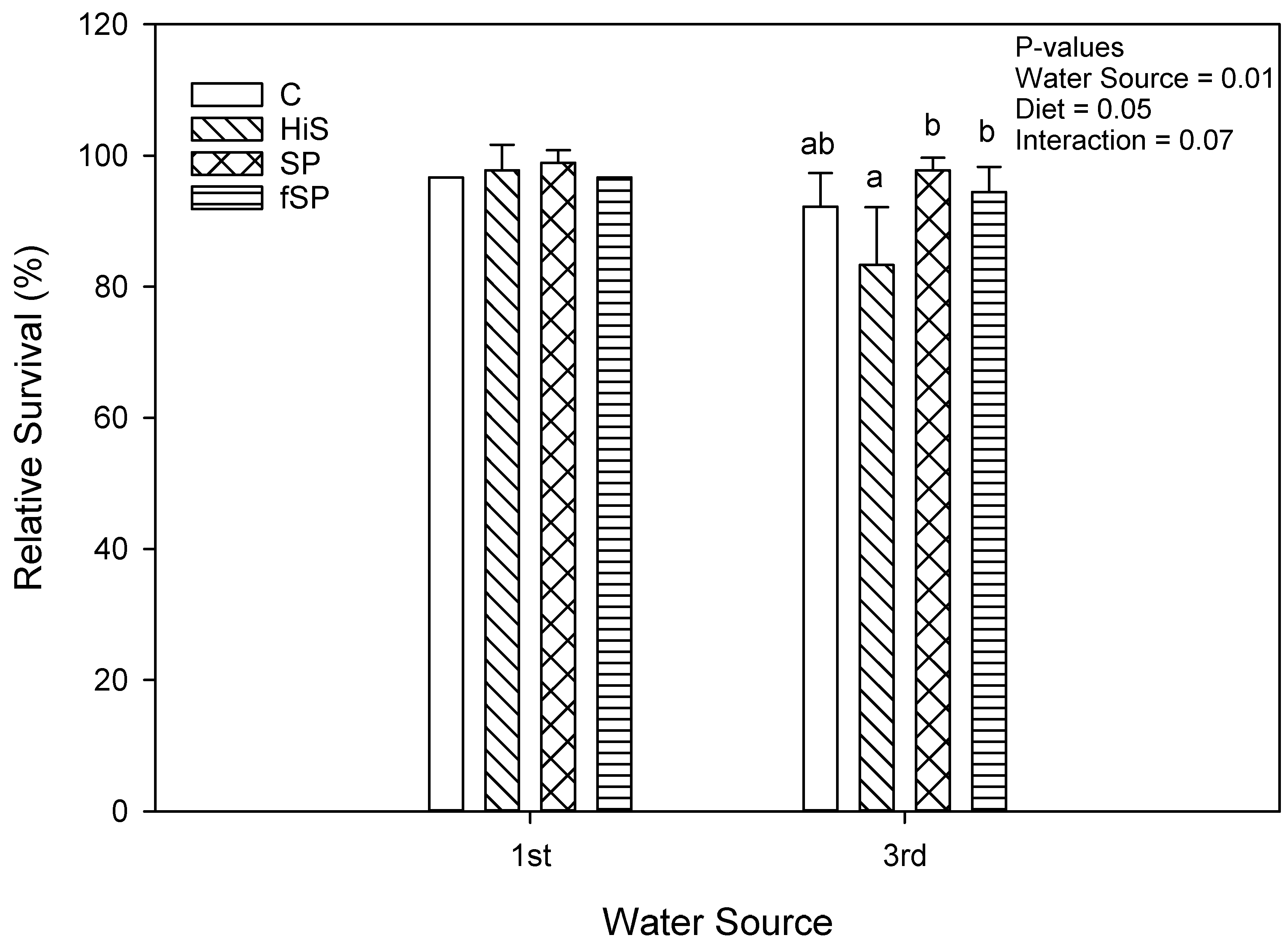
3.2. Proximate Composition and Growth Performance
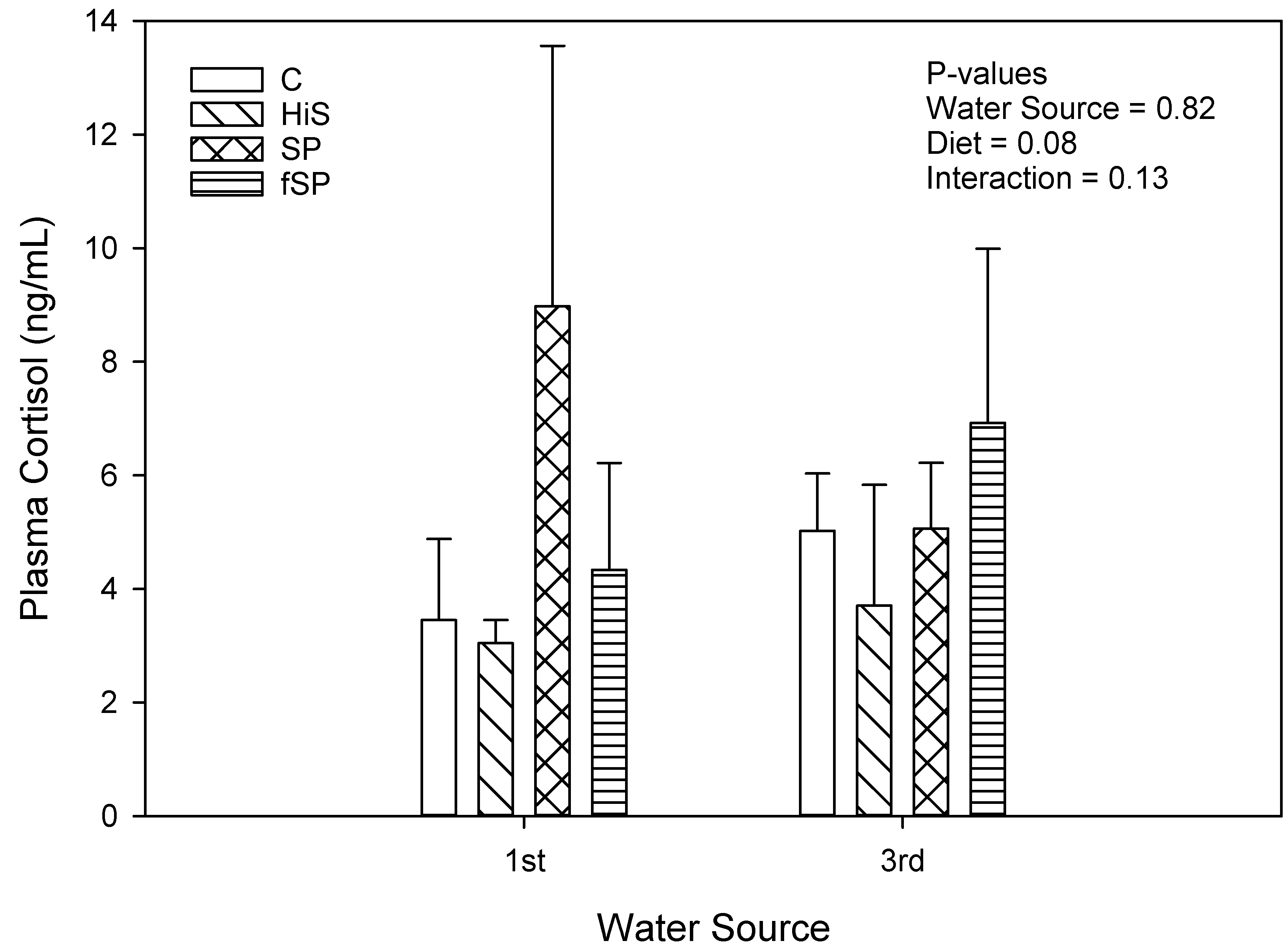
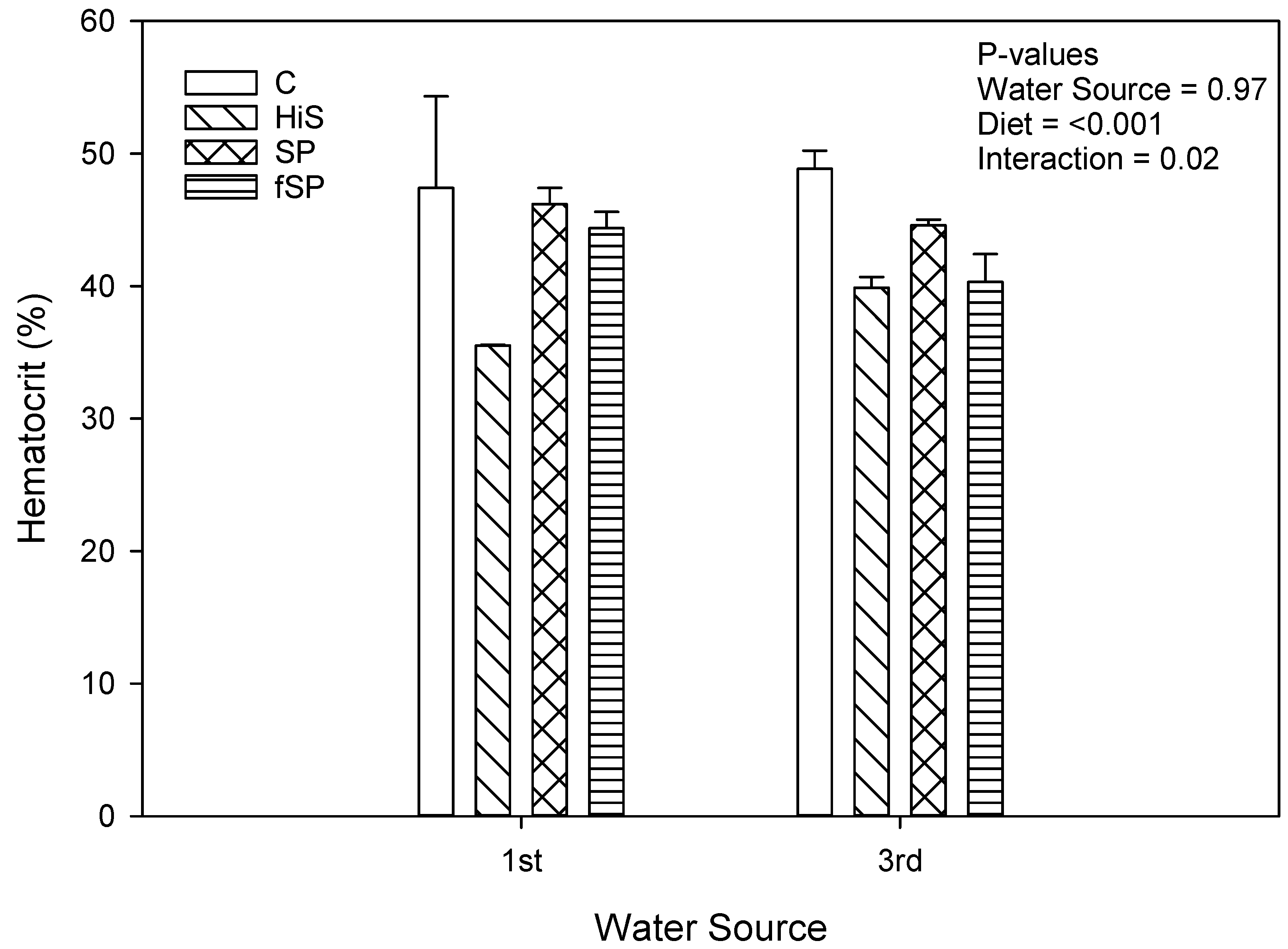
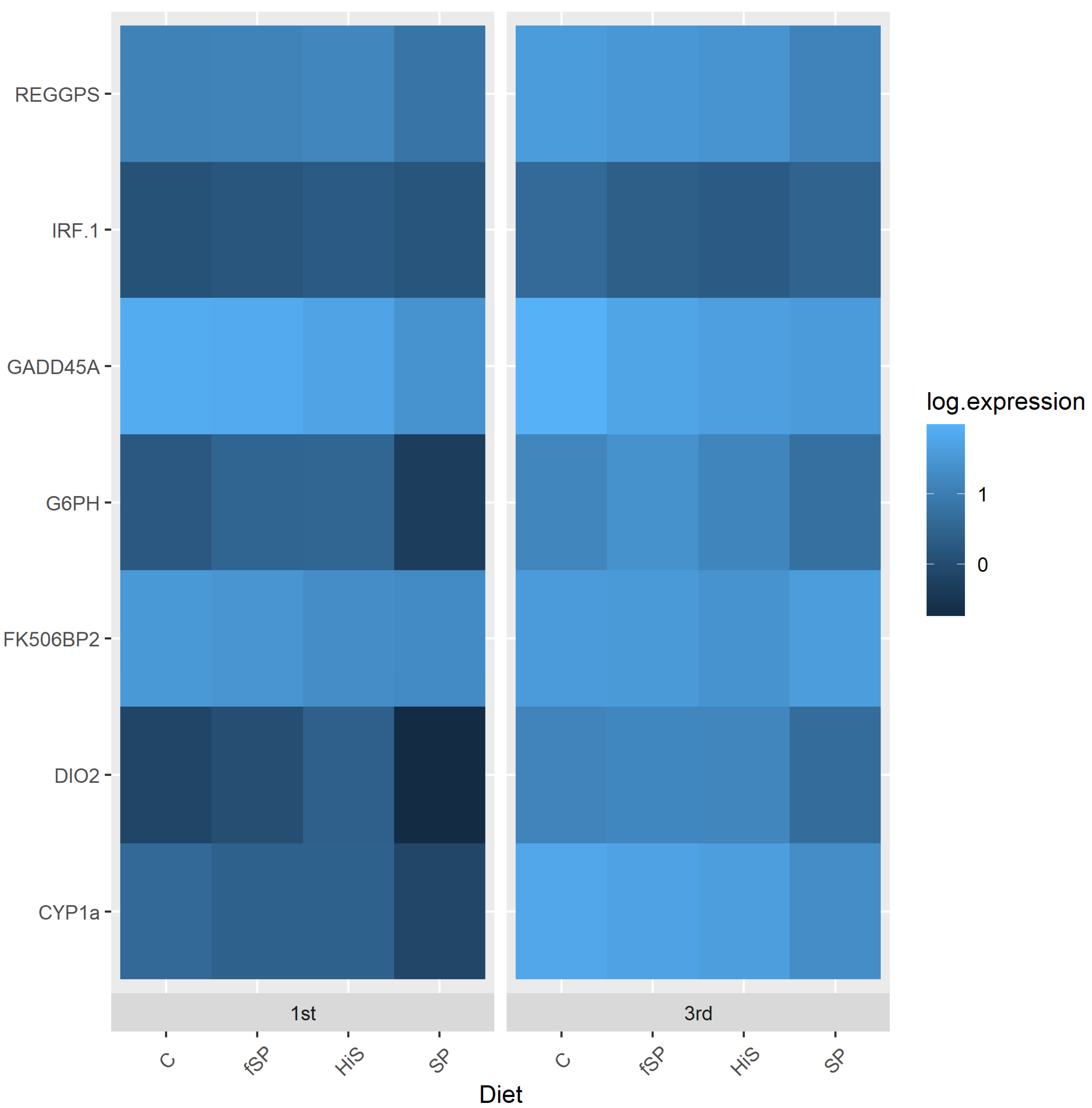
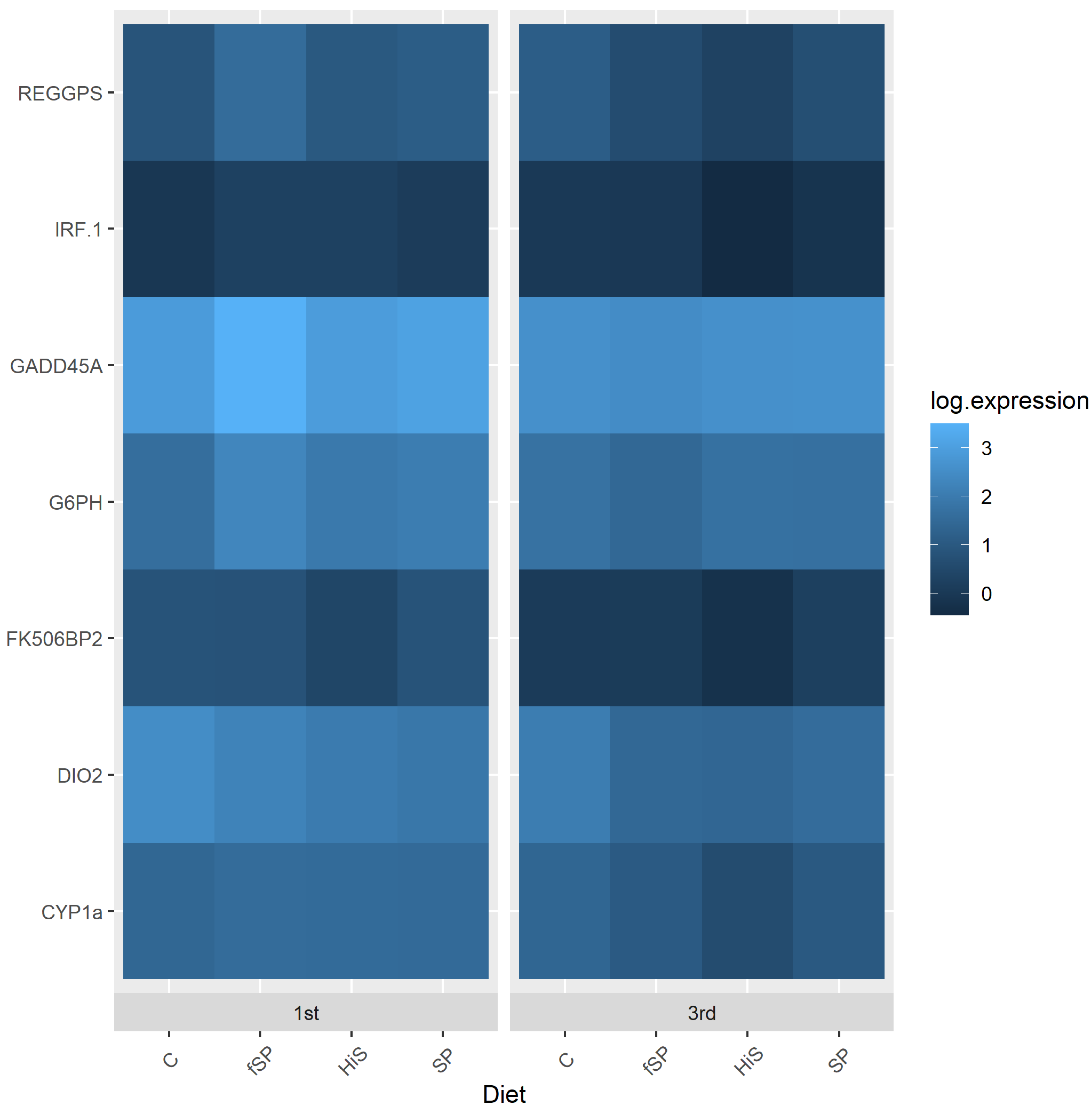
3.3. Physiological and Stress Gene Responses
4. Discussion
4.1. Water Quality
4.2. Growth Performance
4.3. Physiological and Stress Gene Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gatlin, D.I.; Barrows, F.; Brown, P.; Dabrowski, K.; Gaylord, T.; Hardy, R. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar]
- Mateo-Sagasta, J.; Zadeh, S.M.; Turral, H. More People, More Food, Worse Water? A Global Review of Water Pollution from Agriculture? FAO: Rome, Italy, 2018. [Google Scholar]
- Olsen, R.; Hasan, M. A limited supply of fishmeal: Impact on future increases in global aquaculture production. Trends Food Sci. Technol. 2012, 27, 1120–1128. [Google Scholar]
- Ayadi, F.; Muthukumarappan, K.; Rosentrater, K.; Brown, M. Twin-screw extrusion processing of rainbow trout (Oncorhynchus mykiss) feeds using various levels of corn-based distillers dried grains with solubles (DDGS). Cereal Chem. 2011, 88, 363–374. [Google Scholar]
- Francis, G.; Makkar, H.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar]
- Drew, M.; Borgeson, T.; Thiessen, D. A review of processing of feed ingredients to enhance diet digestibility in finfish. Anim. Feed Sci. Technol. 2007, 138, 118–136. [Google Scholar]
- Gaylord, T.; Barrows, F.; Rawles, S. Apparent digestibility of gross nutrients from feedstuffs in extruded feeds for rainbow trout (Oncorhynchus mykiss). J. World Aquac. Soc. 2008, 39, 827–834. [Google Scholar]
- Hardy, R. Rainbow trout, Oncorhynchus mykiss. In Nutrient Requirements and Feeding of Finfish for Aquaculture; CABI Publishing: New York, NY, USA, 2002; pp. 184–202. [Google Scholar]
- Olli, J.; Krogdahl, Å. Alcohol soluble components of soybeans seem to reduce fat digestibility in fish-meal-based diets for Atlantic salmon, Salmo salar L. Aquac. Res. 1995, 26, 831–835. [Google Scholar]
- Olli, J.; Krogdahl, Å.; Vabeno, A. Dehulled, solvent-extracted soybean meal as a protein source in diets for Atlantic salmon, Salmo salar L. Aquac. Res. 1995, 26, 167–177. [Google Scholar]
- Médale, F.; Boujard, T.; Vallée, F.; Blanc, D.; Mambrini, M.; Roem, A.; Kaushik, S. Voluntary feed intake, nitrogen and phosphorus losses in rainbow trout (Oncorhynchus mykiss) fed increasing dietary levels of soy protein concentrate. Aquat. Living Resour. 1998, 11, 239–246. [Google Scholar] [CrossRef]
- Burrells, C.; Williams, P.; Southgate, P.; Crampton, V. Immunological, physiological and pathological responses of rainbow trout (Oncorhynchus mykiss) to increasing dietary concentrations of soybean proteins. Vet. Immunol. Pathol. 1999, 72, 277–288. [Google Scholar]
- Cho, C.; Bureau, D. A review of diet formulation strategies and feeding systems to reduce excretory and feed wastes in aquaculture. Aquac. Res. 2001, 32, 349–360. [Google Scholar]
- Heikkinen, J.; Vielma, J.; Kemilainen, O.; Tiirola, M.; Eskelinen, P.; Kiuru, T.; Navia-Paldanius, D.; von Wright, A. Effects of soybean meal based diet on growth performance, gut histopathology and intestinal microbiota of juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 259–268. [Google Scholar]
- Barrows, F.; Gaylord, T.; Stone, D.; Smith, C. Effect of protein source and nutrient density on growth efficiency, histology and plasma amino acid concentration of rainbow trout (Oncorhynchus mykiss Walbaum). Aquac. Res. 2007, 38, 1747–1758. [Google Scholar]
- Kaushik, S.; Cravedi, J.; Lalles, J.; Sumpter, J.; Fauconneau, B.; Laroche, M. Partial or total replacement of fish meal by soybean protein on growth, protein utilization, potential estrogenic effects, cholesterolemia and flesh quality in rainbow trout. Aquaculture 1995, 133, 257–274. [Google Scholar]
- Adelizi, P.; Rosati, R.; Warner, K.; Wu, Y.; Muench, T.; White, M.; Brown, P. Evaluation of fish-meal free diets for rainbow trout, Onchorynchus myskiss. Aquac. Nutr. 1998, 4, 256–266. [Google Scholar]
- Bureau, D.; Harris, A.; Cho, C. The effects of purified alcohol extracts from soy products on feed intake and growth of Chinook salmon (Onchorynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss) diets. Aquaculture 1998, 161, 27–43. [Google Scholar] [CrossRef]
- Caprita, A.; Caprita, R. Modification of soluble protein content of heat-processed soybean flour. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 98–101. [Google Scholar]
- Mukhopadhyay, N.; Ray, A. Effect of fermentation on the nutritive value of sesame seed meal in the diets for rohu, Labeo rohita (Hamilton), fingerlings. Aquac. Nutr. 1999, 5, 229–236. [Google Scholar]
- Skrede, G.; Storebakken, T.; Skrede, A.; Sahlstrøm, S.; Sørensen, M.; Shearer, K.D.; Slinde, E. Lactic acid fermentation of wheat and barley whole meal flours improves digestibility of nutrients and energy in Atlantic salmon (Salmo salar L.) diets. Aquaculture 2002, 210, 305–321. [Google Scholar]
- Ng, W.; Lim, H.; Lim, S.; Ibrahim, C. Nutritive value of palm kernel meal pretreated with enzyme or fermented with Trichoderma kongii (Oudemans) as a dietary ingredient for red hybrid tilapia (Oreochromis sp.). Aquac. Res. 2002, 33, 1199–1207. [Google Scholar] [CrossRef]
- Bairagi, A.; Gosh, K.; Sen, S.; Ray, A. Evaluation of the nutritive value of Leucaena leucocephala leaf meal, inoculated with fish intestinal bacteria Bacillus subtilis and Bacillus circulans in formulated diets for roho, Labeo rohita (Hamilton) fingerlings. Aquac. Res. 2004, 35, 436–446. [Google Scholar] [CrossRef]
- Refstie, S.; Sahlström, S.; Bråthen, E.; Baeverfjord, G.; Krogedal, P. Lactic acid fermentation eliminates indigestible carbohydrates and anti-nutritional factors in soybean meal for Atlantic salmon (Salmo salar). Aquaculture 2005, 246, 331–345. [Google Scholar] [CrossRef]
- Welker, T.; Overturf, K.; Barrows, F. Development and evaluation of a volumetric quantification method for fecal particle size classification in rainbow trout (Oncorhynchus mykiss) fed different diets. N. Am. J. Aquac. 2020, 82, 159–168. [Google Scholar] [CrossRef]
- Welker, T.; Liu, K.; Overturf, K.; Abernathy, J.; Barrows, F. Effect of soy protein products and gum inclusion in feed on fecal particle size profile of rainbow trout. Aquac. J. 2021, 1, 14–25. [Google Scholar] [CrossRef]
- Piedrahita, R. Reducing the potential environmental impact of tank aquaculture effluents through intensification and recirculation. Aquaculture 2003, 226, 35–44. [Google Scholar] [CrossRef]
- Pillay, T. Aquaculture and the Environment, 2nd ed.; Blackwell Publishing: Oxford, UK, 2004. [Google Scholar]
- State of Idaho Department of Environmental Quality. Idaho Pollutant Discharge Elimination System: Effluent Limit Development Guide; State of Idaho Department of Environmental Quality: Boise, ID, USA, 2017. [Google Scholar]
- Ellis, T.; North, B.; Scott, A.; Bromage, N.; Porter, M.; Gadd, D. The relationships between stocking density and welfare in farmed rainbow trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. In Compendium of State and Regional NPDES Nutrient Permitting Approaches; U.S. Environmental Protection Agency: Washington, DC, USA, 2022.
- Rumsey, G.; Siwicki, A.; Anderson, D.; Bowser, P. Effect of soybean protein on serological response, non-specific defense mechanisms, growth and protein utilization in rainbow trout. Vet. Immunol. Immunopathol. 1994, 41, 323–339. [Google Scholar] [CrossRef]
- Brinker, A.; Friedrich, C. Fish meal replacement by plant protein substitution and guar gum addition in trout feed. Part II: Effects on faeces stability and rheology. Biorheology 2012, 49, 27–48. [Google Scholar] [CrossRef]
- Barton, B. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Gagne, F.; Smyth, S.; Andre, C.; Douville, M.; Gelinas, M.; Barclay, K. Stress-related gene expression changes in rainbow trout hepatocytes exposed to various municipal wastewater treatment influents and effluents. Environ. Sci. Pollut. Res. 2013, 20, 1706–1718. [Google Scholar] [CrossRef]
- Welker, T.; Overturf, K.; Abernathy, J. Effect of water source and trout strain on expression of stress-affected genes in a commercial setting. N. Am. J. Aquac. 2018, 80, 249–262. [Google Scholar] [CrossRef]
- Welker, T.; Overturf, K.; Snyder, S.; Liu, K.; Abernathy, J.; Frost, J.; Barrows, F. Effects of feed processing method (extrusion and expansion-compression pelleting) on water quality and growth of rainbow trout in a commercial setting. J. Appl. Aquac. 2018, 30, 97–124. [Google Scholar] [CrossRef]
- Sahlmann, C.; Sutherland, B.; Kortner, T.; Koop, B.; Krogdahl, A.; Bakke, A. Early response of gene expression in the distal intestine of Atlantic salmon (Salmo salar L.) during the development of soybean meal induced enteritis. Fish Shellfish. Immunol. 2013, 24, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Weber, G.; Gao, G.; Cleveland, B.; Yao, J.; Rexroad, C. Generation of a reference transcriptome for evaluating rainbow trout responses to various stressors. BMC Genom. 2011, 12, 626. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, A.; Koskinen, H.; Pehkonen, P.; Rexroad, C.; Afanasyev, S.; Molsa, H. Gene expression in the brain and kidney of rainbow trout in response to handling stress. BMC Genom. 2005, 6, 3. [Google Scholar] [CrossRef]
- Cairns, M.; Johnson, M.; Talbot, A.; Pemmasani, J.; McNeill, R.; Houeix, B.; Sandrador-Vegas, A.; Pottinger, T. A cDNA microarray assessment of gene expression in the liver of rainbow trout (Oncorhynchus mykiss) in response to a handling and confinement stressor. Comp. Biochem. Physiol. Part D Genom. Proteom. 2008, 3, 51–66. [Google Scholar] [CrossRef]
- Watanabe, T. Strategies for further development of aquatic feeds. Fish. Sci. 2002, 68, 242–252. [Google Scholar] [CrossRef]
- MacMillan, J.; Huddleston, T.; Wooley, M.; Fothergill, K. Best management practice development to minimize environmental impact from large flow-through trout farms. Aquaculture 2003, 226, 91–99. [Google Scholar] [CrossRef]
- US Environmental Protection Agency. Method 160.1: Residue, filterable (gravimetric, dried at 180 °C). In Methods for the Chemical Analysis of Water and Wastes (MCAWW) (EPA/600/4-79/020); US Environmental Protection Agency: Washington, DC, USA, 1971. [Google Scholar]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- Han, B.; Meng, Y.; Tian, H.; Li, C.; Gonbao, C.; Fan, W.; Ma, R. Effects of acute hypoxic stress on physiological and hepatic metabolic responses of triploid rainbow trout (Oncorhynchus mykiss). Front. Physiol. 2022, 13, 921709. [Google Scholar] [CrossRef]
- Turpeinen, U.; Hämäläinen, E. Determination of cortisol in serum, saliva and urine. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 795–801. [Google Scholar] [CrossRef]
- Overturf, K.; Welker, T.; Barrows, F.; Towner, R.; Schneider, R.; LaPatra, S. Variation in Rainbow Trout, Oncorhynchus mykiss to biosynthesize eicosapentaenoic acid and docosahexaenoic acid when reared on plant oil replacement feeds. J. World Aquac. Soc. 2013, 44, 326–337. [Google Scholar] [CrossRef]
- Sokal, R.; Rohlf, J. Biometry: The Principles and Practice of Statistics in Biological Research, 2nd ed.; W.H. Freeman and Company: San Francisco, CA, USA, 1981. [Google Scholar]
- Maillard, V.; Boardman, G.; Nyland, J.; Kuhn, D. Water quality and sludge characterization at raceway-system trout farms. Aquac. Eng. 2005, 33, 271–284. [Google Scholar] [CrossRef]
- Welker, T.; Barrows, F. Effect of diet on phosphorus and nitrogen partitioning in fecal particle size classes in rainbow trout. N. Am. J. Aquac. 2023; in press. [Google Scholar]
- Cho, C. Fish nutrition, feeds, and feeding: With special emphasis on salmonid aquaculture. Food Rev. Int. 1990, 6, 333–357. [Google Scholar] [CrossRef]
- Bureau, D.; Hua, K. Future Directions in Feed Formulation for Waste Reduction. AAC Special Publication: Quebec City, QC, Canada, 2006; Volume 11, pp. 48–52. [Google Scholar]
- Duchrow, R.M.; Everhart, W.H. Turbidity measurement. Trans. Am. Fish. Soc. 1971, 4, 682–690. [Google Scholar] [CrossRef]
- Lloyd, D. Turbidity as a water quality standard for salmonid habitats in Alaska. N. Am. J. Fish. Manag. 1987, 7, 34–45. [Google Scholar] [CrossRef]
- Gregory, R. The influence of ontogeny, perceived risk of predation, and visual ability on the foraging behavior of juvenile chinook salmon. Theory Appl. Fish Feed. Ecol. 1992, 18, 271–284. [Google Scholar]
- Boyd, C. Water Quality Management for Pond Fish Culture; Elsevier: Oxford, UK, 1982. [Google Scholar]
- Alabaster, J.; Lloyd, R. Water Quality Criteria for Freshwater Fish; Butterworth & Co Ltd.: London, UK, 1980. [Google Scholar]
- Meade, J. Allowable ammonia for fish culture. Progress. Fish-Cult. 1985, 47, 135–145. [Google Scholar] [CrossRef]
- Thurston, R.; Luedtke, R.R.R.; Smith, C.; Meyn, E.; Chakoumakos, C.; Wang, K.; Brown, C. Chronic toxicity of ammonia to rainbow trout. Trans. Am. Fish. Soc. 1984, 113, 56–73. [Google Scholar] [CrossRef]
- Smart, G. Aspects of water quality producing stress in intensive fish culture. In Stress and Fish; Academic Press: New York, NY, USA, 1981; pp. 277–293. [Google Scholar]
- Piper, R.; McElwain, I.; Orme, L.; McCraren, J.; Fowler, L.; Leonard, J. Fish Hatchery Management; US Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 1982. [Google Scholar]
- Wedemeyer, G. Physiology of Fish in Intensive Culture Systems; Chapman and Hall: New York, NY, USA, 1996. [Google Scholar]
- Delgado, C.; Rosegrant, M.; Wada, N.; Meijer, S.; Ahmed, M. Fish as food: Projections to 2020 under different scenarios. In MSSD Discussion Paper No. 52; International Food Policy Research Institute: Washington, DC, USA, 2002. [Google Scholar]
- Deak, N.; Johnson, L.; Lusas, E.; Rhee, K. Soy protein products, processing, and utilization. In Soybeans, Chemistry, Production, Processing, and Utilization; AOCS Press: Urbana, IL, USA, 2008; pp. 661–724. [Google Scholar]
- Dabrowski, K.; Poczyczynski, P.; Köck, G.; Berger, R. Effect of partially or totally replacing fishmeal protein by soybean meal protein on growth, food utilisation and proteolytic enzyme activities in rainbow trout (Salmo gairdneri). New in vivo test for endocrine pancreatic secretion. Aquaculture 1989, 77, 29–49. [Google Scholar] [CrossRef]
- Voorhees, J.; Barnes, M.; Chipps, S.; Brown, M. Bioprocessed soybean meal replacement of fish meal in rainbow trout (Oncorhynchus mykiss) diets. Cogent Food Agric. 2019, 5, 1579482. [Google Scholar] [CrossRef]
- Voorhees, J.; Barnes, M.; Chipps, S.; Brown, M. Dietary bioprocessed soybean meal does not affect the growth of exercised juvenil rainbow trout (Oncorhynchus mykiss). J. Anim. Res. Nutr. 2018, 3, 1–13. [Google Scholar]
- Burr, G.; Wolters, W.; Barrows, F.; Hardy, R. Replacing fishmeal with blends of alternative proteins on growth performance of rainbow trout (Oncorhynchus mykiss), and early or late stage juvenile Atlantic salmon (Salmo salar). Aquaculture 2012, 334–337, 110–116. [Google Scholar] [CrossRef]
- Bruce, T.; Sindelar, S.; Voorhees, J.; Brown, M.; Barnes, M. Performance and immunological responses of rainbow trout (Oncorhynchus mykiss) fed bioprocessed plant-based proteins. Aquac. Nutr. 2017, 23, 1160–1168. [Google Scholar] [CrossRef]
- Kumar, V.; Lee, S.; Cleveland, B.; Romano, N.; Lalgudi, R.; Benito, M.; Hardy, R. Comparative evaluation of processed soybean meal (EnzoMealTM) vs. regular soybean meal as a fishmeal replacement in diets of rainbow trout (Oncorhynchus mykiss): Effects on growth performance and growth-related genes. Aquaculture 2020, 516, 734652. [Google Scholar] [CrossRef]
- Mambrini, M.; Roem, A.; Carvédi, J.; Lallés, J.; Kaushik, S. Effects of replacing fish meal with soy protein concentrate and of DL-methionine supplementation in high-energy, extruded diets on the growth and nutrient utilization of rainbow trout, Oncorhynchus mykiss. J. Anim. Sci. 1999, 77, 2990–2999. [Google Scholar] [CrossRef] [PubMed]
- Brinker, A.; Rosch, R. Factors determining the size of suspended solids in a flow-through fish farm. Aquac. Eng. 2005, 33, 1–19. [Google Scholar] [CrossRef]
- Petitjean, Q.; Jean, S.; Gandar, A.; Cote, J.; Laffaille, P.; Jacquin, L. Stress responses in fish: From molecular to evolutionary processes. Sci. Total Environ. 2019, 684, 371–380. [Google Scholar] [CrossRef]
- Sopinka, N.; Donaldson, M.; O’Connor, C.; Suski, C.; Cooke, S. Stress indicators in fish. Fish Physiol. 2016, 35, 405–462. [Google Scholar]
- Seibel, H.; Babmann, B.; Rebl, A. Blood will tell: What hematological analyses can reveal about fish welfare. Front. Vet. Sci. 2021, 8, 616955. [Google Scholar] [CrossRef]
- Ings, J.; Servos, M.; Vijayan, M. Exposure to municipal wastewater effluent impacts stress performance in rainbow trout. Aquat. Toxicol. 2011, 103, 85–91. [Google Scholar] [CrossRef]
- Liu, S.; Gao, G.; Palti, Y.; Cleveland, B.; Weber, G.; Rexroad, C. RNA-seq analysis of early hepatic response to handling and confinement stress in rainbow trout. PLoS ONE 2014, 9, e88492. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Xu, G.; Nie, Z.; Xu, P.; Gu, R. Transcriptome analysis gene expression in the liver of Coilia nasus during the stress response. BMC Genom. 2014, 15, 558. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R.; Nyska, A. Invited review: Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef]
- Devyatkina, B.N.A.T.; Voskresenskii, O.; Val’dman, V. Effect of chronic emotional stress on the state of lipid peroxidation in tissue and blood of emotional and unemotional rats. Bull. Exp. Biol. Med. 1985, 100, 12–14. [Google Scholar]
- Liu, J.; Wang, X.; Shigenaga, M.; Yeo, H.; Mori, A.; Ames, B. Immobilization tress causes oxidative damage to lipid, protein, and DNA in the brain of rats. FASEB J. 1996, 10, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. WAO J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Welker, T.; Congleton, J. Relationship between dietary lipid source, oxidative stress, and the physiological response to stress in sub-yearling chinook salmon (Oncorhynchus tshawytscha). Fish Physiol. Biochem. 2003, 29, 225–235. [Google Scholar] [CrossRef]
- Welker, T.; Congleton, J. Oxidative stress in juvenile chinook salmon, Oncorhynchus tshawytscha (Walbaum). Aquac. Res. 2004, 35, 881–887. [Google Scholar] [CrossRef]
- Finkel, T. Oxidant signals and oxidative stress. Curr. Opin. Cell Biol. 2003, 15, 247–254. [Google Scholar] [CrossRef]
- Abernathy, J.; Brezas, A.; Snekvik, K.; Hardy, R.; Overturf, K. Integrative functional analyses using rainbow trout selected for tolerance to plant diets reveal nutrigenomic signature for soy utilization without the concurrence of enteritis. PLoS ONE 2017, 12, e0180972. [Google Scholar] [CrossRef]
- Saulnier, R.; Best, C.; Kostyniuk, D.; Gilmour, K.; Lamarre, S. Chronic social stress alters protein metabolism in juvenile rainbow trout, Oncorhynchus mykiss. J. Comp. Physiol. B 2021, 191, 517–530. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grams/100 g | C | SP | HiS | fSP |
|---|---|---|---|---|
| Fish meal a | 23.00 | -- | -- | -- |
| Soy protein concentrate b | -- | 24.64 | -- | -- |
| Soybean meal c | 13.38 | 13.30 | 41.25 | -- |
| Soy protein concentrate d | -- | -- | -- | 30.00 |
| Poultry meal e | 13.80 | -- | -- | 25.00 |
| Corn protein concentrate f | 5.53 | 17.54 | 14.65 | -- |
| Blood meal g | 3.20 | -- | -- | -- |
| Wheat gluten meal h | -- | -- | 6.00 | -- |
| Wheat flour i | 20.30 | 16.45 | 9.87 | 23.46 |
| Menhaden oil j | 15.80 | 19.49 | 19.49 | 14.40 |
| Lysine HCl | 1.25 | 1.79 | 1.99 | 0.87 |
| Methionine | 0.45 | 0.55 | 0.55 | 0.55 |
| Threonine | 0.45 | 0.22 | 0.38 | 0.08 |
| Taurine k | 0.50 | 0.50 | 0.50 | |
| Dicalcium phosphate | --- | 2.65 | 2.45 | 2.25 |
| Vitamin premix l | 1.00 | 1.00 | 1.00 | 1.00 |
| Choline chloride m | 0.60 | 0.60 | 0.60 | 0.60 |
| Trace min premix n | 0.10 | 0.10 | 0.10 | 0.10 |
| Vitamin C o | 0.20 | 0.20 | 0.20 | 0.20 |
| Potassium Chloride | 0.56 | 0.56 | 0.56 | 0.56 |
| Sodium Chloride | 0.28 | 0.28 | 0.28 | 0.28 |
| Magnesium oxide | 0.05 | 0.05 | 0.05 | 0.05 |
| Astaxanthin p | 0.08 | 0.08 | 0.08 | 0.08 |
| Calculated analysis, % | ||||
| Crude Protein, % | 40.1 | 40.2 | 40.2 | 40.2 |
| Lipid, % | 20.1 | 20.1 | 20.0 | 20.0 |
| Phosphorus, % | 0.97 | 0.94 | 0.92 | 0.92 |
| Measured analysis, % | ||||
| Crude Protein, % | 44.9 | 43.5 | 43.0 | 41.9 |
| Lipid, % | 18.8 | 18.0 | 18.6 | 17.7 |
| Energy, kcal/kg | 5602.5 | 5543.3 | 5646.9 | 5505.1 |
| Ash, % | 8.63 | 5.90 | 5.99 | 6.93 |
| Genebank | Primer Efficiency (%) | |||
|---|---|---|---|---|
| Gene | Accession # | Gill | Liver | Primer/Probe Sequence (Listed 5′-3′) |
| β-actin | AF254414 | 99.2 | 98.3 | BactinF: CCCTCTTCCAGCCCTCCTT |
| BactinR: AGTTGTAGGTGGTCTCGTGGATA | ||||
| BactinMGB: 6FAM-CCGCAAGACTCCATACCGA-NFQ | ||||
| FK506bp2 | NM_001165227 | 95.6 | 93.0 | FK506bp2OM-F: GAACCAGCCCTTCACATTTACTCT |
| FK506bp2OM-R: CTTCTCCCCCTCACACATTCC | ||||
| FK506bp2OM-MGB: CTGGTCCCAGCCTTTG | ||||
| Cyp1a | AF015660 | 98.9 | 96.8 | CYPF: CAGACTTCATTCCCATCCTTCGTTA |
| CYPF: CACAAAGTTGTTGAAACGGTCATTG | ||||
| CYPMGB: 6FAM-CCTGCCCAACCGCACC-NFQ | ||||
| DIO2 | NM_001123268 | 96.1 | 97.2 | DIO2F: CTCCAAAGTGGTCAAGGTTCCT |
| DIO2R: CGTGGTGCTGGTCAAGCT | ||||
| DIO2MGB: 6FAM-CCGCCGGATGCTACC-NFQ | ||||
| GADD45a | CA058640.1 | 101.8 | 95.0 | GADD45aF: CCTCCACAGGGTAATCCAGAAC |
| GADD45aR: GCTGTAACCCAGGACTCAATGTG | ||||
| GADD45aMGB: 6FAM-CTGCACTGCCATCCC-FQ | ||||
| G6PH | AF157514 | 94.2 | 95.7 | G6PHF: CCTCCTCCTCCTCACAAGCT |
| G6PHR: CAGGAGAGCACGGTACATGATTTAA | ||||
| G6PHMGB: ATGGGTGCTGTGGTCACA | ||||
| IRF-1 | NM_001124293.1 | 97.3 | 94.8 | IRF1F: GAAGACAGTCACCAAGAAACCCTTA |
| EIRF1R: GCTCAGGAACCTCTTGTCGTTT | ||||
| IRF1MGB: 6FAM-ACACTGCCTTGCTCCC-NFQ | ||||
| REGPS | BT074111 | 97.2 | 95.0 | REGPSF: TCTCATCAGGCGGAATGTGAAG |
| REGPSR: CTCTGGGCCTCGTCGAA | ||||
| REGPSPR: 6FAM-ACGCCCACCACAGTTT-NFQ | ||||
| Source | Diet | Temp | pH | DO | DO | TDS | NH3 | Turbidity |
|---|---|---|---|---|---|---|---|---|
| °C | % | mg/L | mg/L | mg/L | NTU | |||
| Diet (D) | ||||||||
| FM-C | 14.0 ± 0.7 | 7.80 ± 0.06 | 75.4 ± 9.0 | 6.62 ± 0.15 | 457.8 ± 31.4b | 0.0007 ± 0.0012 | 0.147 ± 0.227 | |
| SPC | 14.0 ± 0.8 | 7.72 ± 0.06 | 79.3 ± 11.7 | 6.90 ± 1.08 | 502.3 ± 26.0a | 0.0022 ± 0.0019 | 0.178 ± 0.288 | |
| SBM | 14.0 ± 0.8 | 7.78 ± 0.05 | 74.0 ± 11.5 | 6.96 ± 0.86 | 451.4 ± 23.7b | 0.0006 ± 0.0016 | 2.972 ± 7.363 | |
| SPC-f | 13.6 ± 0.9 | 7.76 ± 0.08 | 73.9 ± 11.4 | 6.94 ± 1.06 | 440.6 ± 19.4b | 0.0014 ± 0.0013 | 2.004 ± 2.110 | |
| Water Source (WS) | 1st Use | 13.9 ± 0.9 | 7.78 ± 0.07 | 85.2 ± 6.0x | 7.80 ± 0.13x | 452.7 ± 32.7x | 0.0011 ± 0.0016 | 0.092 ± 0.307 |
| 3rd Use | 14.0 ± 0.7 | 7.76 ± 0.07 | 69.0 ± 7.3y | 5.91 ± 0.11y | 476.6 ± 33.3y | 0.0014 ± 0.0017 | 1.939 ± 5.006 | |
| D × WS | ||||||||
| 1st Use | Inlet | 13.5 ± 0.7 | 7.84 ± 0.04 | 99.9 ± 0.5 | 9.31 ± 0.05 | 473.1 ± 21.7 | 0.0004 ± 0.0005 | 0.000 ± 0.000 |
| FM-C | 13.9 ± 0.9 | 7.81 ± 0.07 | 81.5 ± 3.2 | 7.37 ± 0.30 | 436.4 ± 22.4 | 0.0007 ± 0.0008 | 0.001 ± 0.018 | |
| SPC | 13.9 ± 0.9 | 7.73 ± 0.04 | 88.3 ± 5.9 | 7.84 ± 0.54 | 490.8 ± 17.9 | 0.0019 ± 0.0024 | 0.002 ± 0.004 | |
| SBM | 13.9 ± 1.1 | 7.79 ± 0.02 | 86.0 ± 10.3 | 7.97 ± 0.95 | 431.6 ± 12.2 | 0.0001 ± 0.0000 | 0.443 ± 0.656 | |
| SPC-f | 13.6 ± 1.4 | 7.83 ± 0.02 | 85.5 ± 0.1 | 8.02 ± 0.01 | 429.9 ± 19.8 | 0.0016 ± 0.0021 | 0.000 ± 0.020 | |
| 3rd Use | Inlet | 13.7 ± 0.5 | 7.74 ± 0.04 | 76.0 ± 9.5 | 7.07 ± 0.88 | 432.0 ± 7.6 | 0.0010 ± 0.0001 | 0.248 ± 0.201 |
| FM-C | 14.1 ± 0.7 | 7.78 ± 0.05 | 70.3 ± 9.2 | 5.87 ± 0.85 | 475.6 ± 27.0 | 0.0008 ± 0.0016 | 0.262 ± 0.261 | |
| SPC | 14.1 ± 0.8 | 7.70 ± 0.09 | 71.1 ± 8.4 | 5.96 ± 0.77 | 514.9 ± 29.4 | 0.0026 ± 0.0013 | 0.354 ± 0.330 | |
| SBM | 14.0 ± 0.7 | 7.78 ± 0.06 | 68.1 ± 6.4 | 5.96 ± 0.59 | 461.2 ± 22.1 | 0.0009 ± 0.0020 | 4.237 ± 8.989 | |
| SPC-f | 13.6 ± 0.7 | 7.71 ± 0.07 | 66.1 ± 5.7 | 5.85 ± 0.53 | 447.7 ± 19.1 | 0.0014 ± 0.0011 | 3.339 ± 1.487 | |
| ANOVA | ||||||||
| Water Source (WS) | F-value | 0.20 | 2.92 | 41.88 | 125.70 | 10.83 | 2.079 | 1.877 |
| p-value | 0.66 | 0.10 | <0.001 | <0.001 | 0.003 | 0.161 | 0.182 | |
| Diet (D) | F-value | 0.38 | 2.81 | 0.48 | 1.01 | 13.16 | 2.418 | 0.681 |
| p-value | 0.77 | 0.06 | 0.70 | 0.41 | <0.001 | 0.088 | 0.572 | |
| Interaction (D × WS) | F-value | 0.03 | 0.97 | 0.62 | 0.72 | 0.33 | 0.506 | 0.485 |
| p-value | 0.99 | 0.42 | 0.61 | 0.55 | 0.80 | 0.682 | 0.696 |
| Water Source | Diet | Energy 3 | Protein | Fat | Ash |
|---|---|---|---|---|---|
| 1st Use | Control | 6496.9 ± 70.3 | 55.8 ± 1.0 | 34.8 ± 1.1 | 7.41 ± 0.04 |
| SP | 6480.6 ± 112.8 | 56.1 ± 2.3 | 33.9 ± 3.2 | 7.45 ± 0.58 | |
| HiS | 6291.9 ± 89.1 | 58.9 ± 1.6 | 30.6 ± 1.7 | 7.64 ± 0.54 | |
| fSP | 6361.0 ± 68.0 | 58.5 ± 1.3 | 30.3 ± 0.4 | 7.56 ± 0.28 | |
| 3rd Use | Control | 6390.7 ± 56.2 | 56.8 ± 2.0 | 32.9 ± 1.4 | 7.81 ± 0.70 |
| SP | 6412.0 ± 73.6 | 57.4 ± 0.7 | 33.1 ± 0.6 | 6.96 ± 0.35 | |
| HiS | 6227.8 ± 184.6 | 59.0 ± 4.4 | 30.1 ± 5.2 | 7.81 ± 0.17 | |
| fSP | 6377.0 ± 16.7 | 57.2 ± 2.5 | 32.2 ± 1.5 | 7.09 ± 1.25 | |
| p-Value | |||||
| Water Source | 0.19 | 0.79 | 0.73 | 0.71 | |
| Diet | 0.02 | 0.25 | 0.05 | 0.47 | |
| Interaction | 0.76 | 0.77 | 0.63 | 0.56 | |
| Diet Avg. | |||||
| Control | 6443.9 ± 63.2a | 56.3 ± 1.5 | 33.9 ± 1.2a | 7.61 ± 0.37 | |
| SP | 6446.3 ± 93.2a | 56.8 ± 1.5 | 33.5 ± 1.9a | 7.21 ± 0.46 | |
| HiS | 6259.9 ± 136.9b | 59.0 ± 3.0 | 30.4 ± 3.4b | 7.73 ± 0.36 | |
| fSP | 6369.0 ± 42.3ab | 57.9 ± 1.9 | 31.3 ± 1.0ab | 7.32 ± 0.77 |
| Water Source | Diet | Interaction | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Tissue | 1st Use | 3rd Use | Expression | p-Value | HiS | C | fSP | SP | p-Value | p-Value |
| IRF-1 | Liver | 1.201 ± 0.752 | 0.880 ± 0.365 | Unchanged | 0.06 | 0.980 ± 0.710 | 1.005 ± 0.441 | 1.157 ± 0.721 | 0.987 ± 0.475 | 0.61 | 0.16 |
| Gill | 1.242 ± 0.362 x | 1.597 ± 0.577 y | Up | 0.002 | 1.372 ± 0.403 | 1.577 ± 0.781 | 1.363 ± 0.454 | 1.423 ± 0.391 | 0.89 | 0.14 | |
| FK506bp2 | Liver | 2.063 ± 1.115 x | 1.065 ± 0.386y | Down | <0.001 | 1.159 ± 0.737 a | 1.570 ± 1.056 ab | 1.672 ± 1.019 b | 1.774 ± 0.943 b | 0.01 | 0.95 |
| Gill | 4.003 ± 1.603 | 4.702 ± 1.615 | Unchanged | 0.06 | 3.940 ± 2.110 | 4.795 ± 1.501 | 4.524 ± 1.395 | 4.283 ± 1.4808 | 0.08 | 0.46 | |
| Dio2 | Liver | 8.609 ± 7.723 | 5.257 ± 4.467 | Unchanged | 0.07 | 5.809 ± 5.909 | 9.549 ± 6.745 | 6.802 ± 7.368 | 5.728 ± 5.285 | 0.15 | 0.82 |
| Gill | 0.990 ± 1.140x | 2.919 ± 2.837 y | Up | <0.001 | 2.381 ± 2.760 | 2.212 ± 2.557 | 2.191 ± 2.548 | 1.238 ± 1.501 | 0.32 | 0.61 | |
| REGPS | Liver | 3.327 ± 2.042 x | 2.097 ± 1.057 y | Down | <0.001 | 2.045 ± 1.252 a | 2.837 ± 1.142 ab | 3.326 ± 2.522 b | 2.558 ± 1.299 ab | 0.02 | 0.003 |
| Gill | 2.884 ± 0.932 x | 4.182 ± 2.332 y | Up | <0.001 | 3.709 ± 1.495 ab | 4.149 ± 2.620 a | 3.789 ± 1.766 ab | 2.676 ± 1.431 b | 0.003 | 0.86 | |
| Cyp1a | Liver | 4.588 ± 2.986 x | 2.874 ± 1.499 y | Down | 0.003 | 3.281 ± 2.828 | 4.055 ± 1.898 | 3.837 ± 2.655 | 3.664 ± 2.420 | 0.19 | 0.14 |
| Gill | 1.419 ± 0.865 x | 5.125 ± 2.724 y | Up | <0.001 | 3.322 ± 2.807 ab | 4.374 ± 3.258 a | 3.577 ± 2.947 ab | 2.309 ± 1.685 b | 0.004 | 0.63 | |
| G6PH | Liver | 7.732 ± 4.245 x | 5.281 ± 2.180 y | Down | 0.005 | 6.303 ± 3.431 | 5.511 ± 1.978 | 7.196 ± 4.903 | 6.645 ± 2.991 | 0.68 | 0.02 |
| Gill | 1.366 ± 1.7366 x | 3.139 ± 3.082 y | Up | <0.001 | 2.443 ± 2.417 | 2.463 ± 2.412 | 2.879 ± 3.682 | 1.408 ± 1.660 | 0.17 | 0.66 | |
| GADD45A | Liver | 23.275 ± 20.460 x | 13.169 ± 9.329 y | Down | 0.006 | 16.072 ± 9.539 | 15.459 ± 14.991 | 22.393 ± 21.670 | 17.661 ± 16.924 | 0.54 | 0.40 |
| Gill | 5.668 ± 2.364 | 5.820 ± 2.074 | Unchanged | 0.68 | 5.506 ± 1.808 ab | 7.046 ± 2.533 a | 6.128 ± 2.447 ab | 4.514 ± 1.181 b | 0.007 | 0.41 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Welker, T.L.; Overturf, K. Effect of Dietary Soy Protein Source on Effluent Water Quality and Growth Performance of Rainbow Trout Reared in a Serial Reuse Water System. Animals 2023, 13, 3090. https://doi.org/10.3390/ani13193090
Welker TL, Overturf K. Effect of Dietary Soy Protein Source on Effluent Water Quality and Growth Performance of Rainbow Trout Reared in a Serial Reuse Water System. Animals. 2023; 13(19):3090. https://doi.org/10.3390/ani13193090
Chicago/Turabian StyleWelker, Thomas L., and Ken Overturf. 2023. "Effect of Dietary Soy Protein Source on Effluent Water Quality and Growth Performance of Rainbow Trout Reared in a Serial Reuse Water System" Animals 13, no. 19: 3090. https://doi.org/10.3390/ani13193090