
Dietary Inclusion of Halobacterium salinarum Modulates Growth Performances and Immune Responses in Farmed Gilthead Seabream (Sparus aurata L.)
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish
2.2. Preparation of Diets
2.3. Experimental Design and Sampling
2.4. Growth Performance
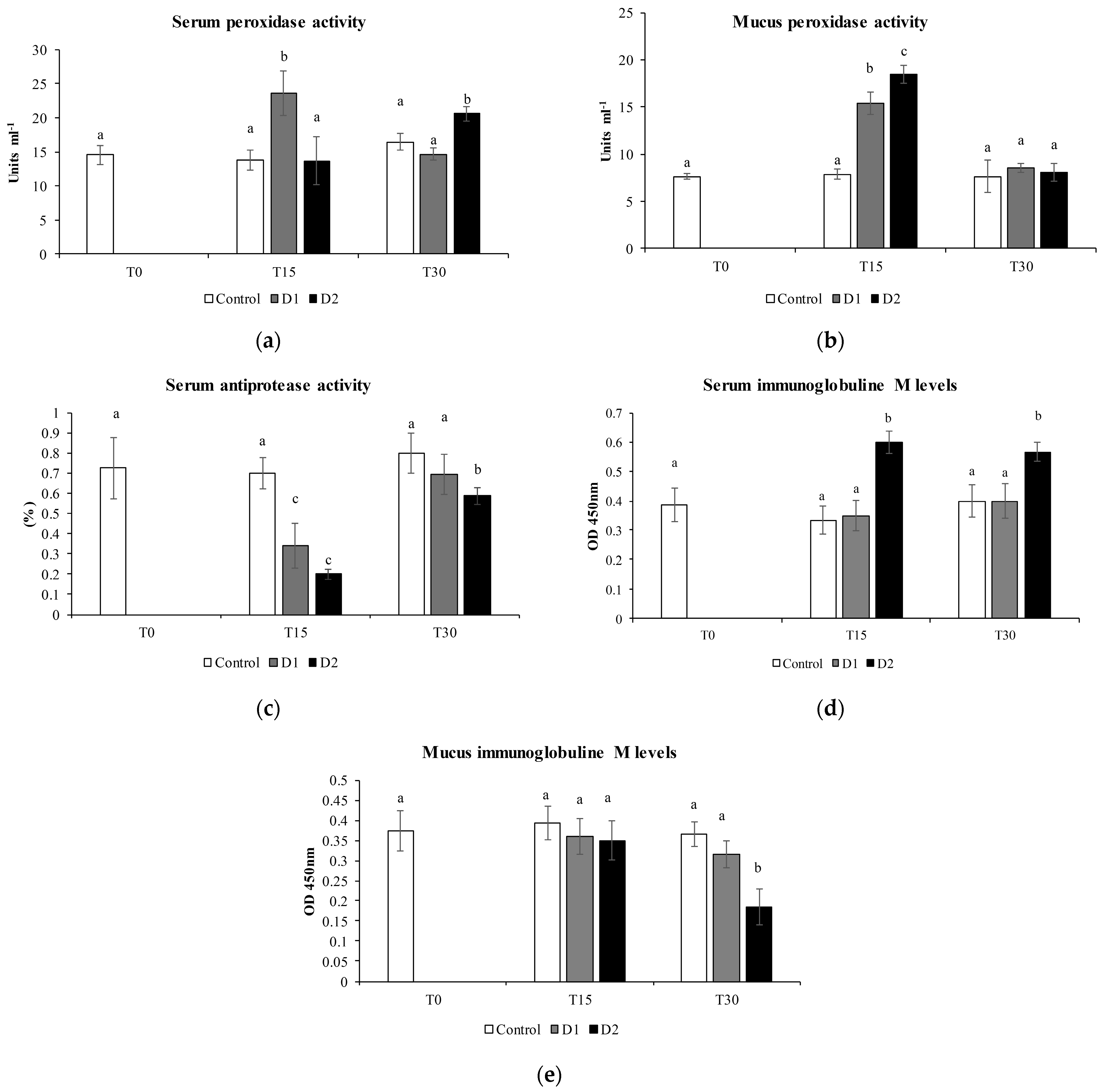
2.5. Humoral Immune Parameters
2.5.1. Peroxidase Activity
2.5.2. Protease Activity
2.5.3. Antiprotease Activity
2.5.4. Serum and Mucus IgM Levels
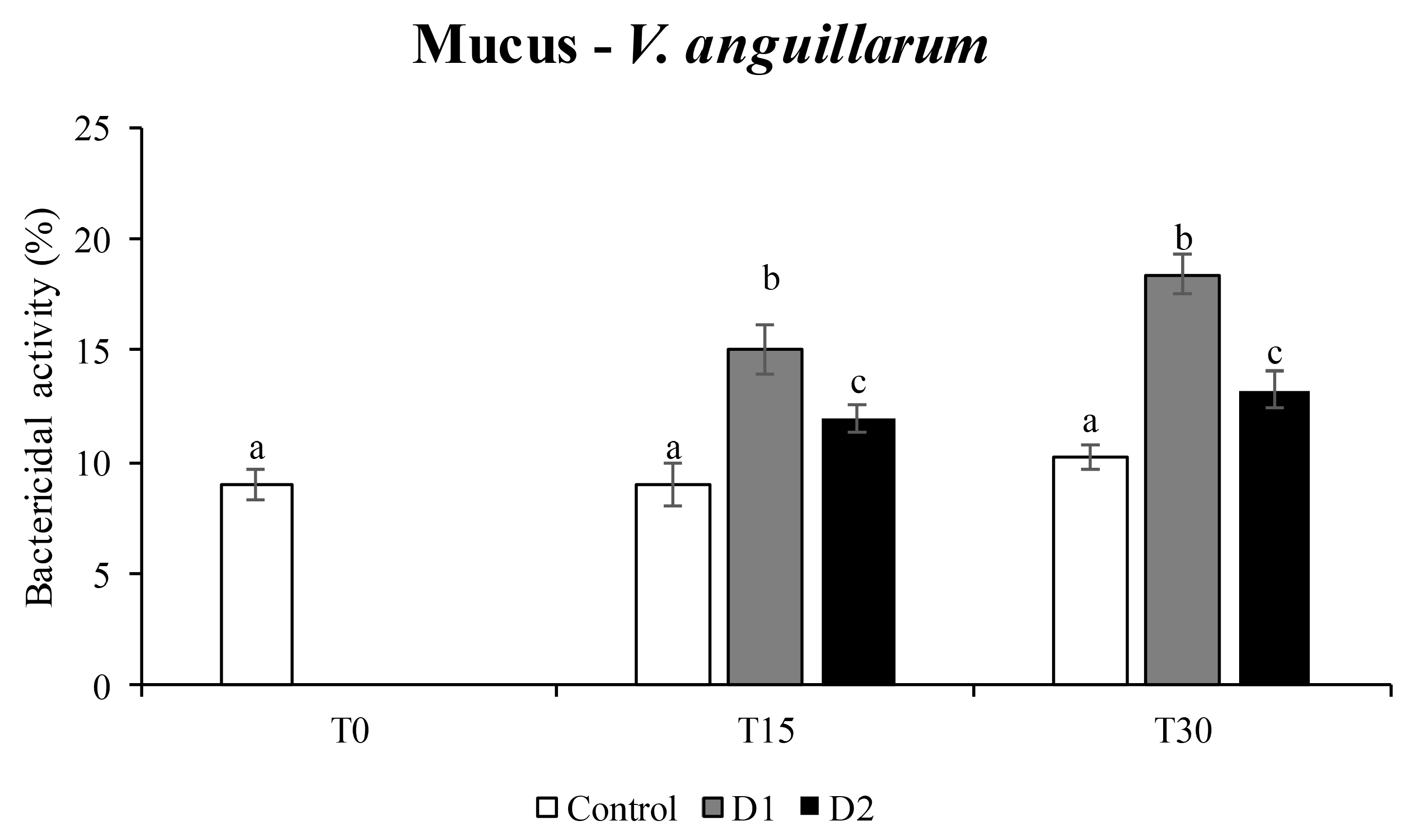
2.6. Bactericidal Activity
2.7. Cellular Immune Parameter
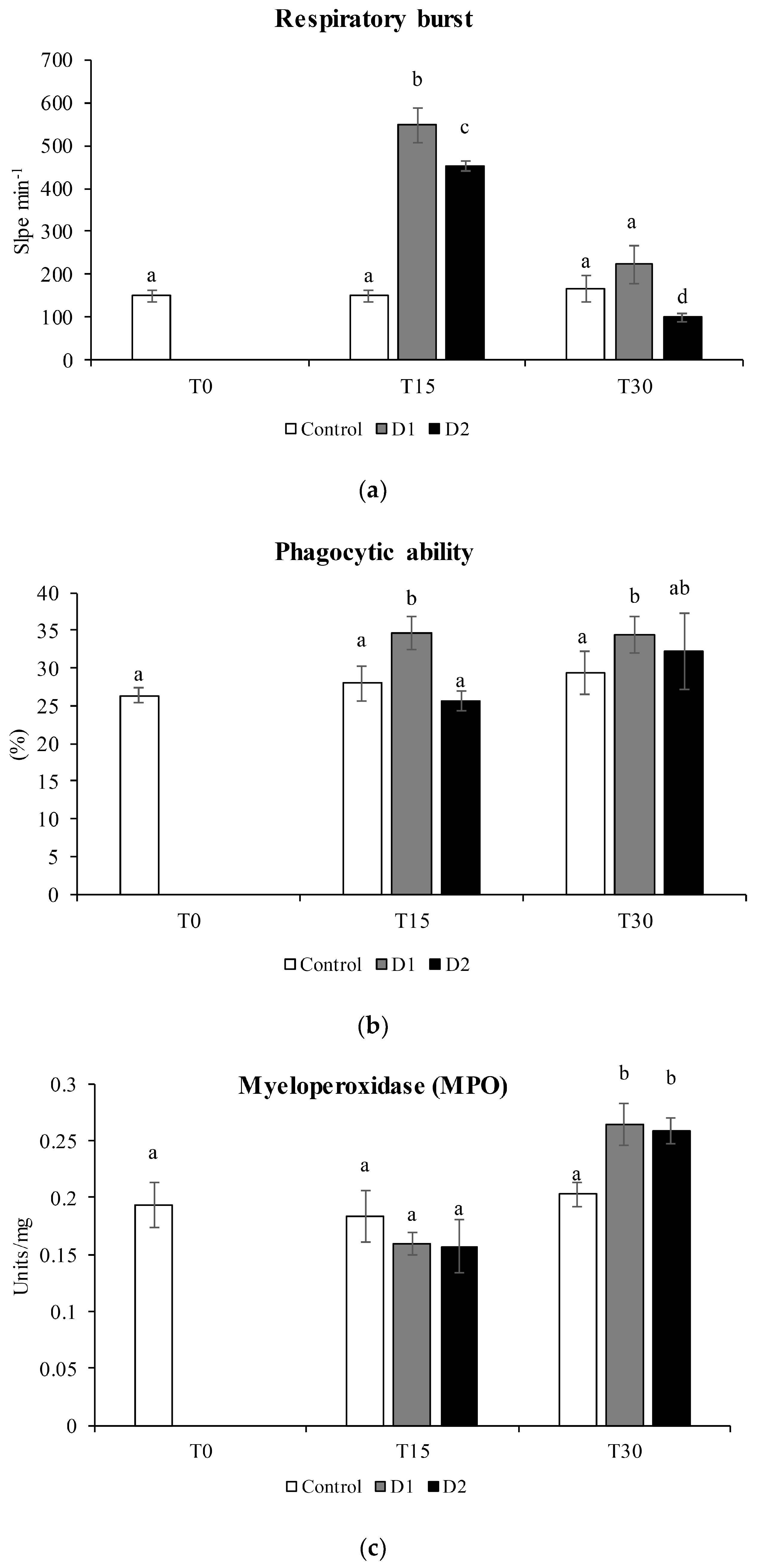
2.7.1. Respiratory Burst Activity
2.7.2. Phagocytosis
2.7.3. Peroxidase Activity
2.8. Statistical Analyses
3. Results
3.1. Growth Performances
3.2. Humoral Immune Parameters
3.3. Bactericidal Activity
3.4. Cellular Immune Parameter
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, M.; Pasquet, A.; Aubin, J.; Nahon, S.; Lecocq, T. When more is more: Taking advantage of species diversity to move towards sustainable aquaculture. Biol. Rev. 2021, 96, 767–784. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2022; FAO: Rome, Italy, 2022; ISBN 9789251363645.
- Lech, G.P.; Reigh, R.C. Plant products affect growth and digestive efficiency of cultured florida pompano (Trachinotus carolinus) fed compounded diets. PLoS ONE 2012, 7, e34981. [Google Scholar] [CrossRef]
- Elangovan, P.; Nathan, F.; Prabu Research Scholar, E.; Scholar, R.; Prabu, E.; Felix, S.; Felix, N.; Ahilan, B.; Ruby, P. An overview on significance of fish nutrition in aquaculture industry NADP-Development of cost efficient feed for enhancing freshwater fish production View project An overview on significance of fish nutrition in aquaculture industry. Int. J. Fish. Aquat. Stud. 2017, 5, 349–355. [Google Scholar]
- Magnadottir, B. Immunological control of fish diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef]
- Subedi, B.; Shrestha, A. A review: Application of probiotics in aquaculture. Int. J. For. Anim. Fish. Res. 2020, 4, 52–60. [Google Scholar]
- Okocha, R.C.; Olatoye, I.O.; Adedeji, O.B. Food safety impacts of antimicrobial use and their residues in aquaculture. Public Health Rev. 2018, 39, 21. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish, 6th ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 1–732. [Google Scholar] [CrossRef]
- Iorizzo, M.; Albanese, G.; Letizia, F.; Testa, B.; Tremonte, P.; Vergalito, F.; Lombardi, S.J.; Succi, M.; Coppola, R.; Sorrentino, E. Probiotic Potentiality from Versatile Lactiplantibacillus plantarum Strains as Resource to Enhance Freshwater Fish Health. Microorganisms 2022, 10, 463. [Google Scholar] [CrossRef]
- Chuphal, N.; Singha, K.P.; Sardar, P.; Sahu, N.P.; Shamna, N.; Kumar, V. Scope of Archaea in Fish Feed: A New Chapter in Aquafeed Probiotics? Probiotics Antimicrob. Proteins 2021, 13, 1668–1695. [Google Scholar] [CrossRef]
- Wuertz, S.; Schroeder, A.; Wanka, K.M. Probiotics in Fish Nutrition—Long-Standing Household Remedy or Native Nutraceuticals? Water 2021, 13, 1348. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish. Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in Food Systems: Significance and Emerging Strategies Towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [PubMed]
- Wan-Mohtar, W.A.A.Q.I.; Ibrahim, M.F.; Rasdi, N.W.; Zainorahim, N.; Taufek, N.M. Microorganisms as a sustainable aquafeed ingredient: A review. Aquac. Res. 2022, 53, 746–766. [Google Scholar] [CrossRef]
- Xie, W.; Deng, H.; Li, K.; Ma, Y.; Gao, M.; Duan, H.; Sui, L. Dietary supplementation of archaeal carotenoids improved antioxidative capacity and regulated immune-related gene expression of golden trout Oncorhynchus mykiss against challenge. Aquac. Res. 2022, 53, 5053–5062. [Google Scholar] [CrossRef]
- Vauclare, P.; Natali, F.; Kleman, J.P.; Zaccai, G.; Franzetti, B. Surviving salt fluctuations: Stress and recovery in Halobacterium salinarum, an extreme halophilic Archaeon. Sci. Rep. 2020, 10, 3298. [Google Scholar] [CrossRef]
- Egorova, K.; Antranikian, G. Industrial relevance of thermophilic Archaea. Curr. Opin. Microbiol. 2005, 8, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Franzetti, B.; Schoehn, G.; Ebel, C.; Gagnon, J.; Ruigrok, R.W.H.; Zaccai, G. Characterization of a Novel Complex from Halophilic Archaebacteria, Which Displays Chaperone-like Activities in Vitro. J. Biol. Chem. 2001, 276, 29906–29914. [Google Scholar] [CrossRef]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological properties of carotenoids extracted from Halobacterium halobium isolated from a Tunisian solar saltern. BMC Complement. Altern. Med. 2013, 13, 255. [Google Scholar] [CrossRef]
- Gómez-Villegas, P.; Vigara, J.; Vila, M.; Varela, J.; Barreira, L.; Léon, R. Antioxidant, antimicrobial, and bioactive potential of two new haloarchaeal strains isolated from odiel salterns (Southwest Spain). Biology 2020, 9, 298. [Google Scholar] [CrossRef]
- Giani, M.; Gervasi, L.; Loizzo, M.R.; Martínez-Espinosa, R.M. Carbon Source Influences Antioxidant, Antiglycemic, and Antilipidemic Activities of Haloferax mediterranei Carotenoid Extracts. Mar. Drugs 2022, 20, 659. [Google Scholar] [CrossRef]
- Díaz-Rosales, P.; Salinas, I.; Rodríguez, A.; Cuesta, A.; Chabrillón, M.; Balebona, M.C.; Moriñigo, M.Á.; Esteban, M.Á.; Meseguer, J. Gilthead seabream (Sparus aurata L.) innate immune response after dietary administration of heat-inactivated potential probiotics. Fish Shellfish Immunol. 2006, 20, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.A.; Mulero, V.; Muñoz, J.; Meseguer, J. Methodological aspects of assessing phagocytosis of Vibrio anguillarum by leucocytes of gilthead seabream (Sparus aurata L.) by flow cytometry and electron microscopy. Cell Tissue Res. 1998, 293, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Silva-Carrillo, Y.; Hernández, C.; Hardy, R.W.; González-Rodríguez, B.; Castillo-Vargasmachuca, S. The effect of substituting fish meal with soybean meal on growth, feed efficiency, body composition and blood chemistry in juvenile spotted rose snapper Lutjanus guttatus (Steindachner, 1869). Aquaculture 2012, 364–365, 180–185. [Google Scholar] [CrossRef]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Ross, N.W.; Firth, K.J.; Wang, A.; Burka, J.F.; Johnson, S.C. Changes in hydrolytic enzyme activities of naïve Atlantic salmon Salmo salar skin mucus due to infection with the salmon louse Lepeophtheirus salmonis and cortisol implantation. Dis. Aquat. Organ. 2000, 41, 43–51. [Google Scholar] [CrossRef]
- Rodríguez, A.; Esteban, M.Á.; Meseguer, J. Phagocytosis and peroxidase release by seabream ( Sparus aurata L.) leucocytes in response to yeast cells. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2003, 272A, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.G.; Kehrli, M.E.; Canning, P.C. A colorimetric assay for quantitating bovine neutrophil bactericidal activity. Vet. Immunol. Immunopathol. 1991, 28, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival: Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Bayne, C.J.; Levy, S. Modulation of the Oxidative Burst in Trout Myeloid Cells by Adrenocorticotropic Hormone and Catecholamines: Mechanisms of Action. J. Leukoc. Biol. 1991, 50, 554–560. [Google Scholar] [CrossRef]
- Espinosa, C.; García Beltrán, J.M.; Messina, C.M.; Esteban, M.Á. Effect of Jasonia glutinosa on immune and oxidative status of gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2020, 100, 58–69. [Google Scholar] [CrossRef]
- Talpur, A.D. Mentha piperita (Peppermint) as feed additive enhanced growth performance, survival, immune response and disease resistance of Asian seabass, Lates calcarifer (Bloch) against Vibrio harveyi infection. Aquaculture 2014, 420–421, 71–78. [Google Scholar] [CrossRef]
- Nahid Akter, M.; Parvez, I.; Parveen Patwary, Z.; Mst Nahid Akter, C. Beneficial effects of probiotics in aquaculture. Int. J. Fish. Aquat. Stud. 2016, 4, 494–499. [Google Scholar]
- Magnadóttir, B. Innate immunity of fish (overview). Fish Shellfish. Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Bradley, G.; Baker, R.T.M.; Davies, S.J. Probiotic applications for rainbow trout (Oncorhynchus mykiss Walbaum) II. Effects on growth performance, feed utilization, intestinal microbiota and related health criteria postantibiotic treatment. Aquac. Nutr. 2010, 16, 496–503. [Google Scholar] [CrossRef]
- Varela, J.L.; Ruiz-Jarabo, I.; Vargas-Chacoff, L.; Arijo, S.; León-Rubio, J.M.; García-Millán, I.; Martín del Río, M.P.; Moriñigo, M.A.; Mancera, J.M. Dietary administration of probiotic Pdp11 promotes growth and improves stress tolerance to high stocking density in gilthead seabream Sparus auratus. Aquaculture 2010, 309, 265–271. [Google Scholar] [CrossRef]
- Chauhan, A.; Singh, R. Probiotics in aquaculture: A promising emerging alternative approach. Symbiosis 2019, 77, 99–113. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef]
- Barbosa, T.M.; Serra, C.R.; La Ragione, R.M.; Woodward, M.J.; Henriques, A.O. Screening for Bacillus isolates in the broiler gastrointestinal tract. Appl. Environ. Microbiol. 2005, 71, 968–978. [Google Scholar] [CrossRef]
- Cherif, A.; Ouzari, H.; Daffonchio, D.; Cherif, H.; Ben Slama, K.; Hassen, A.; Jaoua, S.; Boudabous, A. Thuricin 7: A novel bacteriocin produced by Bacillus thuringiensis BMG1.7, a new strain isolated from soil. Lett. Appl. Microbiol. 2001, 32, 243–247. [Google Scholar] [CrossRef]
- Sakai, M. Current research status of fish immunostimulants. Aquaculture 1999, 172, 63–92. [Google Scholar] [CrossRef]
- El-Haroun, E.R.; Goda, A.M.A.S.; Kabir Chowdhury, M.A. Effect of dietary probiotic Biogen® supplementation as a growth promoter on growth performance and feed utilization of Nile tilapia Oreochromis niloticus (L.). Aquac. Res. 2006, 37, 1473–1480. [Google Scholar] [CrossRef]
- Kennedy, J.; Baker, P.; Piper, C.; Cotter, P.D.; Walsh, M.; Mooij, M.J.; Bourke, M.B.; Rea, M.C.; O’Connor, P.M.; Ross, R.P.; et al. Isolation and analysis of bacteria with antimicrobial activities from the marine sponge haliclona simulans collected from irish waters. Mar. Biotechnol. 2009, 11, 384–396. [Google Scholar] [CrossRef]
- Liu, C.-H.; Wu, K.; Chu, T.-W.; Wu, T.-M. Dietary supplementation of probiotic, Bacillus subtilis E20, enhances the growth performance and disease resistance against Vibrio alginolyticus in parrot fish (Oplegnathus fasciatus). Aquac. Int. 2018, 26, 63–74. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic Bacteria as Biological Control Agents in Aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The mucosal immune system of fish: The evolution of tolerating commensals while fighting pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Whyte, S.K. The innate immune response of finfish—A review of current knowledge. Fish Shellfish Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef]
- Subramanian, S.; MacKinnon, S.L.; Ross, N.W. A comparative study on innate immune parameters in the epidermal mucus of various fish species. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 148, 256–263. [Google Scholar] [CrossRef]
- Dash, S.; Das, S.K.; Samal, J.; Thatoi, H.N. Epidermal mucus, a major determinant in fish health: A review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar]
- Brunt, J.; Newaj-Fyzul, A.; Austin, B. The development of probiotics for the control of multiple bacterial diseases of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2007, 30, 573–579. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.M.; Austin, B. Kocuria SM1 controls vibriosis in rainbow trout (Oncorhynchus mykiss, Walbaum). J. Appl. Microbiol. 2010, 108, 2162–2170. [Google Scholar] [CrossRef] [PubMed]
- Parra, D.; Reyes-Lopez, F.E.; Tort, L. Mucosal immunity and B cells in teleosts: Effect of vaccination and stress. Front. Immunol. 2015, 6, 354. [Google Scholar] [CrossRef]
- García Beltrán, J.M.; Espinosa, C.; Guardiola, F.A.; Esteban, M.Á. Dietary dehydrated lemon peel improves the immune but not the antioxidant status of gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2017, 64, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B. Phagocytosis induces superoxide formation and apoptosis in macrophages. Exp. Mol. Med. 2003, 35, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Cathcart, M.K. Regulation of Superoxide Anion Production by NADPH Oxidase in Monocytes/Macrophages: Contributions to Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, S.J. Myeloperoxidase: Friend and foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef] [PubMed]
- Biller-Takahashi, J.D.; Takahashi, L.S.; Saita, M.V.; Gimbo, R.Y.; Urbinati, E.C. Leukocytes respiratory burst activity as indicator of innate immunity of pacu Piaractus mesopotamicus. Brazilian J. Biol. 2013, 73, 425–429. [Google Scholar] [CrossRef]
- Simón, R.; Docando, F.; Nuñez-Ortiz, N.; Tafalla, C.; Díaz-Rosales, P. Mechanisms Used by Probiotics to Confer Pathogen Resistance to Teleost Fish. Front. Immunol. 2021, 12, 1259. [Google Scholar] [CrossRef]
- Nikoskelainen, S.; Ouwehand, A.C.; Bylund, G.; Salminen, S.; Lilius, E.M. Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol. 2003, 15, 443–452. [Google Scholar] [CrossRef]
- Gullian, M.; Thompson, F.; Rodriguez, J. Selection of probiotic bacteria and study of their immunostimulatory effect in Penaeus vannamei. Aquaculture 2004, 233, 1–14. [Google Scholar] [CrossRef]
- Park, Y.; Lee, S.; Hong, J.; Kim, D.; Moniruzzaman, M.; Bai, S.C. Use of probiotics to enhance growth, stimulate immunity and confer disease resistance to Aeromonas salmonicida in rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2017, 48, 2672–2682. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control | D1 | D2 | |
|---|---|---|---|
| Weight | 11.48 ± 1.9 a | 13.18 ± 0.83 a | 15.11 ± 1.1 b |
| WG | 68.33 ± 1.70 a | 72 ± 1.24 a | 77.44 ± 1.43 b |
| SGR | 1.89 ± 0.78 a | 3.95 ± 0.86 ab | 5.80 ± 0.87 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messina, C.M.; Madia, M.; Manuguerra, S.; Espinosa-Ruiz, C.; Esteban, M.A.; Santulli, A. Dietary Inclusion of Halobacterium salinarum Modulates Growth Performances and Immune Responses in Farmed Gilthead Seabream (Sparus aurata L.). Animals 2023, 13, 2743. https://doi.org/10.3390/ani13172743
Messina CM, Madia M, Manuguerra S, Espinosa-Ruiz C, Esteban MA, Santulli A. Dietary Inclusion of Halobacterium salinarum Modulates Growth Performances and Immune Responses in Farmed Gilthead Seabream (Sparus aurata L.). Animals. 2023; 13(17):2743. https://doi.org/10.3390/ani13172743
Chicago/Turabian StyleMessina, Concetta Maria, Manfredi Madia, Simona Manuguerra, Cristobal Espinosa-Ruiz, María Angeles Esteban, and Andrea Santulli. 2023. "Dietary Inclusion of Halobacterium salinarum Modulates Growth Performances and Immune Responses in Farmed Gilthead Seabream (Sparus aurata L.)" Animals 13, no. 17: 2743. https://doi.org/10.3390/ani13172743