
Microsporidia in Commercially Harvested Marine Fish: A Potential Health Risk for Consumers
 , , , ,
, , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Fish Sampling
2.2. DNA Extraction
2.3. Real-Time PCR for Human Pathogenic Microsporidia Detection and Species Identification
3. Results
3.1. Fish Sampled
3.2. Zoonotic Microsporidia in Gastrointestinal Tract Samples
3.3. Zoonotic Microsporidia in Muscle Samples
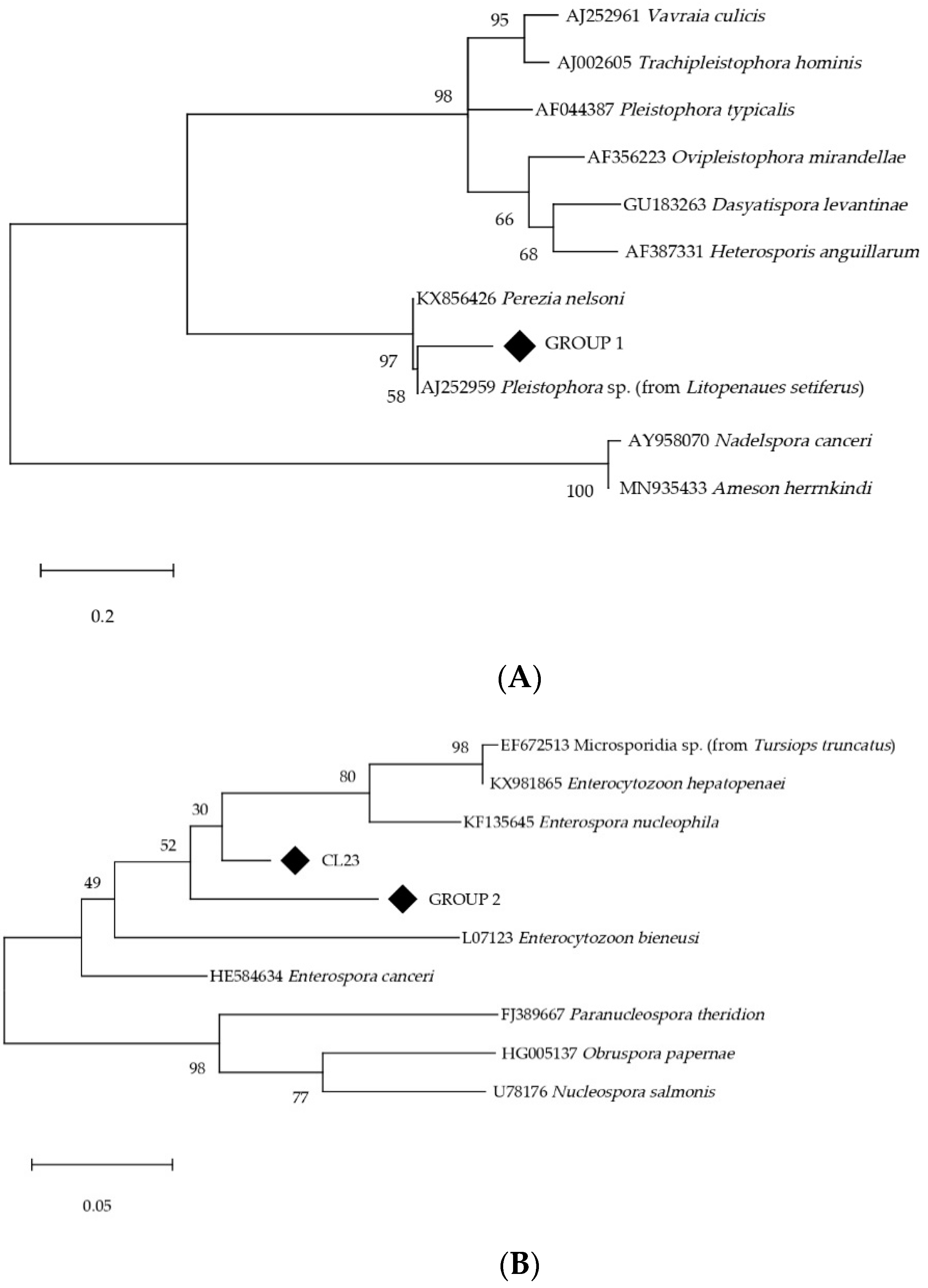
3.4. Other Microsporidia Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [PubMed]
- Cali, A.; Takvorian, P.M. Developmental morphology and life cycles of the Microsporidia. In Microsporidia: Pathogens of Opportunity, 1st ed.; Weiss, L.M., Becnel, J.J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; pp. 71–133. [Google Scholar]
- Han, B.; Takvorian, P.M.; Weiss, L.M. The Function and Structure of the Microsporidia Polar Tube. Exp. Suppl. 2022, 114, 179–213. [Google Scholar] [CrossRef] [PubMed]
- Cali, A.; Becnel, J.J.; Takvorian, P.M. Microsporidia. In Handbook of the Protists; Archibald, J., Simpson, A., Slamovits, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 1559–1618. [Google Scholar]
- Stentiford, G.D.; Feist, S.W.; Stone, D.M.; Bateman, K.S.; Dunn, A.M. Microsporidia: Diverse, dynamic, and emergent pathogens in aquatic systems. Trends Parasitol. 2013, 29, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Lom, J.; Dyková, I. Microsporidian xenomas in fish seen in wider perspective. Folia Parasitol. 2005, 52, 69–81. [Google Scholar] [CrossRef]
- Figueras, A.; Novoa, B.; Santarem, M.; Martinez, E.; Alvarez, J.; Toranzo, A.; Dykova, I. Tetramicra brevifilum, a potential threat to farmed turbot Scophthalmus maximus. Dis. Aquat. Organ. 1992, 14, 127–135. [Google Scholar] [CrossRef]
- Palenzuela, O.; Redondo, M.J.; Cali, A.; Takvorian, P.M.; Alonso-Naveiro, M.; Alvarez-Pellitero, P.; Sitjà-Bobadilla, A. A new intranuclear microsporidium, Enterospora nucleophila n. sp., causing an emaciative syndrome in a piscine host (Sparus aurata), prompts the redescription of the family Enterocytozoonidae. Int. J. Parasitol. 2014, 44, 189–203. [Google Scholar] [CrossRef]
- Monaghan, S.R.; Michael, L.K.; Watral, V.G.; Kaufman, R.J.; Lee, L.E.J.; Bols, N.C. Animal cell cultures in microsporidial research: Their general roles and their specific use for fish microsporidia. Vitr. Cell. Dev. Biol. Anim. 2009, 45, 135–147. [Google Scholar] [CrossRef]
- Han, B.; Weiss, L.M. Microsporidia: Obligate intracellular pathogens within the fungal kingdom. Microbiol. Spectrum 2017, 5, FUNK-0018-2016. [Google Scholar] [CrossRef]
- Mathis, A.; Weber, R.; Deplazes, P. Zoonotic potential of the microsporidia. Clin. Microbiol. Rev. 2005, 18, 423–445. [Google Scholar] [CrossRef]
- Stentiford, G.; Becnel, J.; Weiss, L.; Keeling, P.; Didier, E.; Williams, B.; Bjornson, S.; Kent, M.; Freeman, M.; Brown, M.; et al. Microsporidia–Emergent Pathogens in the Global Food Chain. Trends Parasitol. 2016, 32, 336–348. [Google Scholar] [CrossRef]
- Qiu, L.; Xia, W.; Li, W.; Ping, J.; Ding, S.; Liu, H. The prevalence of microsporidia in China: A systematic review and meta-analysis. Sci. Rep. 2019, 9, 3174. [Google Scholar] [CrossRef]
- Weiss, L.M. Clinical syndromes associated with microsporidiosis. In Microsporidia: Pathogens of Opportunity, 1st ed.; Weiss, L.M., Becnel, J.J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; pp. 371–401. [Google Scholar]
- Del Aguila, C.; Navajas, R.; Gurbindo, D.; Ramos, J.T.; Mellado, M.J.; Fenoy, S.; Fernandez, M.A.M.; Subirats, M.; Ruiz, J.; Pleniazek, N.J. Microsporidiosis in HIV-Positive Children in Madrid (Spain). J. Eukaryot Microbiol. 1997, 44, 84s–85s. [Google Scholar] [CrossRef] [PubMed]
- Lores, B.; López-Miragaya, I.; Arias, C.; Fenoy, S.; Torres, J.; Del Aguila, C. Intestinal microsporidiosis due to Enterocytozoon bieneusi in elderly human immunodeficiency virus-negative patients from Vigo, Spain. Clin. Infect Dis. 2002, 34, 918–921. [Google Scholar] [CrossRef] [PubMed]
- López-Vélez, R.; Turrientes, M.C.; Garrón, C.; Montilla, P.; Navajas, R.; Fenoy, S.; Aguila, C. Microsporidiosis in travelers with diarrhea from the tropics. J. Travel Med. 1999, 6, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Acosta, N.; Lorenzo-Morales, J.; Leal-Guio, Y.; Coronado-Álvarez, N.; Foronda, P.; Alcoba-Florez, J.; Izquierdo, F.; Batista-Díaz, N.; Del Águila, C.; Valladares, B. Enterocytozoon bieneusi (microsporidia) in clinical samples from immunocompetent individuals in Tenerife, Canary Islands, Spain. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Dowd, S.E.; John, D.; Eliopolus, J.; Gerba, C.P.; Naranjo, J.; Klein, R.; López, B.; de Mejía, M.; Mendoza, C.E.; Pepper, I.L. Confirmed detection of Cyclospora cayetanesis, Encephalitozoon intestinalis and Cryptosporidium parvum in water used for drinking. J. Water Health 2003, 1, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Graczyk, T.K.; McOliver, C.; Silbergeld, E.K.; Tamang, L.; Roberts, J.D. Risk of handling as a route of exposure to infectious waterborne Cryptosporidium parvum oocysts via Atlantic blue crabs (Callinectes sapidus). Appl. Environ. Microbiol. 2007, 73, 4069–4070. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, F.; Castro Hermida, J.A.; Fenoy, S.; Mezo, M.; González-Warleta, M.; del Aguila, C. Detection of microsporidia in drinking water, wastewater and recreational rivers. Water Res. 2011, 45, 4837–4843. [Google Scholar] [CrossRef] [PubMed]
- Galván, A.L.; Magnet, A.; Izquierdo, F.; Fenoy, S.; Rueda, C.; Vadillo, C.F.; Henriques-Gil, N.; del Aguila, C. Molecular characterization of human-pathogenic microsporidia and Cyclospora cayetanensis isolated from various water sources in Spain: A year-long longitudinal study. Appl. Environ. Microbiol. 2013, 79, 449–459. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Sunderland, D.; Awantang, G.N.; Mashinski, Y.; Lucy, F.E.; Graczyk, Z.; Chomicz, L.; Breysse, P.N. Relationships among bather density, levels of human waterborne pathogens, and fecal coliform counts in marine recreational beach water. Parasitol. Res. 2010, 106, 1103–1108. [Google Scholar] [CrossRef]
- Moss, J.A.; Snyder, R.A. Surveillance of Microsporidia and Protozoan Pathogens in Pensacola Florida: A One-year Study. J. Eukaryot Microbiol. 2019, 66, 617–624. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Conn, D.B.; Lucy, F.; Minchin, D.; Tamang, L.; Moura, L.N.S.; Da Silva, A.J. Human waterborne parasites in zebra mussels (Dreissena polymorha) from the Shannon River drainage area, Ireland. Parasitol. Res. 2004, 93, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Lucy, F.E.; Graczyk, T.K.; Tamang, L.; Miraflor, A.; Minchin, D. Biomonitoring of surface and coastal water for Cryptosporidium, Giardia, and human-virulent microsporidia using molluscan shellfish. Parasitol. Res. 2008, 103, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Marino, A.M.F.; Giunta, R.P.; Salvaggio, A.; Castello, A.; Alfonzetti, T.; Barbagallo, A.; Aparo, A.; Scalzo, F.; Reale, S.; Buffolano, W.; et al. Toxoplasma gondii in edible fishes captured in the Mediterranean basin. Zoonoses Public Health 2019, 66, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Certad, G.; Follet, J.; Gantois, N.; Hammouma-Ghelboun, O.; Guyot, K.; Benamrouz-Vanneste, S.; Fréalle, E.; Seesao, Y.; Delaire, B.; Creusy, C.; et al. Prevalence, molecular identification, and risk factors for Cryptosporidium infection in edible marine fish: A survey across sea areas surrounding France. Front. Microbiol. 2019, 10, 1037. [Google Scholar] [CrossRef]
- Gantois, N.; Lamot, A.; Seesao, Y.; Creusy, C.; Li, L.-L.; Monchy, S.; Benamrouz-Vanneste, S.; Karpouzopoulos, J.; Bourgain, J.-L.; Rault, C.; et al. First report on the prevalence and subtype distribution of Blastocystis sp. in edible marine fish and marine mammals: A large scale-study conducted in atlantic northeast and on the coasts of Northern France. Microorganisms 2020, 8, 460. [Google Scholar] [CrossRef]
- Yang, R.; Reid, A.; Lymbery, A.; Ryan, U. Identification of zoonotic Giardia genotypes in fish. Int. J. Parasitol. 2010, 40, 779–785. [Google Scholar] [CrossRef] [PubMed]
- APROMAR. Informe 2021. La Acuicultura en España. Available online: https://apromar.es/wp-content/uploads/2021/12/La-Acuicultura-en-Espana-2021.pdf (accessed on 14 December 2022).
- Polley, S.D.; Boadi, S.; Watson, J.; Curry, A.; Chiodini, P.L. Detection and species identification of microsporidial infections using SYBR Green real-time PCR. J. Med. Microbiol. 2011, 60, 459–466. [Google Scholar] [CrossRef]
- Andreu-Ballester, J.C.; Garcia-Ballesteros, C.; Amigo, V.; Ballester, F.; Gil-Borrás, R.; Catalán-Serra, I.; Magnet, A.; Fenoy, S.; del Aguila, C.; Ferrando-Marco, J.; et al. Microsporidia and Its Relation to Crohn Disease. A Retrospective Study. PLoS ONE 2013, 8, e62107. [Google Scholar] [CrossRef]
- Park, E.; Poulin, R. Revisiting the phylogeny of microsporidia. Int. J. Parasitol. 2021, 51, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Bojko, J.; Reinke, A.W.; Stentiford, G.D.; Williams, B.; Rogers, M.S.J.; Bass, D. Microsporidia: A new taxonomic, evolutionary, and ecological synthesis. Trends Parasitol. 2022, 38, 642–659. [Google Scholar] [CrossRef] [PubMed]
- Thurston-Enriquez, J.A.; Watt, P.; Dowd, S.E.; Enriquez, R.; Pepper, I.L.; Gerba, C.P. Detection of protozoan parasites and microsporidia in irrigation waters used for crop production. J. Food Prot. 2002, 65, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Santin-Duran, M. Epidemiology of microsporidia in human infections. In Microsporidia: Pathogens of Opportunity, 1st ed.; Weiss, L.M., Becnel, J.J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; pp. 135–164. [Google Scholar]
- Garcia, L.S. Laboratory identification of the microsporidia. J. Clin. Microbiol. 2002, 40, 1892–1901. [Google Scholar] [CrossRef] [PubMed]
- Matos, O.; Lobo, M.L.; Xiao, L. Epidemiology of Enterocytozoon bieneusi infection in humans. J. Parasitol. Res. 2012, 2012, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Santín, M.; Fayer, R. Microsporidiosis: Enterocytozoon bieneusi in domesticated and wild animals. Res. Vet. Sci. 2011, 90, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Haro, M.; Izquierdo, F.; Henriques-Gil, N.; Andrés, I.; Alonso, F.; Fenoy, S.; del Águila, C. First detection and genotyping of human-associated microsporidia in pigeons from urban parks. Appl. Environ. Microbiol. 2005, 71, 3153–3157. [Google Scholar] [CrossRef]
- Li, W.; Feng, Y.; Santin, M. Host Specificity of Enterocytozoon bieneusi and Public Health Implications. Trends Parasitol. 2019, 35, 436–451. [Google Scholar] [CrossRef]
- Izquierdo, F.; Ollero, D.; Magnet, A.; Galván-Díaz, A.L.; Llorens, S.; Vaccaro, L.; Hurtado-Marcos, C.; Valdivieso, E.; Miró, G.; Hernández, L.; et al. Microsporidia as a Potential Threat to the Iberian Lynx (Lynx pardinus). Animals 2022, 12, 2507. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation. 2022. Available online: https://www.fao.org/publications/home/fao-flagship-publications/the-state-of-world-fisheries-and-aquaculture/2022/en (accessed on 15 May 2023).
- Skov, J.; Kania, P.W.; Olsen, M.M.; Lauridsen, J.H.; Buchmann, K. Nematode infections of maricultured and wild fishes in Danish waters: A comparative study. Aquaculture 2009, 298, 24–28. [Google Scholar] [CrossRef]
- Cheng, H.W.A.; Lucy, F.E.; Graczyk, T.K.; Broaders, M.A.; Mastitsky, S.E. Municipal wastewater treatment plants as removal systems and environmental sources of human-virulent microsporidian spores. Parasitol. Res. 2011, 109, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Moratal, S.; Dea-Ayuela, M.A.; Martí-Marco, A.; Puigcercós, S.; Marco-Hirs, N.M.; Doménech, C.; Corcuera, E.; Cardells, J.; Lizana, V.; López-Ramon, J. Molecular Characterization of Cryptosporidium spp. in Cultivated and Wild Marine Fishes from Western Mediterranean with the First Detection of Zoonotic Cryptosporidium ubiquitum. Animals 2022, 12, 1052. [Google Scholar] [CrossRef] [PubMed]
- Chintagari, S.; Hazard, N.; Edwards, G.; Jadeja, R.; Janes, M. Risks Associated with Fish and Seafood. Microbiol. Spectrum 2017, 5, PFS-0013-2016. [Google Scholar] [CrossRef] [PubMed]
- Broglia, A.; Kapel, C. Changing dietary habits in a changing world: Emerging drivers for the transmission of foodborne parasitic zoonoses. Vet. Parasitol. 2011, 182, 2–13. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Group | Presence (Number of Positives/Total) | |||
|---|---|---|---|---|
| Enterocytozoonidae | E. intestinalis/hellem | E. cuniculi | Total | |
| Farmed fish | 2.90% (4/138) | 1.45% (2/138) | 0% (0/138) | 4.35% (6/138) |
| Wild fish | 18.58% (21/113) | 7.96% (9/113) | 0% (0/113) | 26.55% (30/113) |
| Total | 9.96% (25/251) | 4.38% (11/251) | 0% (0/251) | 14.34% (36/251) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moratal, S.; Magnet, A.; Izquierdo, F.; del Águila, C.; López-Ramon, J.; Dea-Ayuela, M.A. Microsporidia in Commercially Harvested Marine Fish: A Potential Health Risk for Consumers. Animals 2023, 13, 2673. https://doi.org/10.3390/ani13162673
Moratal S, Magnet A, Izquierdo F, del Águila C, López-Ramon J, Dea-Ayuela MA. Microsporidia in Commercially Harvested Marine Fish: A Potential Health Risk for Consumers. Animals. 2023; 13(16):2673. https://doi.org/10.3390/ani13162673
Chicago/Turabian StyleMoratal, Samantha, Angela Magnet, Fernando Izquierdo, Carmen del Águila, Jordi López-Ramon, and María Auxiliadora Dea-Ayuela. 2023. "Microsporidia in Commercially Harvested Marine Fish: A Potential Health Risk for Consumers" Animals 13, no. 16: 2673. https://doi.org/10.3390/ani13162673