
Study on the Adaptive Regulation of Light on the Stress Response of Mandarin Fish (Siniperca chuatsi) with Re-Feeding after Starvation
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
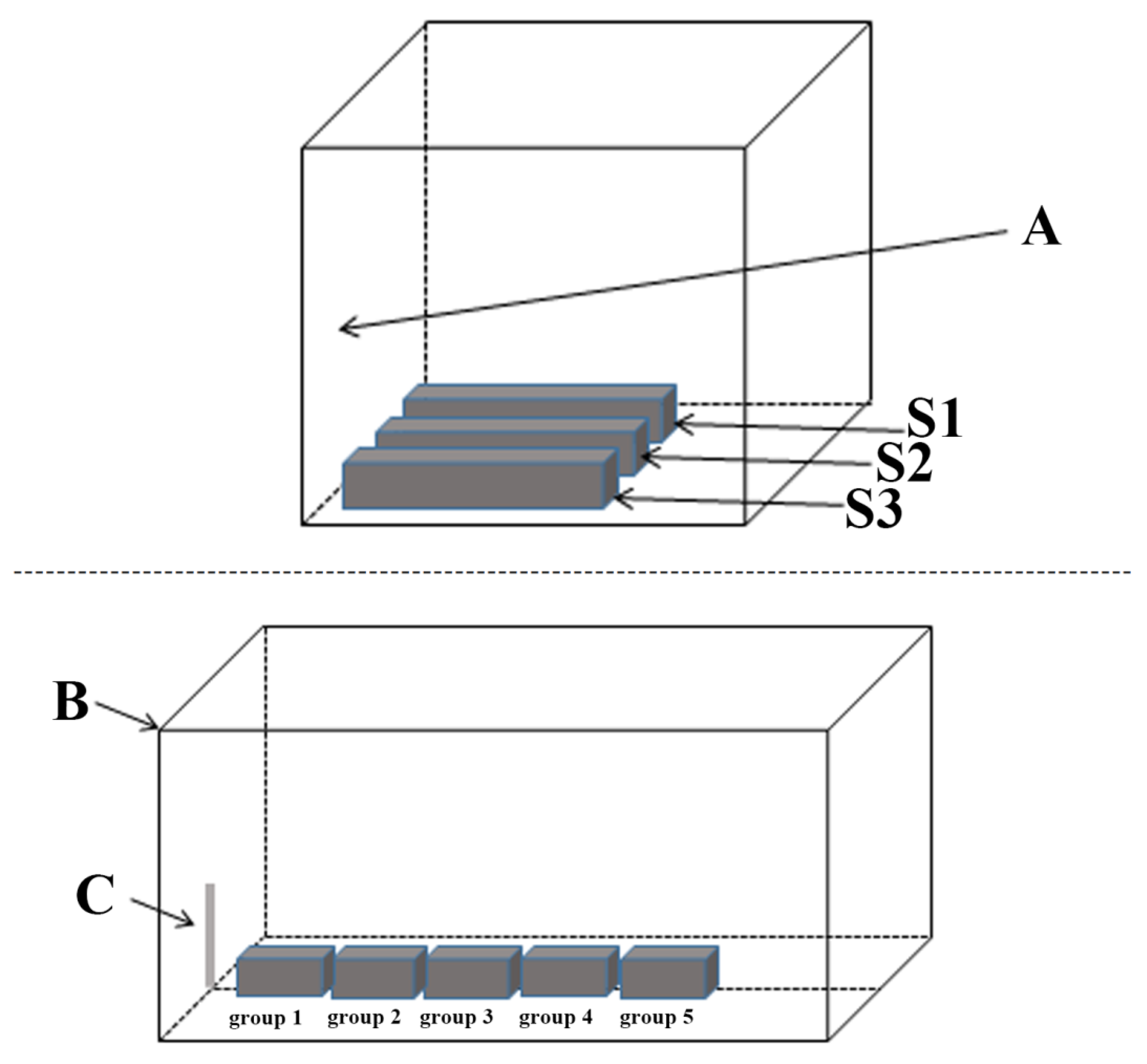
2.2. Source of Experimental Organisms and Experimental Design
2.3. Tissue Sample Collection
2.4. Enzymatic Activity Analysis
2.5. Data Analysis
3. Results
3.1. Total Protein Content Analysis
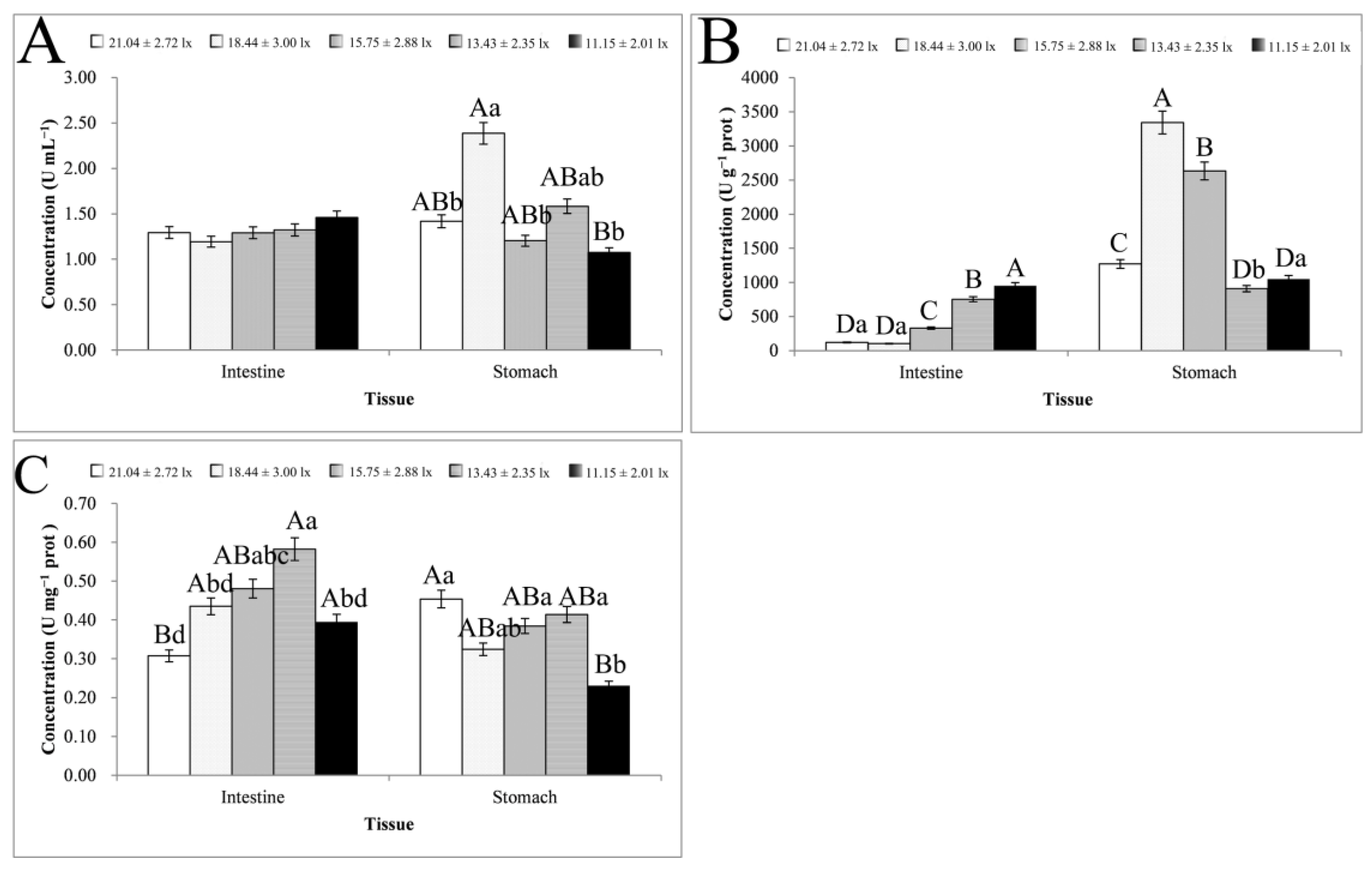
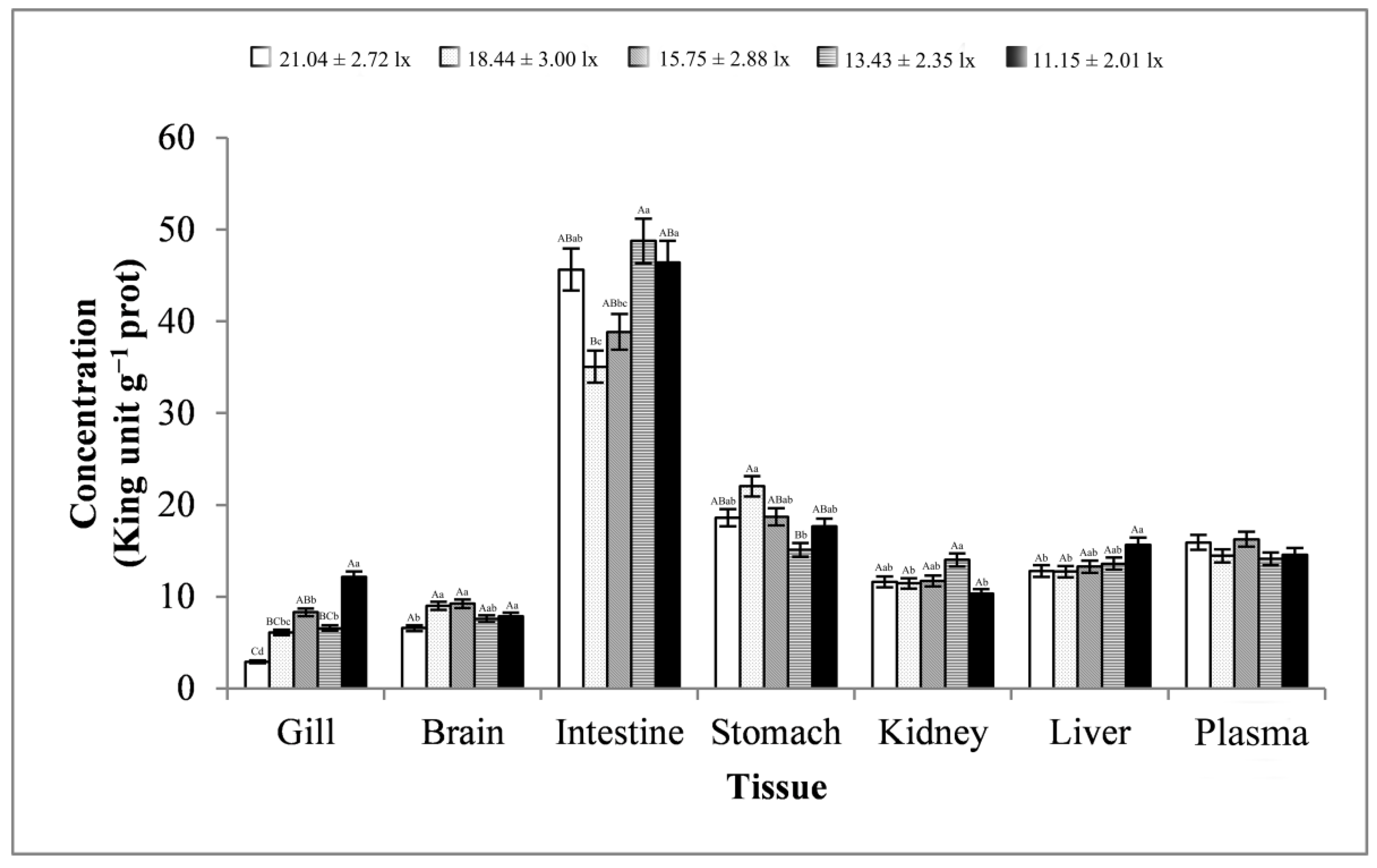
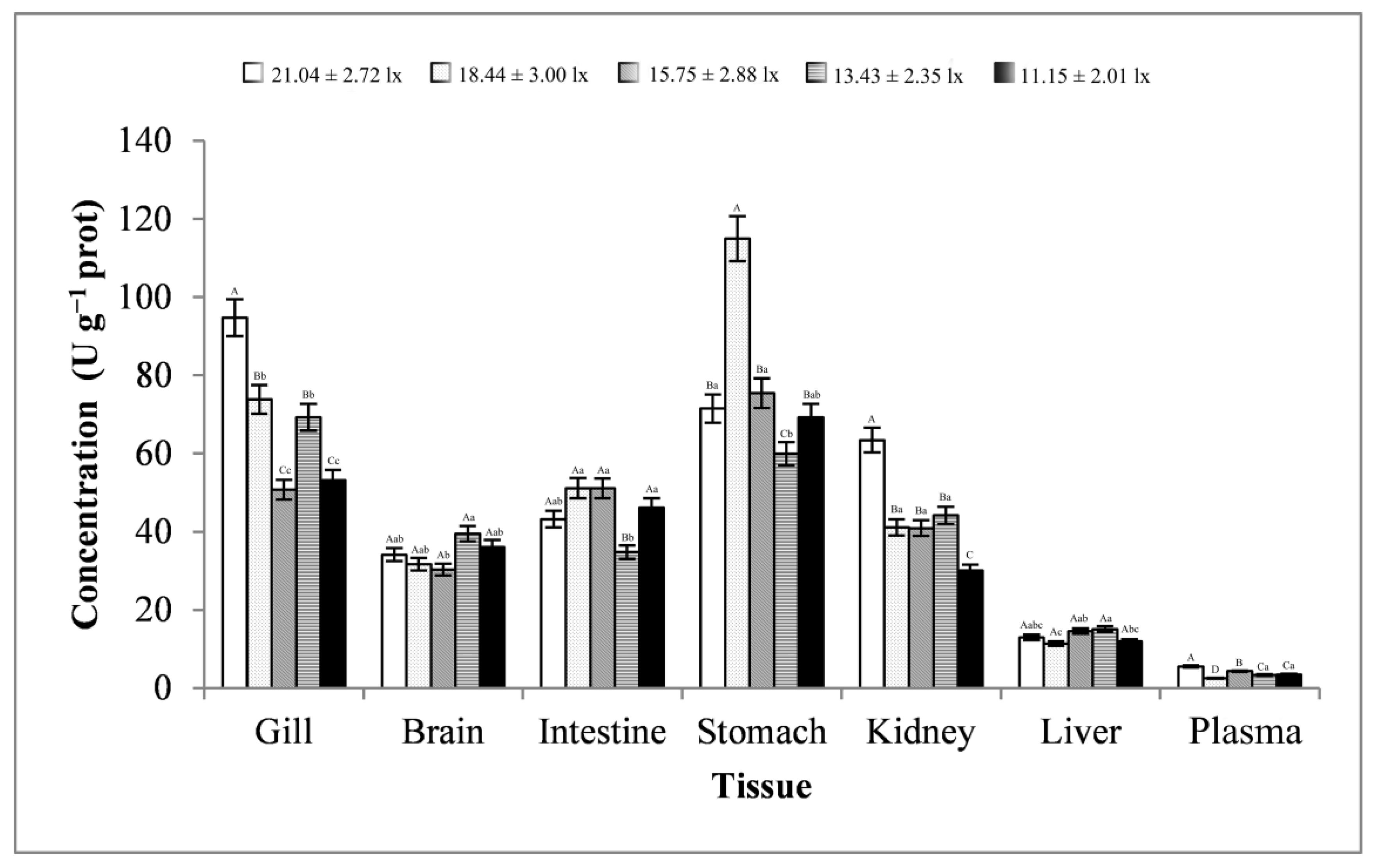
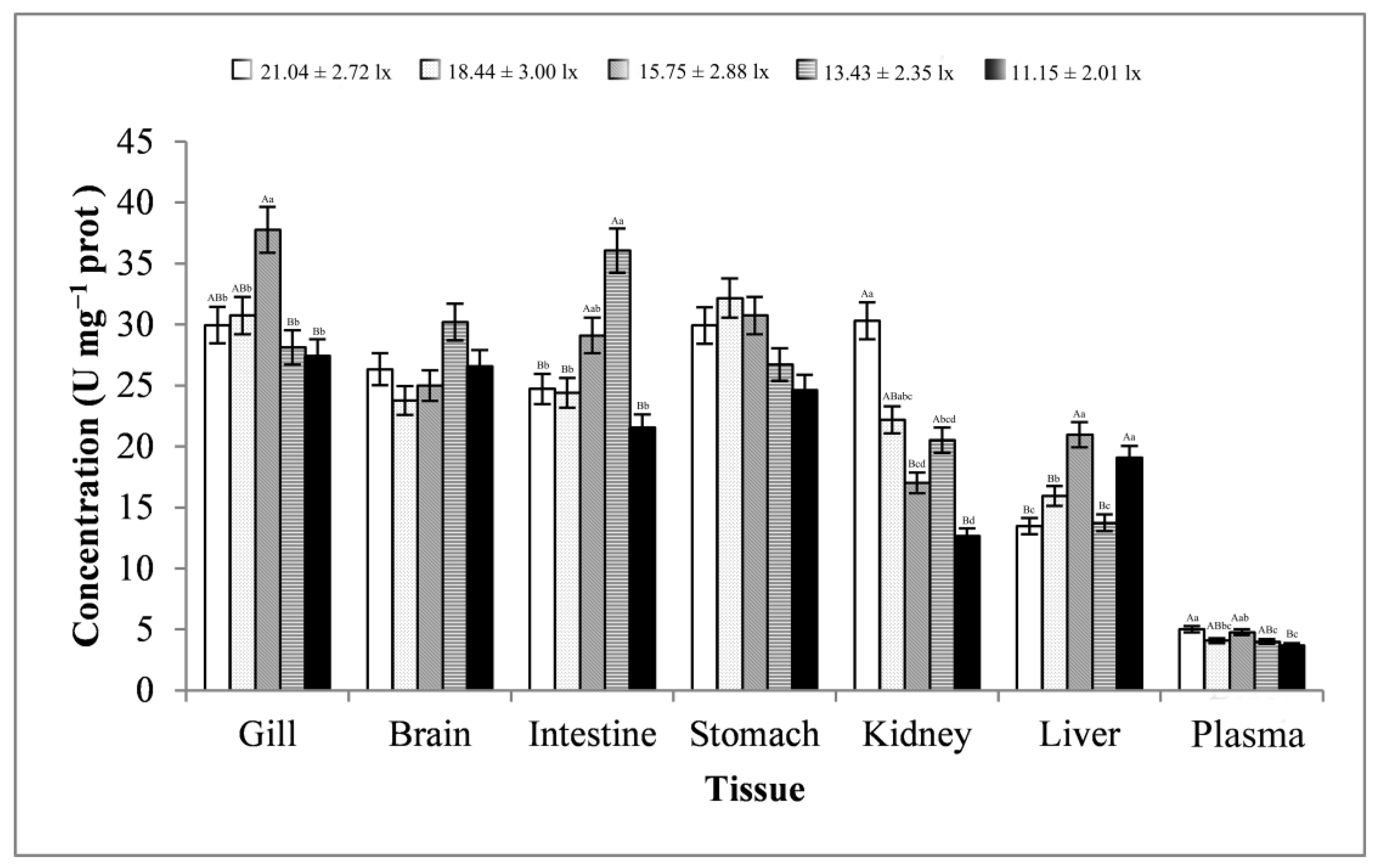
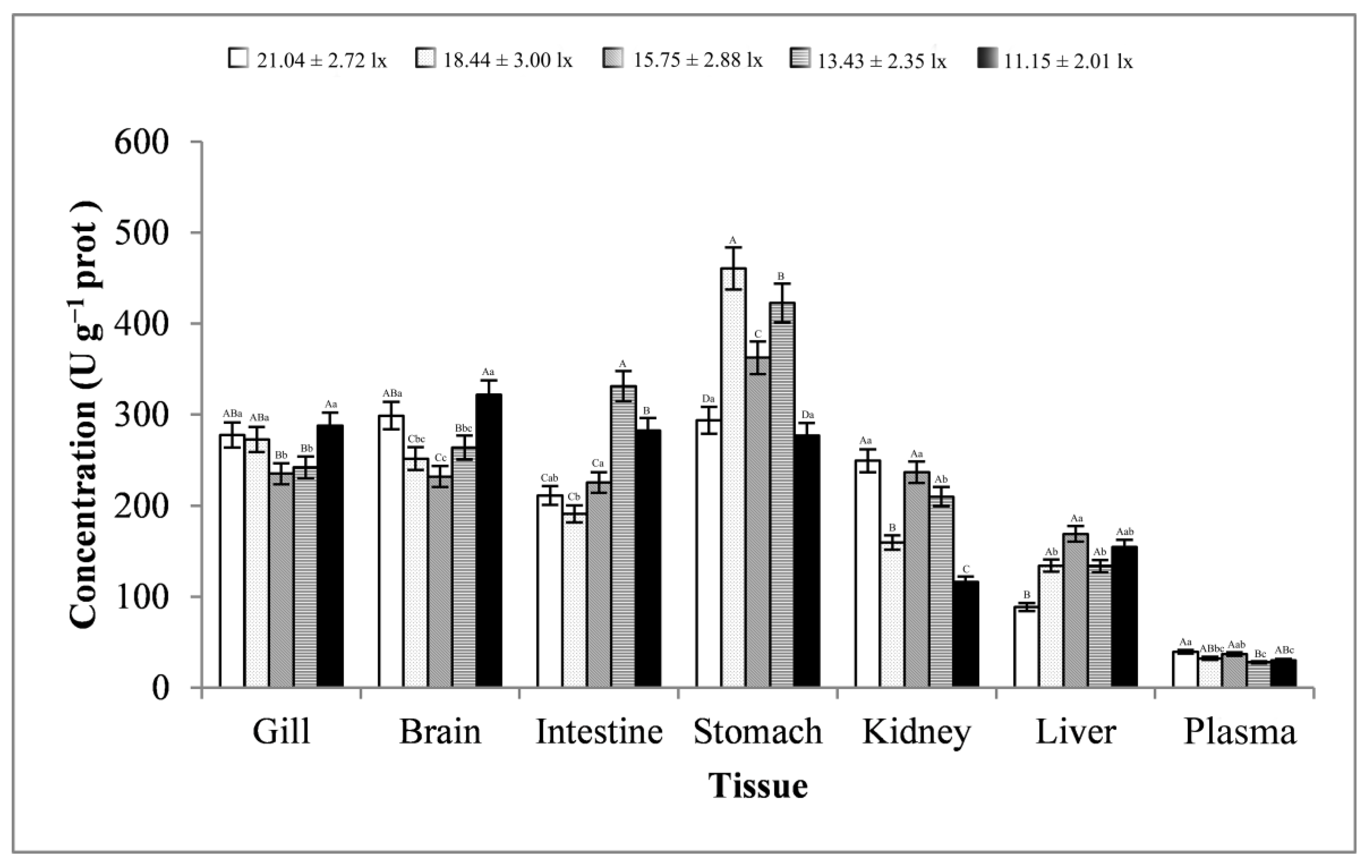
3.2. Digestive Enzyme Activity Analysis
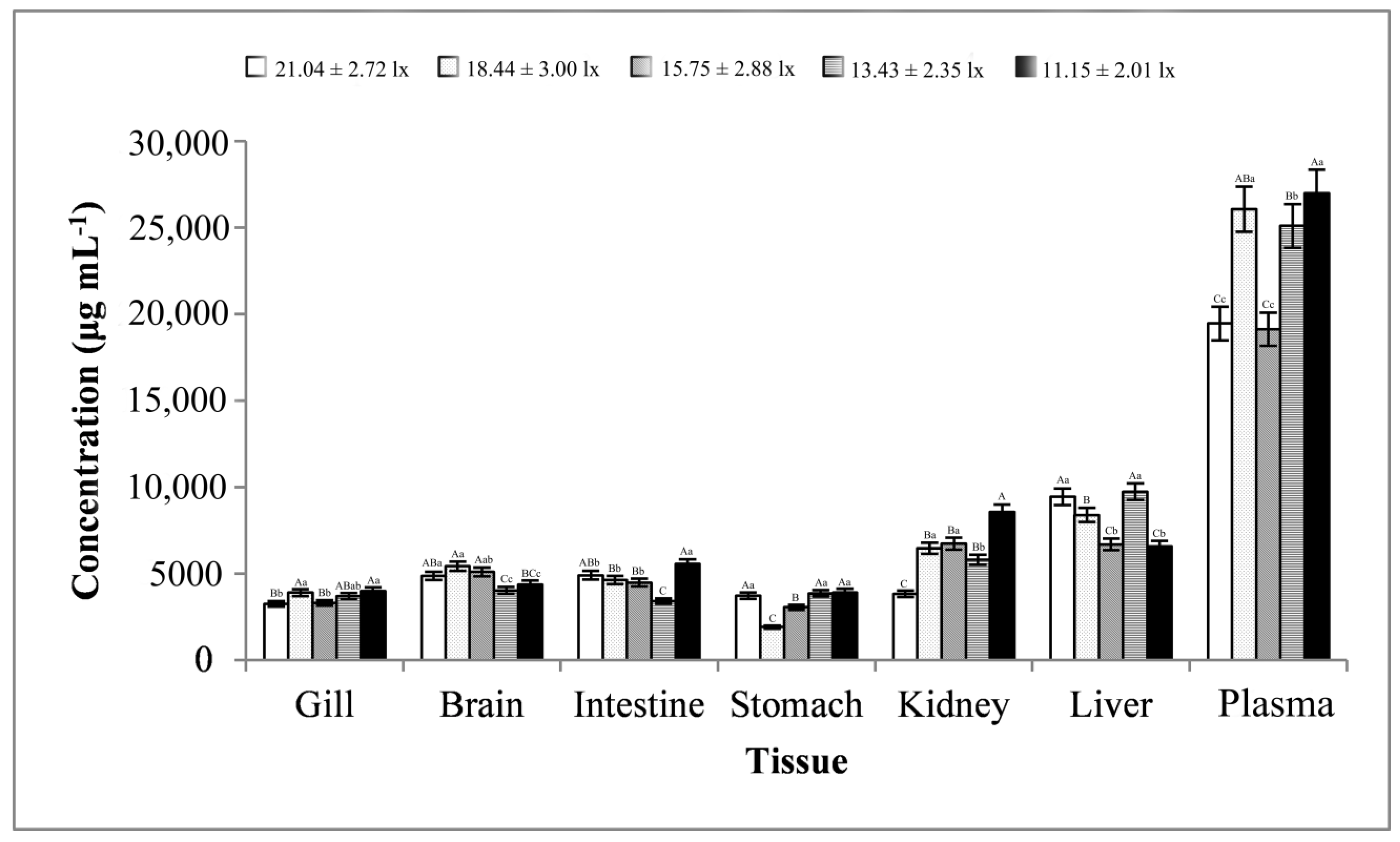
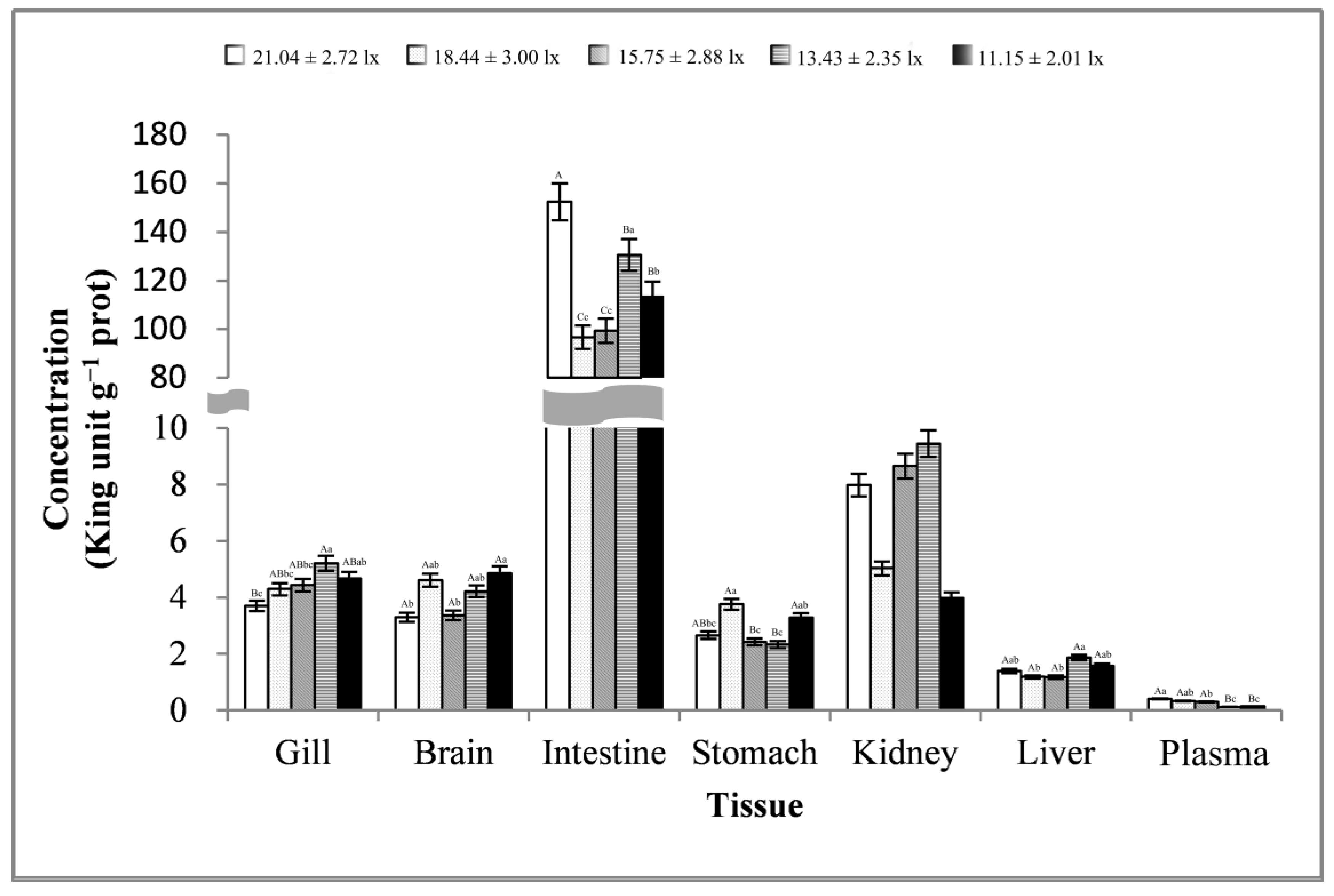
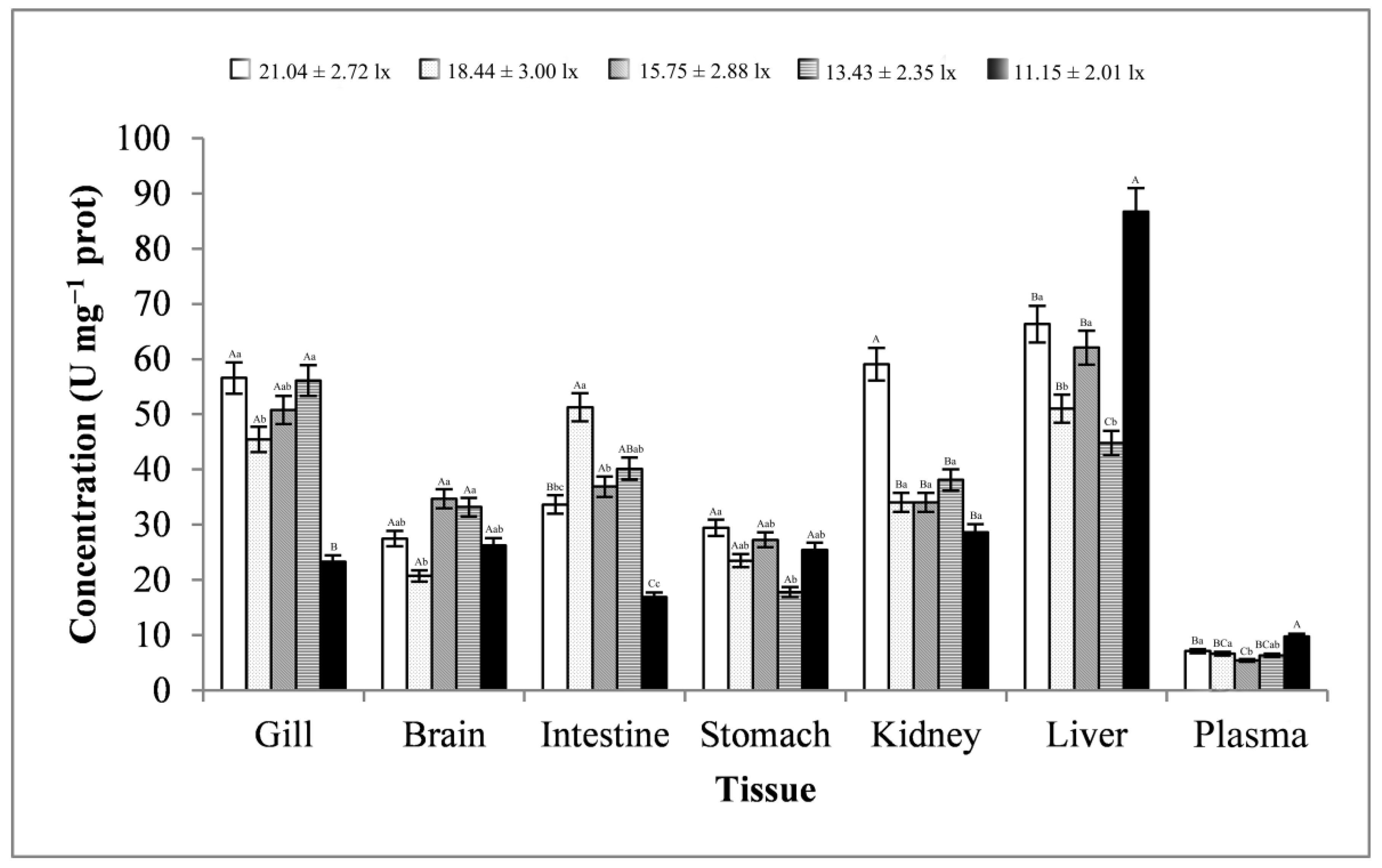
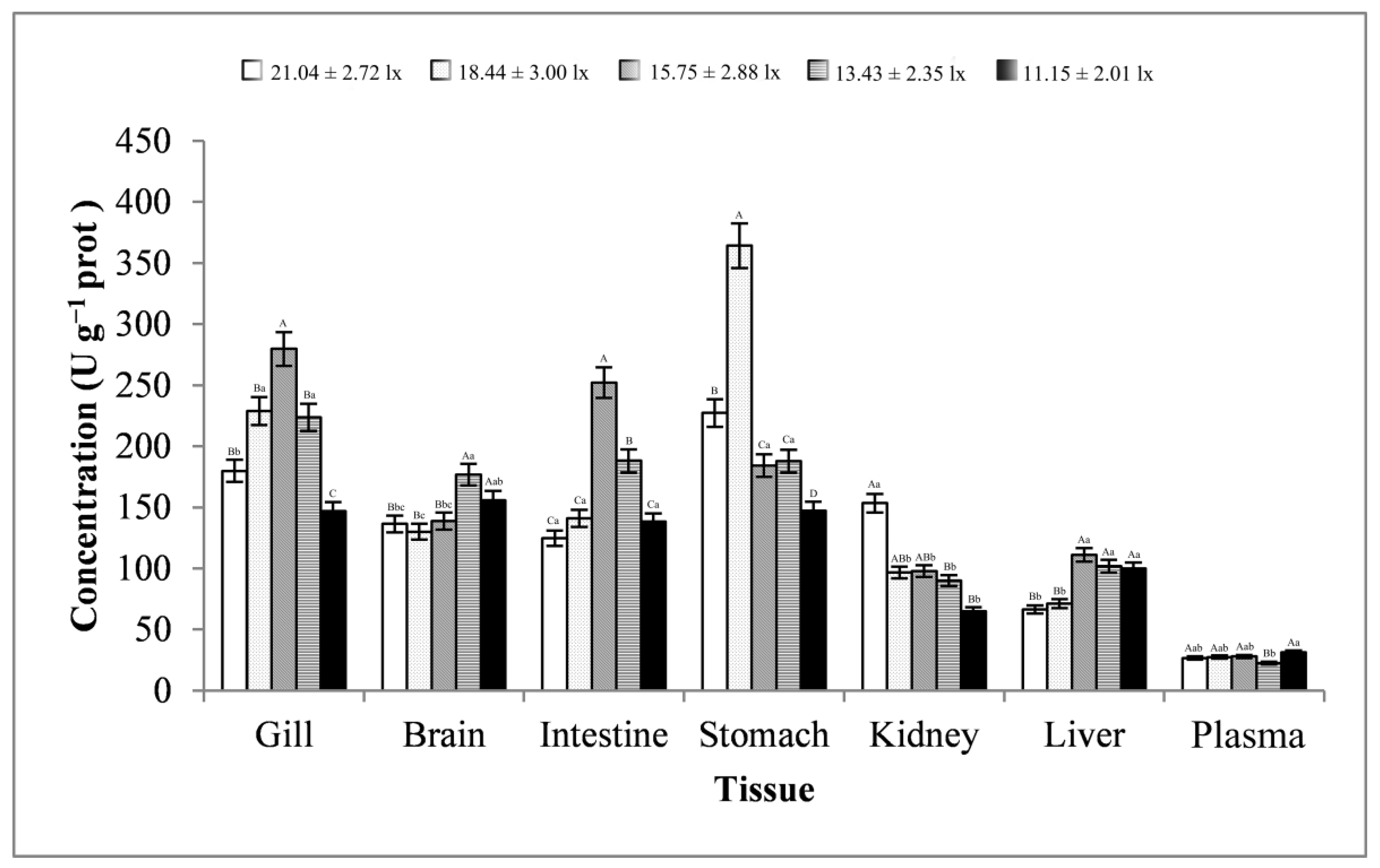
3.3. Stress-Related Enzyme Activity Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, K.; Zhang, Z.; Li, J.; Xie, S.; Shi, L.J.; He, Y.H.; Liang, X.F.; Zhu, Q.S.; He, S. Different regulation of branched-chain amino acid on food intake by TOR signaling in Chinese perch (Siniperca chuatsi). Aquaculture 2021, 530, 735792. [Google Scholar] [CrossRef]
- Ren, Y.; Xiong, M.; Yu, J.; Li, W.; Li, B.; Liu, J.; Zhang, T. Effects of artificial submersed vegetation on consumption and growth of mandarin fish Siniperca chuatsi (Basilewsky) foraging on live prey. J. Freshw. Ecol. 2019, 34, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liang, X.F.; He, S.; Feng, H.; Li, L. Dietary supplementation of exogenous probiotics affects growth performance and gut health by regulating gut microbiota in Chinese Perch (Siniperca chuatsi). Aquaculture 2022, 547, 737405. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, K.; Liang, X.F.; Li, J.; Tang, S.; Zhang, Y.; Cai, W.; Xiao, Q.; Zhang, Q. Effects of early low temperature exposure on the growth, glycolipid metabolism and growth hormone (gh) gene methylation in the late stage of Chinese perch (Siniperca chuatsi). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2022, 259, 110705. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z. Food ingestion, prey selectivity, feeding incidence, and performance of yellowtail kingfish Seriola lalandi larvae under constant and varying temperatures. Aquac. Int. 2014, 22, 1317–1330. [Google Scholar] [CrossRef]
- Volkoff, H.; Rønnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef]
- Wang, F.; Cheng, G.; Dong, J.; Sun, C.; Wang, M.; Lu, M.; Ye, X. Construction of Growth Model for Mandarin Fish (Siniperca chuatsi) “Guangqing No.1” Fed with Compound Diet and Cultivated in Pond. Guangdong Agricultural Sciences 2022. Available online: https://kns.cnki.net/kcms/detail/44.1267.s.20220614.1143.006.html (accessed on 3 May 2023).
- Jang, Y.K. On the biology of mandarin fish, Siniperca chuatsi of Liang-Tze lake. Acta Hydrobiol. Sin. 1959, 375–385. (In Chinese) [Google Scholar]
- Liu, X.; Bin, S.; Wang, K.; Chen, D.; Cheng, J.; Zhang, J.; Chu, W. Artificial Propagation and Embryonic Development Observation of Mandarin Fish. J. Guangxi Norm. Univ. 2013, 31, 100–106. [Google Scholar]
- Juell, J.E.; Fosseidengen, J.E. Use of artificial light to control swimming depth and fish density of Atlantic salmon (Salmo salar) in production cages. Aquaculture 2004, 233, 269–282. [Google Scholar] [CrossRef]
- Juell, J.E.; Oppedal, F.; Boxaspen, K.; Taranger, G.L. Submerged light increases swimming depth and reduces fish density of Atlantic salmon Salmo salar L. in production cages. Aquac. Res. 2003, 34, 469–478. [Google Scholar] [CrossRef]
- Brüning, A.; Hölker, F.; Franke, S.; Preuer, T.; Kloas, W. Spotlight on fish: Light pollution affects circadian rhythms of European perch but does not cause stress. Sci. Total Environ. 2015, 511, 516–522. [Google Scholar] [CrossRef]
- Zan-Bar, T.; Bartoov, B.; Segal, R.; Yehuda, R.; Lavi, R.; Lubart, R.; Avtalion, R.R. Influence of visible light and ultraviolet irradiation on motility and fertility of mammalian and fish sperm. Photomed. Laser Surg. 2005, 23, 549–555. [Google Scholar] [CrossRef]
- Brüning, A.; Hölker, F.; Wolter, C. Artificial light at night: Implications for early life stages development in four temperate freshwater fish species. Aquat. Sci. 2011, 73, 143–152. [Google Scholar] [CrossRef]
- Volpato, G.L.; Bovi, T.S.; de Freitas, R.H.; da Silva, D.F.; Delicio, H.C.; Giaquinto, P.C.; Barreto, R.E. Red light stimulates feeding motivation in fish but does not improve growth. PLoS ONE 2013, 8, e59134. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Fu, H.C.; Wang, Y.Y.; Huang, H.Z. Genome-wide association study reveals growth-related SNPs and candidate genes in mandarin fish (Siniperca chuatsi). Aquaculture 2022, 550, 737879. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Liang, X.F.; He, S.; Tang, S.L.; Peng, D. The potential use of Artemia for larval rearing of mandarin fish (Siniperca chuatsi). Aquac. Rep. 2022, 25, 101216. [Google Scholar] [CrossRef]
- Feng, H.; Peng, D.; Liang, X.F.; Chai, F.; Tang, S.; Li, J. Effect of dietary hydroxyproline supplementation on Chinese perch (Siniperca chuatsi) fed with fish meal partially replaced by fermented soybean meal. Aquaculture 2022, 547, 737454. [Google Scholar] [CrossRef]
- Peng, D.; Liang, X.F.; Chai, F.; Feng, H.; Li, J.; Tang, S.; Lu, K.; Zhang, Q. Effects of dietary carbohydrate to lipid ratios on growth, biochemical indicators, lipid metabolism, and appetite in Chinese perch (Siniperca chuatsi). Fish Physiol. Biochem. 2022, 48, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.M.; Huang, H.Z.; Zhou, Y.; Wang, Y.Y.; Fu, H.C.; Li, Z.; Fu, X.Z.; Li, N.Q. Characterization of mandarin fish (Siniperca chuatsi) IL-6 and IL-6 signal transducer and the association between their SNPs and resistance to ISKNV disease. Fish Shellfish. Immunol. 2021, 113, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gu, Y.; Hou, X.; Wang, H.; Wang, P.; Xu, P.; Zeng, L.; Zhou, L.; Li, G. Molecular characterization, tissue distribution, and expression regulation from fasting and re-feeding of two growth hormone receptors in mandarin fish Siniperca chuatsi. Fish. Sci. 2016, 82, 155–169. [Google Scholar] [CrossRef]
- Chen, X.; Wang, G.; Lu, X.; Xu, P.; Zeng, S.; Chen, Z.; Zhu, Q.; Li, G. Molecular characterization and expression profiles of two insulin-like growth factor 1 receptors during fasting and re-feeding in Siniperca chuatsi. Fish. Sci. 2019, 85, 349–360. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, D.; Hu, Y.; Wu, P.; Wang, K.; Zhang, J.; Chu, W.; Zhang, J. The microRNA signature in response to nutrient restriction and refeeding in skeletal muscle of Chinese perch (Siniperca chuatsi). Mar. Biotechnol. 2015, 17, 180–189. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Chen, D.; Zhu, X.; Liu, Z.; Zhang, J.; Chu, W.; Bin, S. Effect of starvating and refeeding on relative expression of FSRP-1, FSRP-3 and PepT1 gene in mandarin fish (Siniperca chuatsi). J. Guangxi Norm. Univ. 2016, 34, 144–149. [Google Scholar] [CrossRef]
- Shi, L.; Li, J.; Liang, X.F.; He, S.; Dou, Y.; Peng, J.; Cai, W.; Liang, H. Memory regulation in feeding habit transformation to dead prey fish of Chinese perch (Siniperca chuatsi). Fish Physiol. Biochem. 2021, 47, 1893–1907. [Google Scholar] [CrossRef]
- Kari, Z.A.; Kabir, M.A.; Dawood, M.A.; Razab, M.K.A.A.; Ariff, N.S.N.A.; Sarkar, T.; Pati, S.; Edinur, H.A.; Mat, K.; Ismail, T.A.; et al. Effect of fish meal substitution with fermented soy pulp on growth performance, digestive enzyme, amino acid profile, and immune-related gene expression of African catfish (Clarias gariepinus). Aquaculture 2022, 546, 737418. [Google Scholar] [CrossRef]
- Dawood, M.A. Nutritional immunity of fish intestines: Important insights for sustainable aquaculture. Rev. Aquac. 2021, 13, 642–663. [Google Scholar] [CrossRef]
- Jiménez, K.; Solano, K.; Scholz, C.; Redondo-López, S.; Mena, F. Early Toxic Effects in a Central American Native Fish (Parachromis dovii) Exposed to Chlorpyrifos and Difenoconazole. Environ. Toxicol. Chem. 2021, 40, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Bhushan, S.; Gupta, S.K.; Kumar, P.; Chandan, N.K.; Singh, D.K.; Kumar, P. Metal determination and biochemical status of marine fishes facilitate the biomonitoring of marine pollution. Mar. Pollut. Bull. 2021, 170, 112682. [Google Scholar] [CrossRef]
- Qian, D.; Yang, X.; Xu, C.; Chen, C.; Jia, Y.; Gu, Z.; Li, E. Growth and health status of the red claw crayfish, Cherax quadricarinatus, fed diets with four typical plant protein sources as a replacement for fish meal. Aquac. Nutr. 2021, 27, 795–806. [Google Scholar] [CrossRef]
- Yun, H.; Park, G.; Ok, I.; Katya, K.; Hung, S.; Bai, S.C. Determination of the dietary lysine requirement by measuring plasma free lysine concentrations in rainbow trout Oncorhynchus mykiss after dorsal aorta cannulation. Fish. Aquat. Sci. 2016, 19, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Rahimnejad, S.; Wang, Y.R.; Lu, K.; Song, K.; Wang, L.; Mai, K. Substituting fish meal with soybean meal in diets for Japanese seabass (Lateolabrax japonicus): Effects on growth, digestive enzymes activity, gut histology, and expression of gut inflammatory and transporter genes. Aquaculture 2018, 483, 173–182. [Google Scholar] [CrossRef]
- Duman, M.; Saticioglu, I.B.; Suzer, B.; Altun, S. Practices for drawing blood samples from teleost fish. N. Am. J. Aquac. 2019, 81, 119–125. [Google Scholar] [CrossRef]
- Wei, K.; Zhang, G.; Zhang, H. Studies on the phototactic characteristics of mandarin fish (Siniperca chuatsi) during different development stages. J. Huazhong Agric. Univ. 2001, 20, 164–168. [Google Scholar] [CrossRef]
- Liang, X.F.; Lin, X.; Li, S.; Liu, J.K. Impact of environmental and innate factors on the food habit of Chinese perch Siniperca chuatsi (Basilewsky) (Percichthyidae). Aquac. Res. 2008, 39, 150–157. [Google Scholar] [CrossRef]
- Xiufeng, L.; Daoming, H.; Hanyun, Y. Effect of light on young Siniperca kneri Garman’s ingested amount. J. Hunan Agric. Univ. 2005, 31, 187–190. [Google Scholar]
- Shen, W.X. Biological characteristics and artificial propagation fry technology of Siniperca chuatsi. Shanghai Agric. Sci. Technol. 2018, 70–71. (In Chinese) [Google Scholar]
- Lie, K.K.; Tørresen, O.K.; Solbakken, M.H.; Rønnestad, I.; Tooming-Klunderud, A.; Nederbragt, A.J.; Jentoft, S.; Sæle, Ø. Loss of stomach, loss of appetite? Sequencing of the ballan wrasse (Labrus bergylta) genome and intestinal transcriptomic profiling illuminate the evolution of loss of stomach function in fish. BMC Genom. 2018, 19, 186. [Google Scholar] [CrossRef]
- Noureldin, S.M.; Diab, A.M.; Salah, A.S.; Mohamed, R.A. Effect of different monochromatic LED light colors on growth performance, behavior, immune-physiological responses of gold fish, Carassius auratus. Aquaculture 2021, 538, 736532. [Google Scholar] [CrossRef]
- Oliveira, C.C.; Aparício, R.; Blanco-Vives, B.; Chereguini, O.; Martín, I.; Javier Sánchez-Vazquez, F. Endocrine (plasma cortisol and glucose) and behavioral (locomotor and self-feeding activity) circadian rhythms in Senegalese sole (Solea senegalensis Kaup 1858) exposed to light/dark cycles or constant light. Fish Physiol. Biochem. 2013, 39, 479–487. [Google Scholar] [CrossRef]
- Hong, J.; Chen, X.; Liu, S.; Fu, Z.; Han, M.; Wang, Y.; Gu, Z.; Ma, Z. Impact of fish density on water quality and physiological response of golden pompano (Trachinotus ovatus) flingerlings during transportation. Aquaculture 2019, 507, 260–265. [Google Scholar] [CrossRef]
- Sun, A.; Zhu, H.; Wang, X.; Hu, Q.; Tian, Z.; Hu, H. Molecular characterization of manganese superoxide dismutase (MnSOD) from sterlet Acipenser ruthenus and its responses to Aeromonas hydrophila challenge and hypoxia stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 234, 68–76. [Google Scholar] [CrossRef]
- Saetan, W.; Tian, C.; Yu, J.; Lin, X.; He, F.; Huang, Y.; Shi, H.; Zhang, Y.; Li, G. Comparative transcriptome analysis of gill tissue in response to hypoxia in silver sillago (Sillago sihama). Animals 2020, 10, 628. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhang, H.; Guo, M.; Fang, D.; Mei, J.; Xie, J. Analysis of Acute Nitrite Exposure on Physiological Stress Response, Oxidative Stress, Gill Tissue Morphology and Immune Response of Large Yellow Croaker (Larimichthys crocea). Animals 2022, 12, 1791. [Google Scholar] [CrossRef]
- Lallès, J.P. Intestinal alkaline phosphatase in the gastrointestinal tract of fish: Biology, ontogeny, and environmental and nutritional modulation. Rev. Aquac. 2020, 12, 555–581. [Google Scholar] [CrossRef]
- Black, E.C. The transport of oxygen by the blood of freshwater fish. Biol. Bull. 1940, 79, 215–229. [Google Scholar] [CrossRef]
- Wells, R.M.; Baldwin, J.; Seymour, R.S.; Weber, R.E. Blood oxygen transport and hemoglobin function in three tropical fish species from northern Australian freshwater billabongs. Fish Physiol. Biochem. 1997, 16, 247–258. [Google Scholar] [CrossRef]
- Ryu, H.S.; Song, J.A.; Park, H.S.; Choi, Y.J.; Choi, C.Y. Physiological and oxidative stress response of goldfish Carassius auratus induced by a light dimming system. Fish Physiol. Biochem. 2020, 46, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.Y.; Zhang, D.D.; Xu, C.; Wang, F.; Liu, W.B. Effects of light intensity on growth, immune responses, antioxidant capability and disease resistance of juvenile blunt snout bream Megalobrama amblycephala. Fish Shellfish. Immunol. 2015, 47, 674–680. [Google Scholar] [CrossRef]
- Lin, C.; Dai, H.; Shi, X.; Deng, Z.D.; Mao, J.; Luo, J.; Huang, W.; Xu, J.; Zhang, N.; Sun, S. Investigating feasible light configurations for fish restoration: An ethological insight. Fish. Res. 2021, 234, 105807. [Google Scholar] [CrossRef]
- Han, D.; Xie, S.; Lei, W.; Zhu, X.; Yang, Y. Effect of light intensity on growth, survival and skin color of juvenile Chinese longsnout catfish (Leiocassis longirostris Günther). Aquaculture 2005, 248, 299–306. [Google Scholar] [CrossRef]
- Shen, Y.; Li, H.; Zhao, J.; Tang, S.; Zhao, Y.; Bi, Y.; Chen, X. The digestive system of mandarin fish (Siniperca chuatsi) can adapt to domestication by feeding with artificial diet. Aquaculture 2021, 538, 736546. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Li, Q.; Huang, Z.; Zhang, L.; Mou, C.; Zhao, Z.; Zhao, H.; Du, J.; Yang, X.; Liang, X.; et al. Study on the Adaptive Regulation of Light on the Stress Response of Mandarin Fish (Siniperca chuatsi) with Re-Feeding after Starvation. Animals 2023, 13, 2610. https://doi.org/10.3390/ani13162610
Zhou J, Li Q, Huang Z, Zhang L, Mou C, Zhao Z, Zhao H, Du J, Yang X, Liang X, et al. Study on the Adaptive Regulation of Light on the Stress Response of Mandarin Fish (Siniperca chuatsi) with Re-Feeding after Starvation. Animals. 2023; 13(16):2610. https://doi.org/10.3390/ani13162610
Chicago/Turabian StyleZhou, Jian, Qiang Li, Zhipeng Huang, Lu Zhang, Chengyan Mou, Zhongmeng Zhao, Han Zhao, Jun Du, Xiaojun Yang, Xufang Liang, and et al. 2023. "Study on the Adaptive Regulation of Light on the Stress Response of Mandarin Fish (Siniperca chuatsi) with Re-Feeding after Starvation" Animals 13, no. 16: 2610. https://doi.org/10.3390/ani13162610