
Molecular Detection and Epidemiology of Potentially Zoonotic Cryptosporidium spp. and Giardia duodenalis in Wild Boar (Sus scrofa) from Eastern Spain
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
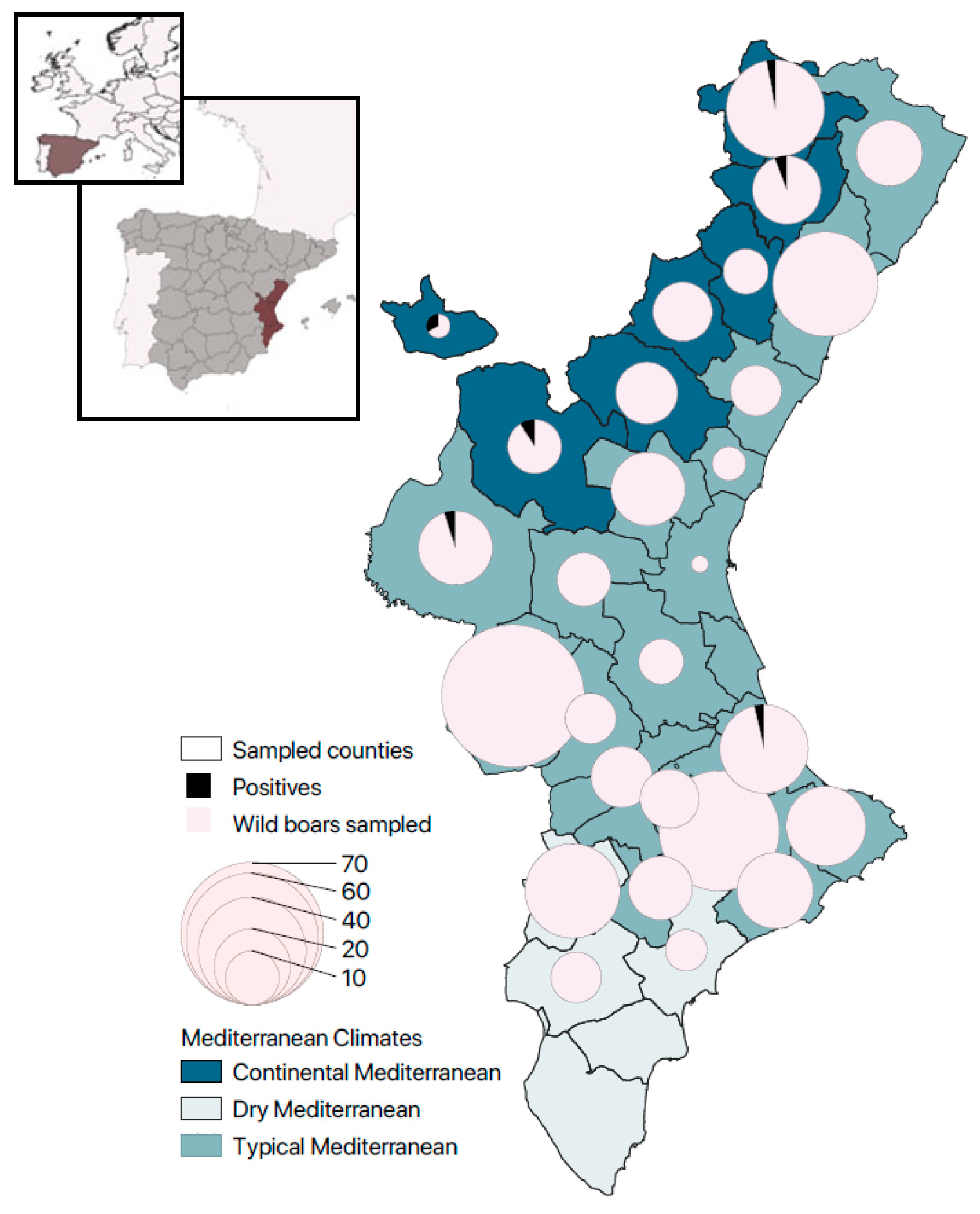
2.1. Study Area and Wild Boar Sampling
2.2. Environmental and Population Data Collection
2.2.1. Climate, Rainfall Regime, and Seasonality
2.2.2. Land Use
2.2.3. Wild Boar Population
2.2.4. Human Population
2.3. DNA Extraction
2.4. Molecular Detection and Characterisation of Giardia duodenalis and Cryptosporidium spp.
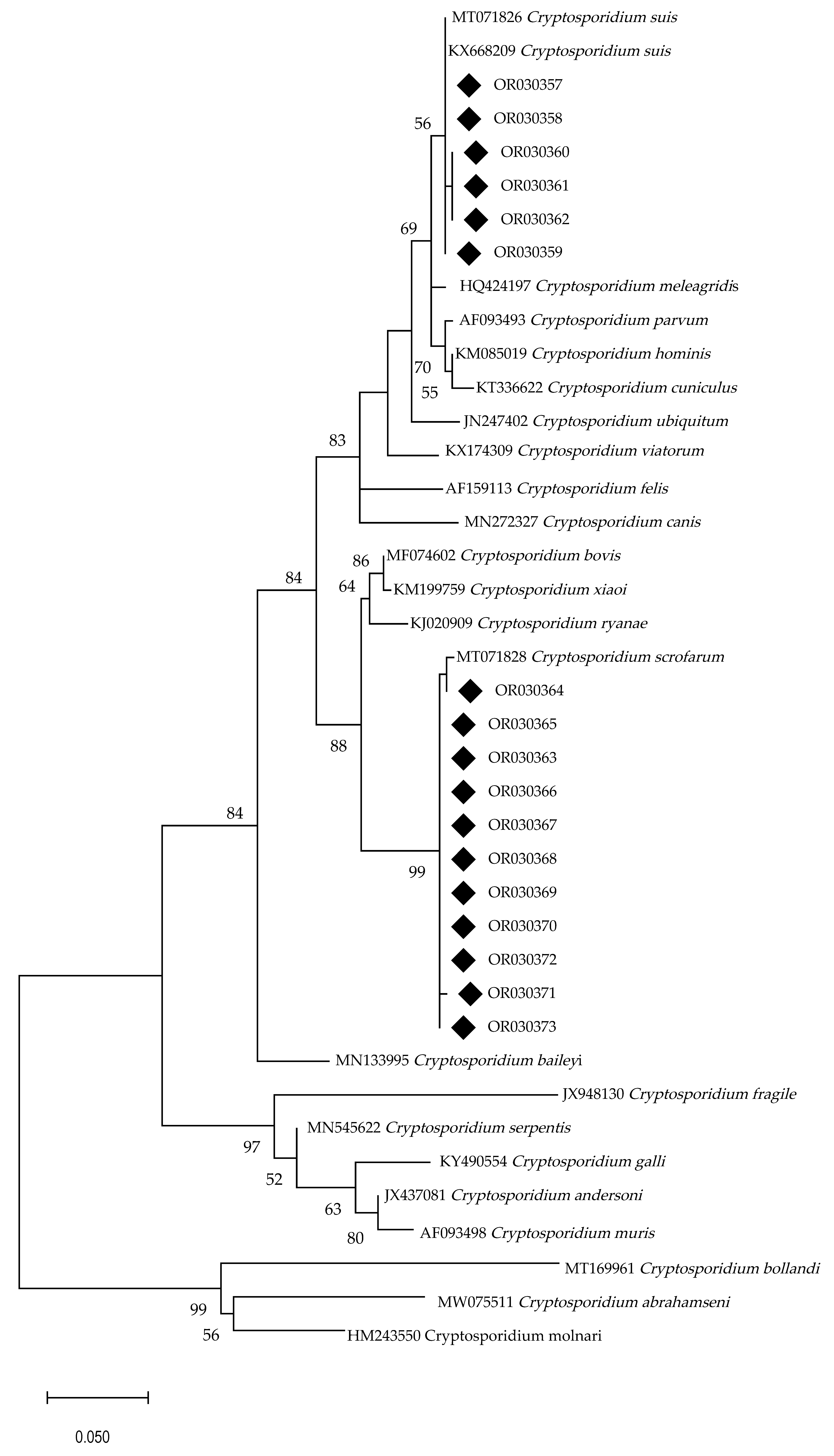
2.5. Sequencing and Phylogenetic Analysis
2.6. Data Analysis
2.6.1. Giardia duodenalis
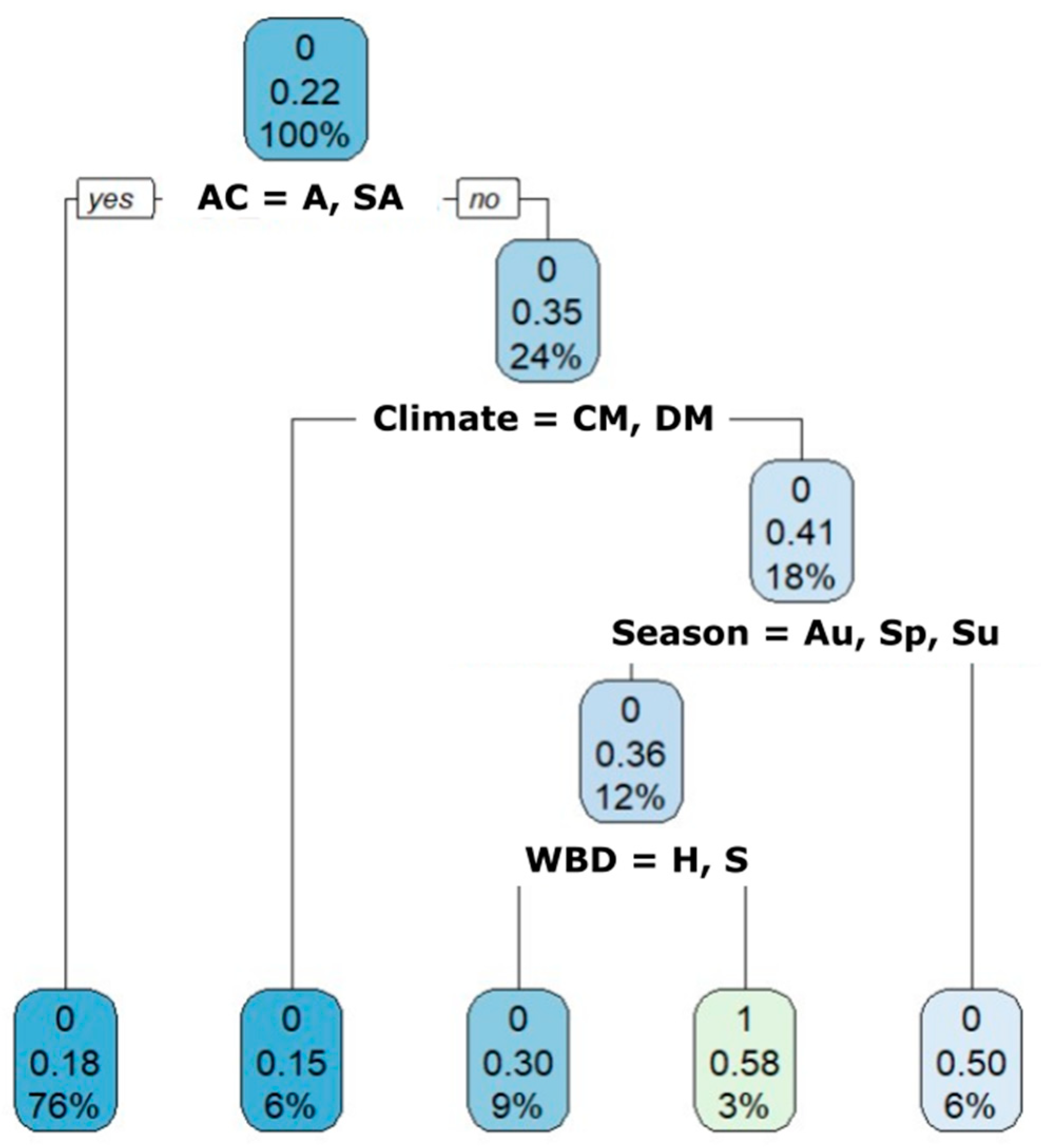
2.6.2. Cryptosporidium spp.
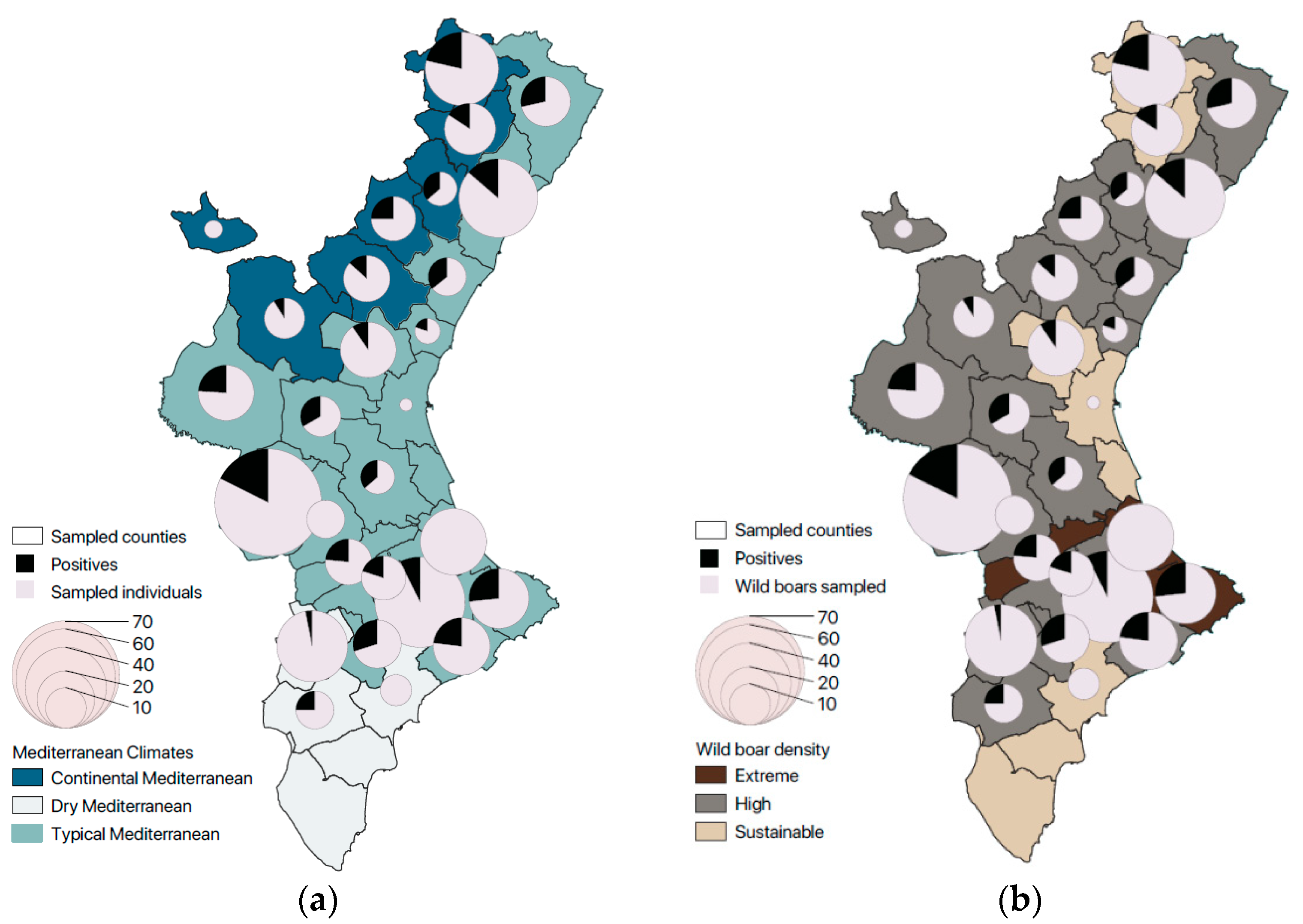
3. Results
3.1. Giardia duodenalis
3.2. Cryptosporidium spp.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild Boar Populations up, Numbers of Hunters down? A Review of Trends and Implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Gómez, C.; Nieto, M.d.P.; Burgui, J.M. Estadísticas Cinegéticas de La Comunidad Valenciana. Memoria 2021 (Temporada 2020/2021); Servicio de Caza y Pesca: Valencia, Spain, 2022. [Google Scholar]
- Hrazdilová, K.; Lesiczka, P.M.; Bardoň, J.; Vyroubalová, Š.; Šimek, B.; Zurek, L.; Modrý, D. Wild Boar as a Potential Reservoir of Zoonotic Tick-Borne Pathogens. Ticks Tick Borne Dis. 2021, 12, 101558. [Google Scholar] [CrossRef] [PubMed]
- Rhimi, W.; Sgroi, G.; Aneke, C.I.; Annoscia, G.; Latrofa, M.S.; Mosca, A.; Veneziano, V.; Otranto, D.; Alastruey-Izquierdo, A.; Cafarchia, C. Wild Boar (Sus scrofa) as Reservoir of Zoonotic Yeasts: Bioindicator of Environmental Quality. Mycopathologia 2022, 187, 235–248. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Marín, M.; López-Olvera, J.R.; Ayats, T.; Fernandez Aguilar, X.; Lavín, S.; Mentaberre, G.; Cerdà-Cuéllar, M. Zoonotic Campylobacter spp. and Salmonella spp. Carried by Wild Boars in a Metropolitan Area: Occurrence, Antimicrobial Susceptibility and Public Health Relevance. Sci. Total Environ. 2022, 822, 153444. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Dubey, J.P.; Lindsay, D.S. Zoonotic Protozoa: From Land to Sea. Trends Parasitol. 2004, 20, 531–536. [Google Scholar] [CrossRef]
- Thompson, R.C.A.; Monis, P.T. Variation in Giardia: Implications for Taxonomy and Epidemiology. Adv. Parasitol. 2004, 58, 69–137. [Google Scholar] [CrossRef]
- Appelbee, A.J.; Thompson, R.C.A.; Olson, M.E. Giardia and Cryptosporidium in Mammalian Wildlife—Current Status and Future Needs. Trends Parasitol. 2005, 21, 370–376. [Google Scholar] [CrossRef]
- Einarsson, E.; Ma’ayeh, S.; Svärd, S.G. An Up-Date on Giardia and Giardiasis. Curr. Opin. Microbiol. 2016, 34, 47–52. [Google Scholar] [CrossRef]
- Ryan, U.; Hijjawi, N.; Feng, Y.; Xiao, L. Giardia: An under-Reported Foodborne Parasite. Int. J. Parasitol. 2019, 49, 1–11. [Google Scholar] [CrossRef]
- Ignatius, R.; Gahutu, J.B.; Klotz, C.; Steininger, C.; Shyirambere, C.; Lyng, M.; Musemakweri, A.; Aebischer, T.; Martus, P.; Harms, G.; et al. High Prevalence of Giardia duodenalis Assemblage B Infection and Association with Underweight in Rwandan Children. PLoS Negl. Trop. Dis. 2012, 6, e1677. [Google Scholar] [CrossRef]
- Mohammed Mahdy, A.K.; Surin, J.; Wan, K.L.; Mohd-Adnan, A.; Al-Mekhlafi, M.S.H.; Lim, Y.A.L. Giardia intestinalis Genotypes: Risk Factors and Correlation with Clinical Symptoms. Acta Trop. 2009, 112, 67–70. [Google Scholar] [CrossRef]
- Horlock-Roberts, K.; Reaume, C.; Dayer, G.; Ouellet, C.; Cook, N.; Yee, J. Drug-Free Approach to Study the Unusual Cell Cycle of Giardia intestinalis. mSphere 2017, 2, e00384-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, M.E.; O’Handley, R.M.; Ralston, B.J.; McAllister, T.A.; Thompson, R.C.A. Update on Cryptosporidium and Giardia infections in Cattle. Trends Parasitol. 2004, 20, 185–191. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO (Food and Agriculture Organization of the United Nations and World Health Organization). Multicriteria-Based Ranking for Risk Management of Food-Borne Parasites. Available online: https://apps.who.int/iris/handle/10665/112672 (accessed on 3 January 2023).
- Chalmers, R.M.; Davies, A.P. Minireview: Clinical Cryptosporidiosis. Exp. Parasitol. 2010, 124, 138–146. [Google Scholar] [CrossRef]
- Chappell, C.L.; Okhuysen, P.C.; Langer-Curry, R.; Widmer, G.; Akiyoshi, D.E.; Tzipori, S. Cryptosporidium hominis: Experimental Challenge of Healthy Adults. Citeseer 2006, 75, 851–857. [Google Scholar] [CrossRef]
- Okhuysen, P.C.; Chappell, C.L.; Crabb, J.H.; Sterling, C.R.; DuPont, H.L. Virulence of Three Distinct Cryptosporidium parvum Isolates for Healthy Adults. J. Infect. Dis. 1999, 180, 1275–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teunis, P.F.M.; Chappell, C.L.; Okhuysen, P.C. Cryptosporidium dose response studies: Variation between isolates. Risk Anal. 2002, 22, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.; la Carbona, S.; Dumètre, A.; Robertson, L.J.; Gargala, G.; Escotte-Binet, S.; Favennec, L.; Villena, I.; Gérard, C.; Aubert, D. Assessing Viability and Infectivity of Foodborne and Waterborne Stages (Cysts/Oocysts) of Giardia duodenalis, Cryptosporidium spp., and Toxoplasma gondii: A Review of Methods. Parasite 2018, 25, 14. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.U.; Mi, R.; Zhou, S.; Gong, H.; Ullah, M.; Huang, Y.; Han, X.; Chen, Z. Advances in Therapeutic and Vaccine Targets for Cryptosporidium: Challenges and Possible Mitigation Strategies. Acta Trop. 2022, 226, 106273. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Zahedi, A.; Feng, Y.; Xiao, L. An Update on Zoonotic Cryptosporidium Species and Genotypes in Humans. Animals 2021, 11, 3307. [Google Scholar] [CrossRef] [PubMed]
- Diptyanusa, A.; Sari, I.P. Treatment of Human Intestinal Cryptosporidiosis: A Review of Published Clinical Trials. Int. J. Parasitol. Drugs Drug Resist. 2021, 17, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Sammarro Silva, K.J.; Sabogal-Paz, L.P. Cryptosporidium spp. and Giardia spp. (Oo)Cysts as Target-Organisms in Sanitation and Environmental Monitoring: A Review in Microscopy-Based Viability Assays. Water Res. 2021, 189, 116590. [Google Scholar] [CrossRef] [PubMed]
- Guadano Procesi, I.; Montalbano Di Filippo, M.; de Liberato, C.; Lombardo, A.; Brocherel, G.; Perrucci, S.; di Cave, D.; Berrilli, F. Giardia duodenalis in Wildlife: Exploring Genotype Diversity in Italy and across Europe. Pathogens 2022, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- Stojecki, K.; Sroka, J.; Cacció, S.M.; Cencek, T.; Dutkiewicz, J.; Kusyk, P. Prevalence and Molecular Typing of Giardia duodenalis in Wildlife from Eastern Poland. Folia Parasitol. 2015, 62, 42. [Google Scholar] [CrossRef] [Green Version]
- Kváč, M.; Kestřánová, M.; Pinková, M.; Květoňová, D.; Kalinová, J.; Wagnerová, P.; Kotková, M.; Vítovec, J.; Ditrich, O.; McEvoy, J.; et al. Cryptosporidium Scrofarum n. sp. (Apicomplexa: Cryptosporidiidae) in Domestic Pigs (Sus scrofa). Vet. Parasitol. 2013, 191, 218–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Němejc, K.; Sak, B.; Květoňová, D.; Hanzal, V.; Jeníková, M.; Kváč, M. The First Report on Cryptosporidium suis and Cryptosporidium Pig Genotype II in Eurasian Wild Boars (Sus scrofa) (Czech Republic). Vet. Parasitol. 2012, 184, 122–125. [Google Scholar] [CrossRef]
- Němejc, K.; Sak, B.; Květoňová, D.; Hanzal, V.; Janiszewski, P.; Forejtek, P.; Rajský, D.; Ravaszová, P.; McEvoy, J.; Kváč, M. Cryptosporidium suis and Cryptosporidium scrofarum in Eurasian Wild Boars (Sus scrofa) in Central Europe. Vet. Parasitol. 2013, 197, 504–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Presedo, I.; Pedraza-Díaz, S.; González-Warleta, M.; Mezo, M.; Gómez-Bautista, M.; Ortega-Mora, L.M.; Castro-Hermida, J.A. Presence of Cryptosporidium scrofarum, C. suis and C. parvum Subtypes IIaA16G2R1 and IIaA13G1R1 in Eurasian Wild Boars (Sus scrofa). Vet. Parasitol. 2013, 196, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidium Species in Humans and Animals: Current Understanding and Research Needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef] [Green Version]
- Bodager, J.R.; Parsons, M.B.; Wright, P.C.; Rasambainarivo, F.; Roellig, D.; Xiao, L.; Gillespie, T.R. Complex Epidemiology and Zoonotic Potential for Cryptosporidium suis in Rural Madagascar. Vet. Parasitol. 2015, 207, 140–143. [Google Scholar] [CrossRef]
- Kváč, M.; Květoňová, D.; Sak, B.; Ditrich, O. Cryptosporidium Pig Genotype II in immunocompetent man. Emerg. Infect. Dis. 2009, 15, 982–983. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Bern, C.; Arrowood, M.; Sulaiman, I.; Zhou, L.; Kawai, V.; Vivar, A.; Lal, A.A.; Gilman, R.H. Identification of the Cryptosporidium Pig Genotype in a Human Patient. J. Infect. Dis. 2002, 185, 1846–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, U.; Read, C.; Hawkins, P.; Warnecke, M.; Swanson, P.; Griffith, M.; Deere, D.; Cunningham, M.; Cox, P. Genotypes of Cryptosporidium from Sydney Water Catchment Areas. J. Appl. Microbiol. 2005, 98, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Colomé-Latorre, E.; González-López, E.; de Vicente y López, F.; Navarro-Baixauli, F. Plan de Acción Territorial Forestal de La Comunitat Valenciana—PATFOR; Documento Informativo o de Diagnosis; Elaborado Para: Dirección General Del Medio Natural; Conselleria de Infraestructuras, Territorio y Medio Ambiente, Generalitat Valenciana: Valencia, Spain, 2003. [Google Scholar]
- Castro-Hermida, J.A.; García-Presedo, I.; González-Warleta, M.; Mezo, M. Prevalence of Cryptosporidium and Giardia in Roe Deer (Capreolus capreolus) and Wild Boars (Sus scrofa) in Galicia (NW, Spain). Vet. Parasitol. 2011, 179, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Deng, L.; Wu, K.; Huang, X.; Song, Y.; Su, H.; Hu, Y.; Fu, H.; Zhong, Z.; Peng, G. Presence of Zoonotic Cryptosporidium scrofarum, Giardia duodenalis Assemblage A and Enterocytozoon bieneusi Genotypes in Captive Eurasian Wild Boars (Sus scrofa) in China: Potential for Zoonotic Transmission. Parasites Vectors 2017, 10, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero-Juarez, A.; Dashti, A.; López-López, P.; Muadica, A.S.; Risalde, M.D.L.A.; Risalde, M.D.L.A.; Köster, P.C.; Machuca, I.; Bailo, B.; De Mingo, M.H.; et al. Protist Enteroparasites in Wild Boar (Sus scrofa ferus) and Black Iberian Pig (Sus scrofa domesticus) in Southern Spain: A Protective Effect on Hepatitis E Acquisition? Parasites Vectors 2020, 13, 281. [Google Scholar] [CrossRef] [PubMed]
- De Blas, I.; Ortega, C.; Franjea, K.; Noordhuizen, J.; Trusfield, M. WinEpiscope 2.0 2011. Available online: http://www.winepi.net/sp/index.htm (accessed on 28 March 2019).
- Matschke, G.H. Ageing European Wild Hogs by Dentition. J. Wildl. Manag. 1967, 31, 109–113. [Google Scholar] [CrossRef]
- Saez-Royuela, C.; Gomariz, R.; Telleria, J.L. Age determination of European wild boar. Wildl. Soc. Bull. 1989, 17, 326–329. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- González Hidalgo, J.C.; de Luís, M.; Raventós, J.; Sánchez, J.R. Daily Rainfall Trend in the Valencia Region of Spain. Theor. Appl. Climatol. 2003, 75, 117–130. [Google Scholar] [CrossRef]
- De Luís, M.; Raventós, J.; González-Hidalgo, J.C.; Sánchez, J.R.; Cortina, J. Spatial Analysis of Rainfall Trends in the Region of Valencia (East Spain). Int. J. Climatol. 2000, 20, 1451–1469. [Google Scholar] [CrossRef]
- AVAMET-Associació Valenciana de Meteorologia. Precipitacions Mitjanes 2015–2018. Available online: https://www.avamet.org/mx-mapes.php?id=574&tipus=anuals&data=2018-01-01 (accessed on 5 January 2023).
- Kutta, E.; Hubbart, J.A. Reconsidering Meteorological Seasons in a Changing Climate. Clim. Chang. 2016, 137, 511–524. [Google Scholar] [CrossRef]
- Calvet Sanz, S.; Calafat, M.C.; Estellés, F.; Gallego, A.; Sanchis, E.; Moral, B. Diagnóstico de La Situación Ambiental de La Ganadería En La Comunitat Valenciana: Distribución de La Producción de Estiércoles y Generación de Emisiones Atmosféricas; Memoria Resultados Convenio Impacto Ambiental Granjas; UPV-GVA (Universitat Politècnica de València-Generalitat Valenciana): Valencia, Spain, 2021. [Google Scholar]
- Oroian, T.E.; Oroian, R.G.; Pasca, I.; Oroian, E.; Covrig, L. Methods of Age Estimation by Dentition in Sus scrofa ferus sp. Bull. Univ. Agric. Sci. Vet. Med. Cluj Napoca 2010, 67, 291–295. [Google Scholar]
- Rosell, C.; Fernández-Llario, P.; Herrero, J. El Jabalí (Sus scrofa LINNAEUS, 1758). Galemys 2001, 13, 1–25. [Google Scholar]
- European Union. Rural Areas and the Primary Sector in the EU. Available online: https://ec.europa.eu/agriculture/sites/agriculture/files/statistics/facts-figures/eu-rural-areas-primary-sector.pdf (accessed on 5 September 2019).
- Verweij, J.J.; Schinkel, J.; Laeijendecker, D.; van Rooyen, M.A.A.; van Lieshout, L.; Polderman, A.M. Real-Time PCR for the Detection of Giardia lamblia. Mol. Cell. Probes 2003, 17, 223–225. [Google Scholar] [CrossRef]
- Dacal, E.; Saugar, J.M.; De Lucio, A.; Hernández-De-Mingo, M.; Robinson, E.; Köster, P.C.; Aznar-Ruiz-De-Alegría, M.L.; Espasa, M.; Ninda, A.; Gandasegui, J.; et al. Prevalence and Molecular Characterization of Strongyloides stercoralis, Giardia duodenalis, Cryptosporidium spp., and Blastocystis spp. Isolates in School Children in Cubal, Western Angola. Parasites Vectors 2018, 11, 67. [Google Scholar] [CrossRef] [Green Version]
- Read, C.M.; Monis, P.T.; Thompson, R.C.A. Discrimination of All Genotypes of Giardia duodenalis at the Glutamate Dehydrogenase Locus Using PCR-RFLP. Infect. Genet. Evol. 2004, 4, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M.; Pozio, E.; Capelli, G.; Bruschi, F.; Crotti, D.; Cacciò, S.M. Genetic Heterogeneity at the Beta-Giardin Locus among Human and Animal Isolates of Giardia duodenalis and Identification of Potentially Zoonotic Subgenotypes. Int. J. Parasitol. 2005, 35, 207–213. [Google Scholar] [CrossRef]
- Ryan, U.; Xiao, L.; Read, C.; Zhou, L.; Lal, A.A.; Pavlasek, I. Identification of novel Cryptosporidium genotypes from the Czech Republic. Appl. Environ. Microbiol. 2003, 69, 4302–4307. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, version 4.1.0; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed; Springer: New York, NY, USA, 2022; ISBN 0-387-95457-0. [Google Scholar]
- Jackman, S. Pscl: Classes and Methods for R Developed in the Political Science Computational Laboratory; R Package Version 1.5.5; United States Studies Centre, University of Sydney: Sydney, NSW, Australia, 2020. [Google Scholar]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. Available online: https://www.jstatsoft.org/index.php/jss/article/view/v028i05 (accessed on 31 July 2023). [CrossRef] [Green Version]
- Williams, G. Data Mining with Rattle and R. The Art of Excavating Data for Knowledge Discovery; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Therneau, T.; Atkinson, B.; Ripley, B. “rpart”: Recursive Partitioning and Regression Trees. R Package Version 4.1-15. 2019. Available online: https://cran.r-project.org/src/contrib/Archive/rpart/ (accessed on 31 July 2023).
- Milborrow, S. rpart.plot: Plot ‘rpart’ Models: An Enhanced Version of ‘plot.rpart’. R Package Version 3.0.9. 2020. Available online: https://cran.r-project.org/src/contrib/Archive/rpart.plot/ (accessed on 31 July 2023).
- Feng, Y.; Xiao, L. Zoonotic Potential and Molecular Epidemiology of Giardia Species and Giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar] [CrossRef] [Green Version]
- Alum, A.; Absar, I.M.; Asaad, H.; Rubino, J.R.; Ijaz, M.K. Impact of Environmental Conditions on the Survival of Cryptosporidium and Giardia on Environmental Surfaces. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 210385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzan, A.; Parrington, L.; Coklin, T.; Cook, A.; Pintar, K.; Pollari, F.; Friendship, R.; Farber, J.; Dixon, B. Detection and Characterization of Giardia duodenalis and Cryptosporidium spp. on Swine Farms in Ontario, Canada. Foodborne Pathog. Dis. 2011, 8, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Adeyemo, F.E.; Singh, G.; Reddy, P.; Bux, F.; Stenström, T.A. Efficiency of Chlorine and UV in the Inactivation of Cryptosporidium and Giardia in Wastewater. PLoS ONE 2019, 14, e0216040. [Google Scholar] [CrossRef]
- Utaaker, K.S.; Skjerve, E.; Robertson, L.J. Keeping It Cool: Survival of Giardia Cysts and Cryptosporidium Oocysts on Lettuce Leaves. Int. J. Food Microbiol. 2017, 255, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, S.; Moraes, D.F.d.S.D.; López-López, P.; Palmeira, J.D.; Torres, R.T.; São José Nascimento, M.; Dashti, A.; Carmena, D.; Rivero-Juarez, A.; Mesquita, J.R. Survey of Zoonotic Diarrheagenic Protist and Hepatitis E Virus in Wild Boar (Sus scrofa) of Portugal. Animals 2023, 13, 256. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.; Köster, P.C.; Bailo, B.; de las Matas, A.S.; Habela, M.Á.; Rivero-Juarez, A.; Vicente, J.; Serrano, E.; Arnal, M.C.; de Luco, D.F.; et al. Occurrence and Limited Zoonotic Potential of Cryptosporidium spp., Giardia duodenalis, and Balantioides coli Infections in Free-Ranging and Farmed Wild Ungulates in Spain. Res. Vet. Sci. 2023, 159, 189–197. [Google Scholar] [CrossRef]
- Beck, R.; Sprong, H.; Lucinger, S.; Pozio, E.; Cacciò, S.M. A Large Survey of Croatian Wild Mammals for Giardia duodenalis Reveals a Low Prevalence and Limited Zoonotic Potential. Vector Borne Zoonotic Dis. 2011, 11, 1049–1055. [Google Scholar] [CrossRef]
- Spieler, N.; Schnyder, M. Lungworms (Metastrongylus spp.) and Intestinal Parasitic Stages of Two Separated Swiss Wild Boar Populations North and South of the Alps: Similar Parasite Spectrum with Regional Idiosyncrasies. Int. J. Parasitol. Parasites Wildl. 2021, 14, 202–210. [Google Scholar] [CrossRef]
- Erickson, M.C.; Ortega, Y.R. Inactivation of Protozoan Parasites in Food, Water, and Environmental Systems. J. Food Prot. 2006, 69, 2786–2808. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.; Rivero-Juárez, A.; Santín, M.; George, N.S.; Köster, P.C.; López-López, P.; Risalde, M.A.; García-Bocanegra, I.; Gómez-Villamandos, J.C.; Caballero-Gómez, J.; et al. Diarrhoea-Causing Enteric Protist Species in Intensively and Extensively Raised Pigs (Sus scrofa domesticus) in Southern Spain. Part I: Prevalence and Genetic Diversity. Transbound. Emerg. Dis. 2022, 69, e1051–e1064. [Google Scholar] [CrossRef] [PubMed]
- Vioque, F.; Dashti, A.; Santín, M.; Ruiz-Fons, F.; Köster, P.C.; Hernández-Castro, C.; García, J.T.; Bailo, B.; Ortega, S.; Olea, P.P.; et al. Wild Micromammal Host Spectrum of Zoonotic Eukaryotic Parasites in Spain. Occurrence and Genetic Characterisation. Transbound. Emerg. Dis. 2022, 69, e2926–e2942. [Google Scholar] [CrossRef] [PubMed]
- Mateo, M.; de Mingo, M.H.; de Lucio, A.; Morales, L.; Balseiro, A.; Espí, A.; Barral, M.; Lima Barbero, J.F.; Habela, M.Á.; Fernández-García, J.L.; et al. Occurrence and Molecular Genotyping of Giardia duodenalis and Cryptosporidium spp. in Wild Mesocarnivores in Spain. Vet. Parasitol. 2017, 235, 86–93. [Google Scholar] [CrossRef]
- Reboredo-Fernández, A.; Ares-Mazás, E.; Cacciò, S.M.; Gómez-Couso, H. Occurrence of Giardia and Cryptosporidium in Wild Birds in Galicia (Northwest Spain). Parasitology 2015, 142, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Hashemi-Hafshejani, S.; Meamar, A.R.; Moradi, M.; Hemmati, N.; Solaymani-Mohammadi, S.; Razmjou, E. Multilocus Sequence Typing of Giardia duodenalis Genotypes Circulating in Humans in a Major Metropolitan Area. Front. Med. 2022, 9, 976956. [Google Scholar] [CrossRef]
- Sprong, H.; Cacciò, S.M.; van der Giessen, J.W.B. Identification of Zoonotic Genotypes of Giardia duodenalis. PLoS Negl. Trop. Dis. 2009, 3, e558. [Google Scholar] [CrossRef] [Green Version]
- Foronda, P.; Bargues, M.D.; Abreu-Acosta, N.; Periago, M.V.; Valero, M.A.; Valladares, B.; Mas-Coma, S. Identification of Genotypes of Giardia intestinalis of Human Isolates in Egypt. Parasitol. Res. 2008, 103, 1177–1181. [Google Scholar] [CrossRef]
- Lizana, V.; Muniesa, A.; Cardells, J.; López-Ramon, J.; Aguiló-Gisbert, J.; Lomillos, J.M.; Gortázar, C. Safe Game: Hygienic Habits in Self-Consumption of Game Meat in Eastern Spain. Foods 2022, 11, 368. [Google Scholar] [CrossRef]
- Yui, T.; Nakajima, T.; Yamamoto, N.; Kon, M.; Abe, N.; Matsubayashi, M.; Shibahara, T. Age-Related Detection and Molecular Characterization of Cryptosporidium suis and Cryptosporidium scrofarum in Pre- and Post-Weaned Piglets and Adult Pigs in Japan. Parasitol. Res. 2014, 113, 359–365. [Google Scholar] [CrossRef]
- Naranjo, V.; Gortazar, C.; Vicente, J.; de la Fuente, J. Evidence of the Role of European Wild Boar as a Reservoir of Mycobacterium tuberculosis complex. Vet. Microbiol. 2008, 127, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kern, B.; Depner, K.R.; Letz, W.; Rott, M.; Thalheim, S.; Nitschke, B.; Plagemann, R.; Liess, B. Incidence of Classical Swine Fever (CSF) in Wild Boar in a Densely Populated Area Indicating CSF. Virus Persistence as a Mechanism for Virus Perpetuation. J. Vet. Med. Ser. B 1999, 46, 63–68. [Google Scholar] [CrossRef] [PubMed]
- King, B.J.; Monis, P.T. Critical Processes Affecting Cryptosporidium Oocyst Survival in the Environment. Parasitology 2007, 134, 309. [Google Scholar] [CrossRef] [PubMed]
- Rosell, C.; Navàs, F.; Romero, S. Reproduction of Wild Boar in a Cropland and Coastal Wetland Area: Implications for Management. Anim. Biodivers. Conserv. 2012, 35, 209–217. [Google Scholar] [CrossRef]
- Atwill, E.R.; Sweitzer, R.A.; Pereira, M.G.; Gardner, I.A.; Van Vuren, D.; Boyce, W.M. Prevalence of and Associated Risk Factors for Shedding Cryptosporidium parvum Oocysts and Giardia Cysts within Feral Pig Populations in California. Appl. Environ. Microbiol. 1997, 63, 3946–3949. [Google Scholar] [CrossRef] [PubMed]
- Slifko, T.R.; Smith, H.V.; Rose, J.B. Emerging Parasite Zoonoses Associated with Water and Food. Int. J. Parasitol. 2000, 30, 1379–1393. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R. Cryptosporidium: A Water-Borne Zoonotic Parasite. Vet. Parasitol. 2004, 126, 37–56. [Google Scholar] [CrossRef]
- Smith, H.V.; Cacciò, S.M.; Cook, N.; Nichols, R.A.B.; Tait, A. Cryptosporidium and Giardia as Foodborne Zoonoses. Vet. Parasitol. 2007, 149, 29–40. [Google Scholar] [CrossRef]
- Smith, H.V.; Nichols, R.A.B. Cryptosporidium: Detection in Water and Food. Exp. Parasitol. 2010, 124, 61–79. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County | Climate | Human Density | Land Use | Slurry | Hunting Area (ha) | Wild Boar Hunting Bag | Wild Boar Density | Minimum Sample Size | Achieved Sample Size |
|---|---|---|---|---|---|---|---|---|---|
| Alto Mijares | CM | R | F | 1 | 53,261.19 | 857 | H | 6.7 | 12 |
| BaixMaestrat | TM | R | F-I | 1 | 104,635.04 | 1480 | H | 11.6 | 15 |
| Els Ports | CM | R | F | 1 | 79,599.80 | 731 | S | 5.8 | 33 |
| L’Alcalatén | CM | R | F-I | 0 | 62,539.34 | 712 | H | 5.6 | 7 |
| Alt Maestrat | CM | R | F | 1 | 64,532.02 | 518 | S | 4.1 | 16 |
| Plana Alta | TM | I | F-I | 1 | 74,611.73 | 1283 | H | 10.1 | 38 |
| Plana Baixa | TM | I | I | 0 | 47,337.87 | 544 | H | 4.3 | 9 |
| Alto Palancia | CM | R | F-I | 0 | 87,353.56 | 1292 | H | 10.2 | 13 |
| TOTAL CASTELLÓN | 573,870.59 | 7416 | 58.4 | 143 | |||||
| Hoya de Buñol | TM | R | I | 0 | 74,865.52 | 913 | H | 7.2 | 10 |
| Ribera Alta | TM | I | I | 1 | 80,757.26 | 1021 | H | 8.0 | 7 |
| Camp de Morvedre | TM | I | I | 0 | 19,424.64 | 317 | H | 2.5 | 4 |
| Camp de Turia | TM | I | F-I | 1 | 64,412.67 | 518 | S | 4.1 | 19 |
| Rincón de Ademuz | CM | R | F | 0 | 36,663.14 | 490 | H | 3.9 | 2 |
| Valle de Cofrentes-Ayora | TM | R | F | 1 | 123,981.80 | 2765 | H | 21.8 | 70 |
| L’Horta | TM | U | I | 0 | 15,956.24 | 95 | S | 0.7 | 1 |
| Canal de Navarrés | TM | R | F | 0 | 55,325.44 | 577 | H | 4.5 | 9 |
| La Costera | TM | I | F-R | 0 | 45,618.45 | 1216 | E | 9.6 | 13 |
| La Plana de Utiel-Requena | TM | R | R | 1 | 162,835.77 | 2799 | H | 22 | 19 |
| Ribera Baixa | TM | I | I | - | 14,854.79 | 87 | S | 0.7 | - |
| La Safor | TM | I | I | 0 | 30,855.04 | 2399 | E | 11 | 27 |
| La Valld’Albaida | TM | I | F-R | 1 | 68,677.48 | 916 | H | 7.2 | 12 |
| Los Serranos | CM | R | F | 1 | 144,613.83 | 1393 | H | 11 | 10 |
| TOTAL VALENCIA | 938,842.14 | 14,505 | 114.1 | 203 | |||||
| Baix Segura | DM | I | R | - | 40,695.07 | 149 | S | 1.2 | - |
| BaixVinalopó | DM | U | R | - | 18,503.75 | 62 | S | 0.5 | - |
| El Comtat | TM | R | F | 0 | 35,521.49 | 878 | E | 6.9 | 50 |
| VinalopóMitjà | DM | I | R | 0 | 58,736.34 | 894 | H | 7 | 9 |
| L’Alacantí | DM | U | R | 0 | 39,683.17 | 349 | S | 2.7 | 6 |
| L’Alcoià | TM | I | F-R | 1 | 46,424.67 | 1099 | H | 8.6 | 14 |
| L’AltVinalopó | DM | R | R | 1 | 56,473.58 | 739 | H | 5.8 | 31 |
| Marina Alta | TM | I | F-I | 0 | 56,811.46 | 2034 | E | 16 | 22 |
| Marina Baixa | TM | I | F | 0 | 43,580.76 | 975 | H | 7.7 | 20 |
| TOTAL ALICANTE | 396,430.34 | 7179 | 56.5 | 152 | |||||
| TOTAL VC | 1,909,143.08 | 29,100 | 229 | 498 |
| Giardia duodenalis | Cryptosporidium suis | Cryptosporidium scrofarum | ||
|---|---|---|---|---|
| Climate | CM | 4.3% (4/93) | 1.1% (1/93) | 23.7% (22/93) |
| TM | 0.6% (2/359) | 3.3% (12/359) | 19.5% (70/359) | |
| DM | 0% (0/46) | 0% (0/46) | 8.7% (4/46) | |
| Season | Summer | 3.22% (1/31) | 0% (0/31) | 6.5% (2/31) |
| Autumn | 0.5% (1/209) | 1% (2/209) | 22.9% (48/209) | |
| Winter | 0.5% (1/195) | 3.6% (7/195) | 18.5% (36/195) | |
| Spring | 0% (0/63) | 4.8% (3/63) | 12.7% (8/63) | |
| Land use | Forested land | 1.3% (4/307) | 2.3% (7/307) | 20.1% (64/307) |
| Irrigated land | 0.9% (1/108) | 4.6% (5/108) | 18.5% (20/108) | |
| Rainfed land | 1.2% (1/83) | 1.2% (1/83) | 14.5% (12/83) | |
| Human density | Rural | 1.7% (5/297) | 8.4% (7/83) | 65.1% (54/83) |
| Intermediate | 0.5% (1/194) | 3.1% (6/194) | 21.6% (42/194) | |
| Urban | 0% (0/7) | 0% (0/7) | 0% (0/7) | |
| Wild boar density | Sustainable | 2.7% (2/75) | 0% (0/75) | 18.7% (14/75) |
| High | 0.9% (3/311) | 3.9% (12/311) | 21.5% (67/311) | |
| Extreme | 0.9% (1/112) | 0.9% (1/112) | 13.4% (15/112) | |
| Age group | Piglet | 0% (0/30) | 6.7% (2/30) | 30% (9/30) |
| Juvenile | 0% (0/83) | 2.4% (2/83) | 31.3% (26/83) | |
| Sub-adult | 2.7% (1/37) | 0% (0/37) | 13.5% (5/37) | |
| Adult | 1.2% (4/324) | 3.1% (10/324) | 15.1% (49/324) | |
| Unknown | 4.2% (1/24) | 0% (0/24) | 20.8% (5/24) | |
| Sex | Male | 1.3% (3/244) | 3.6% (8/244) | 19.6% (44/244) |
| Female | 0.8% (2/237) | 2.5% (6/237) | 19.8% (47/237) | |
| Unknown | 5.9% (1/17) | 0% (0/17) | 17.6% (3/17) |
| Factor | Estimate | S.E. | p-Value |
|---|---|---|---|
| Dry Mediterranean | −17.48 | 2672.95 | 0.99 |
| Typical Mediterranean | −2.74 | 1.12 | 0.015 |
| Species | N | Reference Sequence | % ID | SNV | Accession Number |
|---|---|---|---|---|---|
| C. suis | 6 | MT071826 | 100 | None | OR030357– OR030362 |
| 3 | MT071826 | 99.52–99.62 | InsT; T → A | ||
| 3 | KX668209 | 99.78–99.81 | DelA | ||
| 1 | MT561508 | 100 | None | ||
| C. scrofarum | 82 | MT071828 | 100 | None | OR030363– OR030373 |
| 5 | MT071828 | 99.78–99.8 | T → Y | ||
| 4 | MT071828 | 99.8–99.81 | T → C | ||
| 1 | MT071828 | 99.8 | A → R | ||
| 1 | MT071828 | 99.56 | C → T; T → C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martí-Marco, A.; Moratal, S.; Torres-Blas, I.; Cardells, J.; Lizana, V.; Dea-Ayuela, M.A. Molecular Detection and Epidemiology of Potentially Zoonotic Cryptosporidium spp. and Giardia duodenalis in Wild Boar (Sus scrofa) from Eastern Spain. Animals 2023, 13, 2501. https://doi.org/10.3390/ani13152501
Martí-Marco A, Moratal S, Torres-Blas I, Cardells J, Lizana V, Dea-Ayuela MA. Molecular Detection and Epidemiology of Potentially Zoonotic Cryptosporidium spp. and Giardia duodenalis in Wild Boar (Sus scrofa) from Eastern Spain. Animals. 2023; 13(15):2501. https://doi.org/10.3390/ani13152501
Chicago/Turabian StyleMartí-Marco, Alba, Samantha Moratal, Irene Torres-Blas, Jesús Cardells, Victor Lizana, and María Auxiliadora Dea-Ayuela. 2023. "Molecular Detection and Epidemiology of Potentially Zoonotic Cryptosporidium spp. and Giardia duodenalis in Wild Boar (Sus scrofa) from Eastern Spain" Animals 13, no. 15: 2501. https://doi.org/10.3390/ani13152501