1. Introduction
In recent years, direct-fed microbials (DFM) have been gaining more space into ruminant production settings, as these confer health benefits to the dairy animal [
1,
2,
3,
4], regardless of its age or category, while also improving nutrient utilization [
5,
6], overall performance of the herd [
7,
8], and the profitability of the dairy operations. Moreover, DFMs have been recognized as a potential alternative to antibiotic feeding in livestock production. Direct-fed microbials are often split into bacteria- or live yeast-based microorganisms and among the bacterial strains commonly used as DFMs for cattle, special consideration has been given to
Lactobacillus spp.,
Propionibacterium freudenreichii,
Enterococcus faecium,
Bifidobacterium spp., and
Bacillus spp. However, it is important to highlight that not all the aforementioned bacterial strains are able to survive and thrive in the same environments, which include different feed types, moisture, and high temperatures during feed preparation. Therefore, if the strains used in a commercial DFM are not able to survive the process of feed preparation, the health benefits expected may not be achieved and the success of the DFM might be questioned. Nonetheless, to date and to the best of our knowledge, no studies have evaluated the stability of lactic acid-producing bacteria, such as
E. faecium, when added to different feed matrices often included in dairy production systems (i.e., milk replacer and mineral–vitamin premix).
On the other hand,
Bacillus spp. are spore-forming bacteria that can survive the most challenging conditions, such as extreme temperatures, presence/absence of oxygen, wide pH ranges, the presence of radiation and toxic compounds, presence of other feed molecules and/or additives, as well as nutrient depletion in the environment [
9,
10,
11], as bacterial spores maintain their viability, but do not have metabolic activity. This last statement is important and key, as it shows that the
Bacillus spp. are not being lost during the feed manufacturing process [
10,
11], becoming active only when consumed by the animal (calf, developing heifer, or cow). However, limited information is available regarding the stability and recovery of
Bacillus spores following its mixture in different feed matrices often included in dairy production settings that employ different conditions during its preparation, such as temperature, salinity, and moisture.
Based on this rationale, we hypothesized that the recovery of E. faecium 669 and Bacillus spores following different feed preparations (pelletization, mineral–vitamin premix, and milk replacer) would be comparable and stable after inclusion in the different feed matrices. Hence, our objective was to evaluate the recovery of Bacillus spores following its mixture in pellets prepared under different temperatures (75 to 95 °C; Experiment 1), mineral–vitamin premix (Experiment 2), and milk replacer (Experiment 3), but also of E. faecium 669 following its inclusion into a mineral–vitamin premix (Experiment 4), as well as a milk replacer powder (Experiment 5) or milk replacer preparation (Experiment 6).
4. Discussion
Direct-fed microbials, also known as probiotics, are beneficial bacteria that support the health of the host, modulating the microbiome of the gastrointestinal tract (GIT), inhibiting the damaging effects of potentially harmful bacteria [
3,
4,
15,
16], and supporting the immune reactions of the host [
17]. However, in order to provide such benefits on health and ultimately on the performance of livestock animals, the bacterial strains used in commercially available DFM products must survive and drive the processes of feed preparation, which may include but are not limited to, low and high temperatures, presence of macro and trace minerals and high moisture, as well as long-term storage. Besides surviving the feed preparations, bacterial strains must also maintain their viability during the transit through the GIT of the ruminant animals, meaning that it will encounter changes in the type, quantity, and availability of substrates/nutrients, different pH values, competition from other bacteria (commensal or not) for nutrients and space, presence of bile salts, oxygen, and so on [
18]. It is also noteworthy that it was not the goal of the present experiment to compare the recovery of different strains when included in the same feed matrix, so isolated and single analyses were performed across the different Exp. conducted here.
Most of the bacterial strains, such as lactic acid-producing bacteria (LAPB), require an additional coating/protective process to maintain their viability before their introduction into different supplements [
19,
20,
21]. However, it is important to note that most of the studies performed with these bacteria have been performed under the conditions for human foods, which may not be representative of ruminant, and more specifically, dairy production settings. Hence, we evaluated the stability and recovery of
E. faecium (LACTIFERM
®), a well-known and heavily used bacterial strain for livestock [
22] when mixed using different feed matrices and different conditions. Based on the results reported herein,
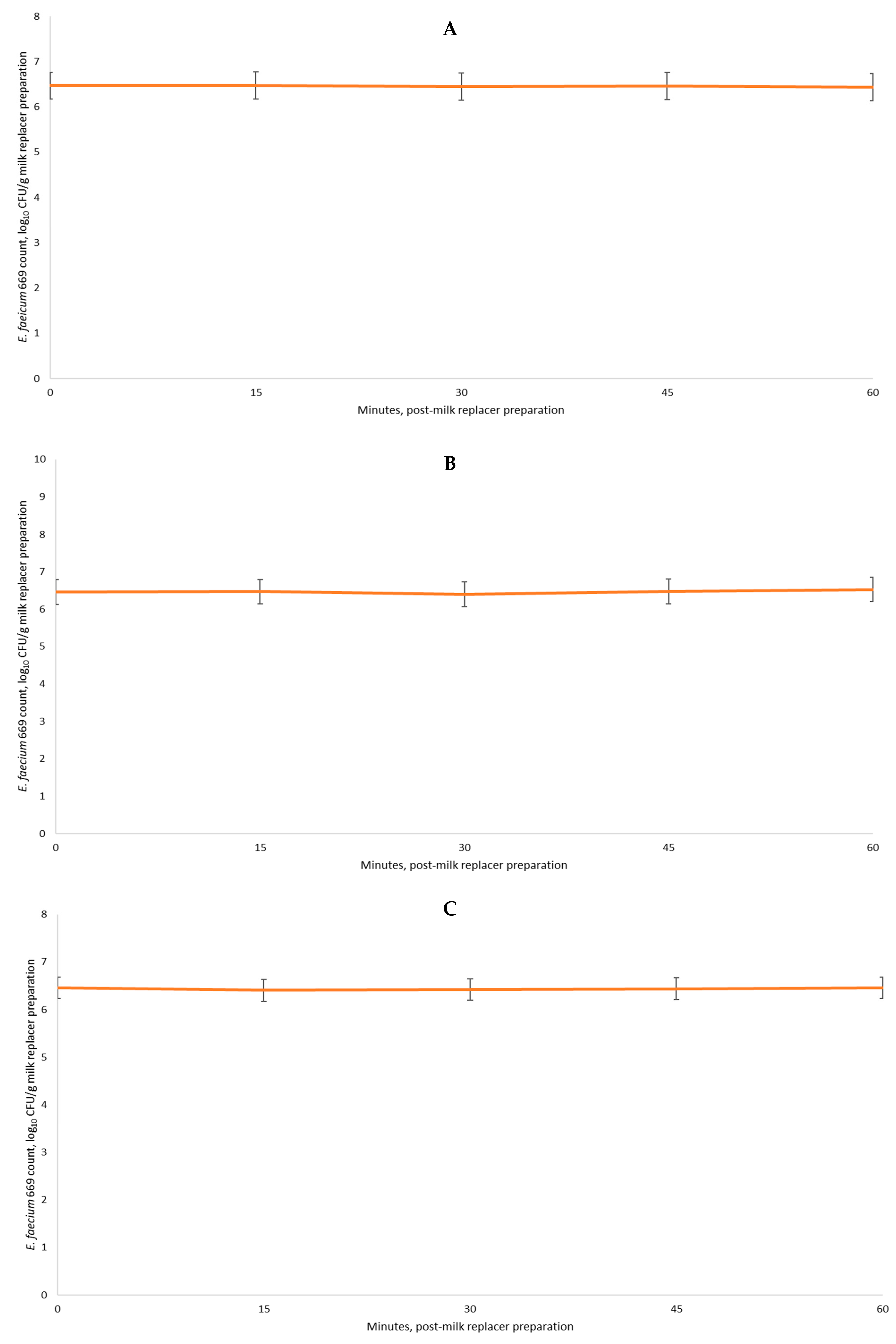
E. faecium 669 was stable over time (Exp. 4 and 6) even when exposed to different temperature conditions during milk replacer preparation and storage (Exp. 5), ensuring the benefits often observed in previous in vivo Exp. from our research group [
8,
23]. Experiments such as those reported here provide the foundation of understanding the features of different bacterial strains included in commercial products, demonstrating its stability and guidance of utilization in commercial settings. For example, the fact that
E. faecium 669 was stable over a long period of time following milk replacer preparation under different conditions is important and noteworthy, as it has been approved for use in pre-weaning and veal calves in the European Union [
13].
Conversely to LAPB, other bacterial strains, such as
Bacillus spp., present a natural resistance to several of the challenging factors, given its spore-forming feature [
10,
24]. More specifically, the spore-forming ability of
Bacillus spp. ensures its full viability following long-term storage [
24]. Moreover, bacilli spores can survive the challenging, low pH of the abomasum, reaching the small intestine at full viability to exert its beneficial effects on the host [
25]. Given all these features and its enzyme-producing ability [
26],
Bacillus spp. have been used as DFM for livestock species, supporting the health and improving nutrient utilization [
6,
27] and performance of ruminants, including beef animals [
7,
28], dairy calves [
29,
30,
31], and dairy cows [
32,
33,
34]. However, to the best of our knowledge, minimal data from previous research have been published evaluating the stability and recovery of
Bacillus spores in different feed matrices often used in dairy production systems, such as pelleted feed, mineral–vitamin premix, and a milk replacer. Therefore, these Exp. evaluated the recovery of spores included into a
Bacillus-based DFM (BOVACILLUS
TM) containing a mixture of
B. licheniformis and
B. subtilis when included in a pelleted feed (high-temperature; Exp. 1), commercial mineral–vitamin premix (presence of macro and trace minerals; Exp. 2), and a commercial milk replacer (high-temperature and moisture; Exp. 3).
Bacilli endospores are known to be highly resistant to physicochemical stress during feed production and storage, such as high pellet temperature, pressure, and shear forces [
24]. Previous research has demonstrated that
Bacillus spores are resistant to a pelleting temperature of up to 90 °C with over 90% of spores remaining viable in feed samples from poultry [
35]. In Exp. 1, using temperatures that ranged from 75 to 95 °C yielded the projected recovery of
Bacillus spores in pelleted feed for dairy cows. These data support the thermostability of
Bacillus spores, adding an important feature to these strains to be used as a reliable source of DFM for ruminants.
In Exp. 2., timepoints were taken over a 12-month period to assess the recovery of
Bacillus spores in a commercial mineral–vitamin premix when maintained in a room temperature environment (25 °C). Over time, no differences were observed among T1–T12 vs. T0, demonstrating the viability and resilience of this commercial DFM (BOVACILLUS
TM) when added to a commercial mineral–vitamin premix containing different macro and trace minerals. Lastly, in Exp. 3, the recovery of
Bacillus spores was assessed up to 60 min post-DFM (BOVACILLUS
TM) mixture and milk replacer preparation under two different temperatures (37 and 50 °C) often used in the industry and previously reported by others [
36]. The recovery of the
Bacillus spores was not impacted by the temperature of milk replacer preparation nor by the time points assessed herein, demonstrating, once again, its stability in an important feed matrix to the dairy industry.
Nonetheless and lastly, it is important for the personnel involved in the dairy production chain (industry, farmers, nutritionists, and veterinarians) to recognize the differences among the genera, species, and strains of bacteria being used in different research efforts, as differences in these features impact the observed results, or lack of such. For other feed additives, such as yeast, ionophores, and non-ionophores, the differences between strains have been demonstrated, impacting the overall performance of ruminants [
37,
38,
39].


 ,
, 

{kind=link}