
Effect of Atomized Black Maca (Lepidium meyenii) Supplementation in the Cryopreservation of Alpaca (Vicugna pacos) Epididymal Spermatozoa
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Obtaining the Epididymal Sperm
2.3. Sperm Cryopreservation and Maca Supplementation
2.4. Evaluation of Sperm Parameters
2.4.1. Sperm Motility
2.4.2. Sperm Viability
2.4.3. Plasma Membrane Integrity
2.4.4. DNA Fragmentation Analysis
2.4.5. Evaluation of ROS Production
2.4.6. Sperm Mitochondrial Activity
2.5. Statistical Analysis
3. Results
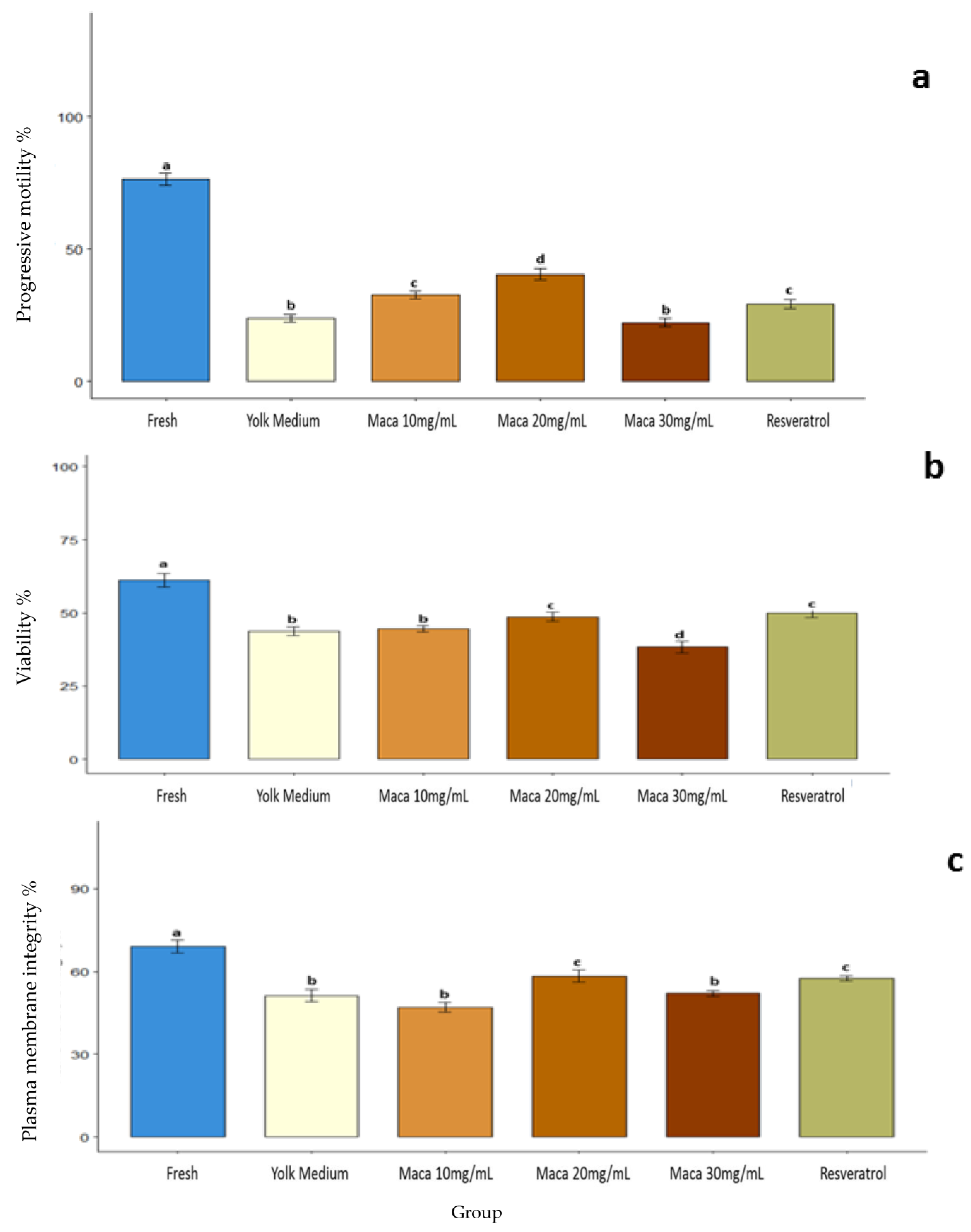
3.1. Effect of Maca (Lepidium meyenii) on Sperm Motility
3.2. Effect of Maca (Lepidium meyenii) on Sperm Viability
3.3. Effect of Maca (Lepidium meyenii) on the Plasma Membrane Integrity
3.4. Effect of Maca (Lepidium meyenii) on Reactive Oxygen Species (ROS)
3.5. Effect of Maca (Lepidium meyenii) on Sperm DNA Fragmentation
3.6. Effect of Maca (Lepidium meyenii) on Mitochondrial Activity Index
3.7. Effect of Maca (Lepidium meyenii) on Cytochemical Activity of Oxidoreductases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quina, E. Inseminación Artificial de Alpacas en un Contexto de Crianza Campesina; Descosur: Arequipa, Peru, 2017. [Google Scholar]
- Landeo, L.; Zuñiga, M.; Gastelu, T.; Artica, M.; Ruiz, J.; Silva, M.; Ratto, M.H. Oocyte Quality, In Vitro Fertilization and Embryo Development of Alpaca Oocytes Collected by Ultrasound-Guided Follicular Aspiration or from Slaughterhouse Ovaries. Animals 2022, 12, 1102. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C.; Beorlegui, N.; Beconi, M.T. Participation of superoxide anion in the capacitation of cryopreserved bovine sperm. Int. J. Androl. 2003, 26, 109–114. [Google Scholar] [CrossRef]
- Santiani, A.; Huanca, W.; Sapana, R.; Huanca, T.; Sepulveda, N.; Sanchez, R. Effects on the quality of frozen-thawed alpaca (Lama pacos) semen using two different cryoprotectants and extenders. Asian J. Androl. 2005, 7, 303–309. [Google Scholar] [CrossRef]
- Casaretto, C.; Sarrasague, M.M.; Giuliano, S.; de Celis, E.R.; Gambarotta, M.; Carretero, I.; Miragaya, M. Evaluation of Lama glama semen viscosity with a cone-plate rotational viscometer. Andrologia 2011, 44, 335–341. [Google Scholar] [CrossRef]
- O’neill, H.C.; Nikoloska, M.; Ho, H.; Doshi, A.; Maalouf, W. Improved cryopreservation of spermatozoa using vitrification: Comparison of cryoprotectants and a novel device for long-term storage. J. Assist. Reprod. Genet. 2019, 36, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Rosato, M.P.; Iaffaldano, N. Cryopreservation of rabbit semen: Comparing the effects of different cryoprotectants, cryoprotectant-free vitrification, and the use of albumin plus osmoprotectants on sperm survival and fertility after standard vapor freezing and vitrification. Theriogenology 2013, 79, 508–516. [Google Scholar] [CrossRef]
- Agarwal, A.; Makker, K.; Sharma, R. REVIEW ARTICLE: Clinical Relevance of Oxidative Stress in Male Factor Infertility: An Update. Am. J. Reprod. Immunol. 2007, 59, 2–11. [Google Scholar] [CrossRef]
- Du Plessis, S.; Makker, K.; Desai, N.; Agarwal, A. The impact of oxidative stress on in vitro fertilization. Expert Rev. Obstet. Gynecol. 2008, 3, 539–554. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treulen, F.; Arias, M.E.; Aguila, L.; Uribe, P.; Felmer, R. Cryopreservation induces mitochondrial permeability transition in a bovine sperm model. Cryobiology 2018, 83, 65–74. [Google Scholar] [CrossRef]
- Treulen, F.; Aguila, L.; Arias, M.E.; Jofré, I.; Felmer, R. Impact of post-thaw supplementation of semen extender with antioxidants on the quality and function variables of stallion spermatozoa. Anim. Reprod. Sci. 2018, 201, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Shojaeian, K.; Nouri, H.; Kohram, H. Does MnTBAP ameliorate DNA fragmentation and in vivo fertility of frozen-thawed Arabian stallion sperm? Theriogenology 2018, 108, 16–21. [Google Scholar] [CrossRef]
- Amidi, F.; Pazhohan, A.; Shabani Nashtaei, M.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Ros-Santaella, J.L.; Pintus, E. Plant Extracts as Alternative Additives for Sperm Preservation. Antioxidants 2021, 10, 772. [Google Scholar] [CrossRef]
- Pasquariello, R.; Verdile, N.; Brevini, T.A.L.; Gandolfi, F.; Boiti, C.; Zerani, M.; Maranesi, M. The Role of Resveratrol in Mammalian Reproduction. Molecules 2020, 25, 4554. [Google Scholar] [CrossRef]
- Piacente, S.; Carbone, V.; Plaza, A.; Zampelli, A.; Pizza, C. Investigation of the Tuber Constituents of Maca (Lepidium meyeniiWalp.). J. Agric. Food Chem. 2002, 50, 5621–5625. [Google Scholar] [CrossRef]
- Gonzales, G.F.; Córdova, A.; Vega, K.; Chung, A.; Villena, A.; Góñez, C. Effect of Lepidium meyenii (Maca), a root with aphrodisiac and fertility-enhancing properties, on serum reproductive hormone levels in adult healthy men. J. Endocrinol. 2003, 176, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.L.; He, K.; Kim, C.H.; Rogers, L.; Shao, Y.; Huang, Z.Y.; Lu, Y.; Yan, S.J.; Qien, L.C.; Zheng, Q.Y. Effect of a lipidic extract from Lepidium meyenii on sexual behavior in mice and rats. Urology 2000, 55, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Zenico, T.; Cicero, A.F.G.; Valmorri, L.; Mercuriali, M.; Bercovich, E. Subjective effects of Lepidium meyenii (Maca) extract on well-being and sexual performances in patients with mild erectile dysfunction: A randomised, double-blind clinical trial. Andrologia 2009, 41, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, C.; Rubio, J.; Gasco, M.; Nieto, J.; Yucra, S.; Gonzales, G.F. Effect of short-term and long-term treatments with three ecotypes of Lepidium meyenii (MACA) on spermatogenesis in rats. J. Ethnopharmacol. 2006, 103, 448–454. [Google Scholar] [CrossRef]
- Sandoval, M.; Okuhama, N.N.; Angeles, F.M.; Melchor, V.V.; Condezo, L.A.; Lao, J.; Miller, M.J. Antioxidant activity of the cruciferous vegetable Maca (Lepidium meyenii). Food Chem. 2002, 79, 207–213. [Google Scholar] [CrossRef]
- Lee, K.-J.; Dabrowski, K.; Sandoval, M.; Miller, M.J. Activity-guided fractionation of phytochemicals of maca meal, their antioxidant activities and effects on growth, feed utilization, and survival in rainbow trout (Oncorhynchus mykiss) juveniles. Aquaculture 2005, 244, 293–301. [Google Scholar] [CrossRef]
- Zha, S.; Zhao, Q.; Chen, J.; Wang, L.; Zhang, G.; Zhang, H.; Zhao, B. Extraction, purification and antioxidant activities of the polysaccharides from maca (Lepidium meyenii). Carbohydr. Polym. 2014, 111, 584–587. [Google Scholar] [CrossRef]
- Valdivia, M..; Yarasca, K.; Levano, G.; Vasquez, J.; Temoche, H.; Torres, L.; Cruz, V. Effect of Lepidium meyenii(maca) on testicular function of mice with chemically and physically induced subfertility. Andrologia 2016, 48, 927–934. [Google Scholar] [CrossRef]
- Rubio, J.; Riqueros, M.I.; Gasco, M.; Yucra, S.; Miranda, S.; Gonzales, G.F. Lepidium meyenii (Maca) reversed the lead acetate induced-damage on reproductive function in male rats. Food Chem. Toxicol. 2006, 44, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Castañeda, C.; Rivera, V.; Chirinos, A.L.; Evelson, P.; Gonzales, G.F. Photoprotection against the UVB-induced oxidative stress and epidermal damage in mice using leaves of three different varieties of Lepidium meyenii (maca). Int. J. Dermatol. 2011, 50, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Dzięcioł, M.; Wróblewska, A.; Janda-Milczarek, K. Comparative Studies of DPPH Radical Scavenging Activity and Content of Bioactive Compounds in Maca (Lepidium meyenii) Root Extracts Obtained by Various Techniques. Appl. Sci. 2023, 13, 4827. [Google Scholar] [CrossRef]
- Del Prete, C.; Tafuri, S.; Ciani, F.; Pasolini, M.P.; Ciotola, F.; Albarella, S.; Carotenuto, D.; Peretti, V.; Cocchia, N. Influences of dietary supplementation with Lepidium meyenii (Maca) on stallion sperm production and on preservation of sperm quality during storage at 5 °C. Andrology 2018, 6, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Leiva-Revilla, J.; Rolón, M.; Siyadatpanah, A.; Pereira, M.D.L.; Nissapatorn, V. First study of in vitro protective effect of Lepidium meyenii (Maca) on frozen–thawed bovine spermatozoa. Vet. World 2022. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Tsujimura, A.; Nagashima, Y.; Hiramatsu, I.; Uesaka, Y.; Nozaki, T.; Ogishima, T.; Shirai, M.; Shoyama, Y.; Tanaka, H.; et al. Effect of Lepidium meyenii on in vitro fertilization via improvement in acrosome reaction and motility of mouse and human sperm. Reprod. Med. Biol. 2018, 18, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales-Arimborgo, C.; Yupanqui, I.; Montero, E.; Alarcón-Yaquetto, D.E.; Zevallos-Concha, A.; Caballero, L.; Gasco, M.; Zhao, J.; Khan, I.A.; Gonzales, G.F. Acceptability, Safety, and Efficacy of Oral Administration of Extracts of Black or Red Maca (Lepidium meyenii) in Adult Human Subjects: A Randomized, Double-Blind, Placebo-Controlled Study. Pharmaceuticals 2016, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Bravo, Z.; Valdivia, M. Follicular fluid stimulates capacitation and acrosome reaction in alpaca sperm (Vicugna pacos). Reprod. Domest. Anim. 2018, 53, 629–635. [Google Scholar] [CrossRef]
- Canorio, N.; Paredes, F.; Valdivia, M. Agentes crioprotectores alternativos para el congelamiento lento de espermatozoides epididimarios de alpaca (Vicugna pacos). Rev. De Investig. Vet. Del Perú 2015, 26, 434–443. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021.
- Jeyendran, R.S.; Van Der Ven, H.H.; Perez-Pelaez, M.; Crabo, B.G.; Zaneveld, L.J.D. Development of an assay to assess the functional integrity of the human sperm membrane and its relationship to other semen characteristics. Reproduction 1984, 70, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Santiani, A.; Evangelista, S.; Cheuquemán, C.; von Baer, A.; Risopatrón, J.; Sánchez, R. Evaluación de la integridad de ADN mediante citometría de flujo en espermatozoides de alpaca criopreservados con análogos de superóxido dismutasa. Rev. De Investig. Vet. Del Perú 2012, 23, 182–191. [Google Scholar]
- Nova, Z.; Skovierova, H.; Strnadel, J.; Halasova, E.; Calkovska, A. Short-Term versus Long-Term Culture of A549 Cells for Evaluating the Effects of Lipopolysaccharide on Oxidative Stress, Surfactant Proteins and Cathelicidin LL-37. Int. J. Mol. Sci. 2020, 21, 1148. [Google Scholar] [CrossRef] [Green Version]
- Hrudka, F. Cytochemistry of Oxidoreductases in Spermatozoa: The Technique Revisited. Andrologia 2009, 11, 337–353. [Google Scholar] [CrossRef]
- Torres, L.; Guevara, B.; Cruz, V.; Valdivia, M. Myrciaria dubia “camu camu” flour as a magnetoprotector in male mouse infertility. Bioelectromagnetics 2019, 40, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Morton, K.M.; Bathgate, R.; Evans, G.; Maxwell, W.M.C. Cryopreservation of epididymal alpaca (Vicugna pacos) sperm: A comparison of citrate-, Tris- and lactose-based diluents and pellets and straws. Reprod. Fertil. Dev. 2007, 19, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.C.; de Verdier, K.; Båge, R.; Morrell, J.M. Semen collection methods in alpacas. Vet. Rec. 2017, 180, 613–614. [Google Scholar] [CrossRef]
- Tamburrino, L.; Traini, G.; Marcellini, A.; Vignozzi, L.; Baldi, E.; Marchiani, S. Cryopreservation of Human Spermatozoa: Functional, Molecular and Clinical Aspects. Int. J. Mol. Sci. 2023, 24, 4656. [Google Scholar] [CrossRef]
- Pabón, D.; Meseguer, M.; Sevillano, G.; Cobo, A.; Romero, J.L.; Remohí, J.; Santos, M.J.D.L. A new system of sperm cryopreservation: Evaluation of survival, motility, DNA oxidation, and mitochondrial activity. Andrology 2019, 7, 293–301. [Google Scholar] [CrossRef]
- Mamani-Mango, G.; Gonzales, M.M.; Hidalgo, M.R.; Mallma, J.M.; Béjar, J.R.; Palma, V.R.; Salas, E.M. Effect of Extender and Freezing Rate on Quality Parameters and In Vitro Fertilization Capacity of Alpaca Spermatozoa Recovered from Cauda Epididymis. Biopreservation Biobanking 2019, 17, 39–45. [Google Scholar] [CrossRef]
- Juárez, J.; Santiani, A. Determinación del porcentaje de viabilidad espermática mediante citometría de flujo durante el proceso de criopreservación en espermatozoides obtenidos de epidídimo de alpaca. Rev. De Investig. Vet. Del Perú 2019, 30, 1175–1183. [Google Scholar] [CrossRef]
- Valdivia, M.; Bravo, Z.; Reyes, J.; Gonzales, G.F. Rescue and Conservation of Male Adult Alpacas (Vicugna pacos) Based on Spermatogonial Stem Cell Biotechnology Using Atomized Black Maca as a Supplement of Cryopreservation Medium. Front. Vet. Sci. 2021, 8, 597964. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, G.; Varela, E.; Duque, J.E.; Gómez, J.E.; Rojas, M. Freezing, Vitrification, and Freeze-Drying of Equine Spermatozoa: Impact on Mitochondrial Membrane Potential, Lipid Peroxidation, and DNA Integrity. J. Equine Vet. Sci. 2019, 72, 8–15. [Google Scholar] [CrossRef]
- Aurich, C.; Ferrusola, C.O.; Vega, F.J.P.; Schrammel, N.; Morcuende, D.; Aurich, J. Seasonal changes in the sperm fatty acid composition of Shetland pony stallions. Theriogenology 2018, 107, 149–153. [Google Scholar] [CrossRef]
- Tafuri, S.; Cocchia, N.; Carotenuto, D.; Vassetti, A.; Staropoli, A.; Mastellone, V.; Peretti, V.; Ciotola, F.; Albarella, S.; Del Prete, C.; et al. Chemical Analysis of Lepidium meyenii (Maca) and Its Effects on Redox Status and on Reproductive Biology in Stallions. Molecules 2019, 24, 1981. [Google Scholar] [CrossRef] [Green Version]
- Inoue, N.; Farfan, C.; Gonzales, G.F. Effect of butanolic fraction of yellow and black maca (Lepidium meyenii) on the sperm count of adult mice. Andrologia 2016, 48, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Večeřa, R.; Orolin, J.; Škottová, N.; Kazdová, L.; Oliyarnik, O.; Ulrichová, J.; Šimánek, V. The Influence of Maca (Lepidium meyenii) on Antioxidant Status, Lipid and Glucose Metabolism in Rat. Plant Foods Hum. Nutr. 2007, 62, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, F. Chemical composition and health effects of maca (Lepidium meyenii). Food Chem. 2019, 288, 422–443. [Google Scholar] [CrossRef]
- Gonzales, G.; Cordova, A.; Gonzales, C.; Chung, A.; Vega, K.; Villena, A. Lepidium meyenii (Maca) improved semen parameters in adult men. Asian J. Androl. 2001, 3, 301–303. [Google Scholar]
- Gonzales, G.; Ruiz, A.; Gonzales, C.; Villegas, L.; Cordova, A. Effect of Lepidium meyenii (maca) roots on spermatogenesis of male rats. Asian J. Androl. 2001, 3, 231–233. [Google Scholar] [PubMed]
- Onaolapo, A.Y.; Oladipo, B.P.; Onaolapo, O.J. Cyclophosphamide-induced male subfertility in mice: An assessment of the potential benefits of Maca supplement. Andrologia 2018, 50, e12911. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Mattson, M.P.; Calabrese, V. Resveratrol commonly displays hormesis: Occurrence and biomedical significance. Hum. Exp. Toxicol. 2010, 29, 980–1015. [Google Scholar] [CrossRef] [PubMed]
- Ghaleno, L.R.; Alizadeh, A.; Drevet, J.R.; Shahverdi, A.; Valojerdi, M.R. Oxidation of Sperm DNA and Male Infertility. Antioxidants 2021, 10, 97. [Google Scholar] [CrossRef]
- Carracedo, S.; Briand-Amirat, L.; Dordas-Perpinyà, M.; Escuredo, Y.R.; Delcombel, R.; Sergeant, N.; Delehedde, M. ProAKAP4 protein marker: Towards a functional approach to male fertility. Anim. Reprod. Sci. 2022, 247, 107074. [Google Scholar] [CrossRef]
- Malo, C.; Carracedo, S.; Delehedde, M.; Sergeant, N.; Skidmore, J.A. Identification of proAKAP4 concentration variations in dromedary sperm and their correlation with monthly semen parameters. Reprod. Fertil. 2021, 2, 268–279. [Google Scholar] [CrossRef]
- Blommaert, D.; Sergeant, N.; Delehedde, M.; Donnay, I.; Lejeune, J.P.; Franck, T.; Serteyn, D. First results about ProAKAP4 concentration in stallion semen after cryopreservation in two different freezing media. Cryobiology 2021, 102, 133–135. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Gałęska, E.; Bubel, A. The Concentration of ProAKAP4 and Other Indicators of Cryopotential of Spermatozoa Cryopreserved in Extender with Holothuroidea Extract Addition. Animals 2022, 12, 521. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rate Activity (%) | ||||||
|---|---|---|---|---|---|---|
| Compact (C) | Sub-Compact (Sub-c) | Focal (F) | Residual (Res) | Intensive Reaction (C+Sub-c) (%) | Reduced Reaction (F+Res) (%) | |
| Fresh | 38 ± 19.4 | 13.5 ± 9.42 | 0.90 ±0.65 | 0 | 51.5 a | 0.9 a |
| * Yolk Medium | 44.2 ± 32.8 | 26.0 ± 19.2 | 0.50 ±0.79 | 0.15 ± 0.28 | 70.2 b | 0.65 b |
| * Maca 10 mg/mL | 47.9 ± 32.1 | 13.7 ± 9.50 | 0.55 ± 1.50 | 0.01 ± 0.03 | 61.6 b | 0.55 c |
| * Maca 20 mg/mL | 21.3 ± 5.61 | 16.9 ± 22.4 | 0.38 ± 0.45 | 0.03 ± 0.12 | 38.2 c | 0.41 c |
| * Maca 30 mg/mL | 40.3 ± 21.2 | 11.0 ± 8.44 | 0.53 ± 0.73 | 0.08 ± 0.15 | 51.3 a | 0.61 b |
| * Resveratrol | 23.5 ± 12.4 | 11.5 ± 9.66 | 2.22 ± 0.46 | 0.02 ± 0.06 | 35 c | 2.22 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levano, G.; Quispe, J.; Vargas, D.; García, M.; López, A.; Aguila, L.; Valdivia, M. Effect of Atomized Black Maca (Lepidium meyenii) Supplementation in the Cryopreservation of Alpaca (Vicugna pacos) Epididymal Spermatozoa. Animals 2023, 13, 2054. https://doi.org/10.3390/ani13132054
Levano G, Quispe J, Vargas D, García M, López A, Aguila L, Valdivia M. Effect of Atomized Black Maca (Lepidium meyenii) Supplementation in the Cryopreservation of Alpaca (Vicugna pacos) Epididymal Spermatozoa. Animals. 2023; 13(13):2054. https://doi.org/10.3390/ani13132054
Chicago/Turabian StyleLevano, Gloria, Juana Quispe, Diego Vargas, Marlon García, Alberto López, Luis Aguila, and Martha Valdivia. 2023. "Effect of Atomized Black Maca (Lepidium meyenii) Supplementation in the Cryopreservation of Alpaca (Vicugna pacos) Epididymal Spermatozoa" Animals 13, no. 13: 2054. https://doi.org/10.3390/ani13132054