

Transcriptomic Response of the Liver Tissue in Trachinotus ovatus to Acute Heat Stress
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Heat Stress Experiment and Sample Collection
2.2. RNA Extraction, Library Construction, and Sequencing
2.3. Sequencing Quality Detection and Comparison
2.4. Differential Gene Expression Analysis
2.5. STEM Analysis
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Validation of mRNA Expression Patterns
3. Results
3.1. Transcriptome Sequencing
3.2. Differential Gene Expression Analysis
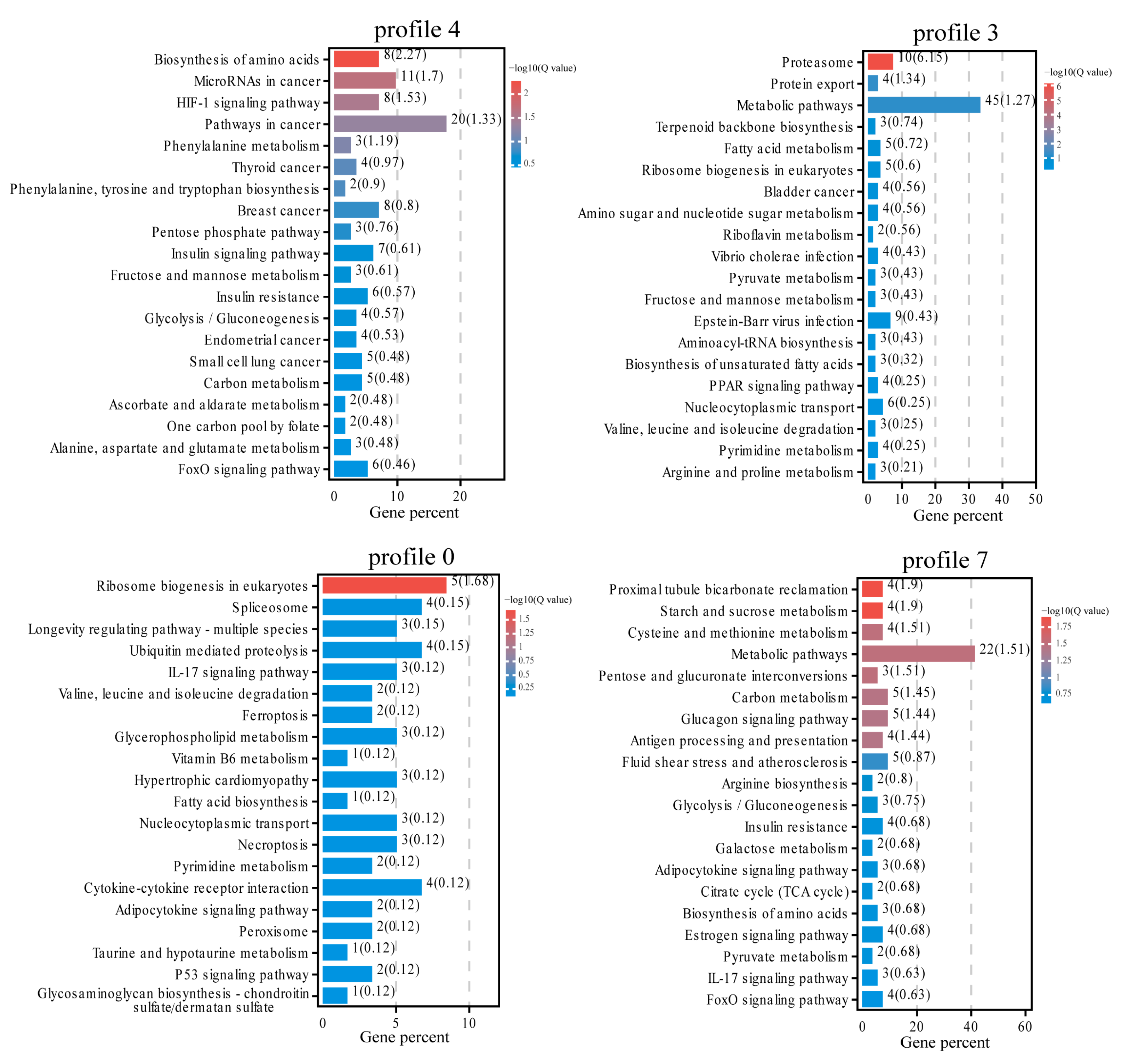
3.3. Expression Trend and Enrichment Analysis of DEGs
3.4. Validation of Gene Expression Patterns by qRT-PCR
4. Discussion
4.1. Protein Homeostasis
4.1.1. HSPs and Correct Folding of Proteins
4.1.2. Ubiquitin-Proteasome System (UPS) and Protein Degradation
4.2. Adaptation to Hypoxia and Transition of Energy Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; Food and Agriculture Organization of the United Nation: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- McKenzie, D.J.; Geffroy, B.; Farrell, A.P. Effects of global warming on fishes and fisheries. J. Fish Biol. 2021, 98, 1489–1492. [Google Scholar] [CrossRef]
- Chen, Q.Z.; Zeng, J.N.; Gao, A.G.; Liao, Y.B.; Yang, G.M. Advances in study of temperature of thermal tolerance of fishes. J. Fish. China 2004, 5, 120–123. [Google Scholar]
- Whitehead, A.; Galvez, F.; Zhang, S.; Williams, L.M.; Oleksiak, M.F. Functional genomics of physiological plasticity and local adaptation in killifish. J. Hered. 2011, 102, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity: Biodiversity and climate change. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhuang, P.; Yan, A.; Zhang, L. Optimum temperatures for growth and feeding of juvenile amur sturgeon Acipenser schrenckii. J. Fish. Sci. China 2005, 3, 294–299. [Google Scholar]
- Handeland, S.O.; Imsland, A.K.; Stefansson, S.O. The effect of temperature and fish size on growth, feed intake, food conversion efficiency and stomach evacuation rate of atlantic salmon post-smolts. Aquaculture 2008, 283, 36–42. [Google Scholar] [CrossRef]
- Ahmad, S.M.; Shah, F.A.; Bhat, F.A.; Bhat, J.I.A.; Balkhi, M.H. Thermal adaptability and disease association in common carp (Cyprinus carpio communis) acclimated to different (four) temperatures. J. Therm. Biol. 2011, 36, 492–497. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, Z.; Song, Z.; Xiao, P.; Liu, Y.; Zhang, P.; You, F. Insight into the heat resistance of fish via blood: Effects of heat stress on metabolism, oxidative stress and antioxidant response of olive flounder Paralichthys olivaceus and turbot Scophthalmus maximus. Fish Shellfish Immun. 2016, 58, 125–135. [Google Scholar] [CrossRef]
- Guillen, A.C.; Borges, M.E.; Herrerias, T.; Kandalski, P.K.; de Arruda Marins, E.; Viana, D.; de Souza, M.R.D.P.; Oliveira do Carmo Daloski, L.; Donatti, L. Effect of gradual temperature increase on the carbohydrate energy metabolism responses of the antarctic fish Notothenia rossii. Mar. Environ. Res. 2019, 150, 104779. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ke, H.; Zhang, L.; Zhao, Z.; Lai, J.; Zhou, J.; Huang, Z.; Li, H.; Du, J.; Li, Q. Integrated analysis about the effects of heat stress on physiological responses and energy metabolism in Gymnocypris chilianensis. Sci. Total Environ. 2022, 806, 151252. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Wu, X.Y.; Lai, J.S.; Liu, Y.; Song, M.J.; Li, F.Y.; Gong, Q. Integrated biochemical, transcriptomic and metabolomic analyses provide insight into heat stress response in Yangtze sturgeon (Acipenser dabryanus). Ecotoxicol. Environ. Saf. 2023, 249, 114366. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Sabry, A.; Abdelaziz, A.; Shukry, M. Deleterious impacts of heat stress on steroidogenesis markers, immunity status and ovarian tissue of Nile tilapia (Oreochromis niloticus). J. Therm. Biol. 2020, 91, 102578. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, Y.; Li, B.; Ding, L.; Wei, X.; Wang, P.; Chen, Z.; Han, S.; Huang, T.; Wang, B.; et al. Physiological responses to heat stress in the liver of rainbow trout (Oncorhynchus mykiss) revealed by UPLC-QTOF-MS metabolomics and biochemical assays. Ecotoxicol. Environ. Saf. 2022, 242, 113949. [Google Scholar] [CrossRef]
- Pankhurst, N.W.; King, H.R. Temperature and salmonid reproduction: Implications for aquaculture. J. Fish Biol. 2010, 76, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Ji, K.; Choi, K. Effects of water temperature on perchlorate toxicity to the thyroid and reproductive system of Oryzias latipes. Ecotoxicol. Environ. Saf. 2014, 108, 311–317. [Google Scholar] [CrossRef]
- Singh, S.P.; Ahmad, T.; Sharma, J.; Chakrabarti, R. Effect of temperature on food consumption, immune system, antioxidant enzymes, and heat shock protein 70 of Channa punctata (bloch, 1793). Fish Physiol. Biochem. 2021, 47, 79–91. [Google Scholar] [CrossRef]
- Chadwick, J.G.; McCormick, S.D. Upper thermal limits of growth in brook trout and their relationship to stress physiology. J. Exp. Biol. 2017, 220, 3976–3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomalty, K.M.H.; Meek, M.H.; Stephens, M.R.; Rincón, G.; Fangue, N.A.; May, B.P.; Baerwald, M.R. Transcriptional response to acute thermal exposure in juvenile chinook salmon determined by RNAseq. G3 Genes Genom. Genet. 2015, 5, 1335–1349. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Xu, X.; Li, J.; Shen, Y. Transcriptomic analysis of the liver and brain in grass carp (Ctenopharyngodon idella) under heat stress. Mar. Biotechnol. 2022, 24, 856–870. [Google Scholar] [CrossRef]
- Klumb, R.A.; Rudstam, L.G.; Mills, E.L. Comparison of alewife young-of-the-year and adult respiration and swimming speed bioenergetics model parameters: Implications of extrapolation. Trans. Am. Fish. Soc. 2003, 132, 1089–1103. [Google Scholar] [CrossRef]
- Li, Y.S.; Bai, S.L.; Pan, L.Z.; Guan, W.J.; Jiao, J.P. An individual model based simulation study on impact of water temperature fluctuation on recruitment of chub mackerel (Scomber japonicus) in the East China Sea. Trans. Oceanol. Limnol. 2018, 6, 118–124. [Google Scholar] [CrossRef]
- Wang, G.Q.; Xia, J.G. Effects of constant and diel-fluctuating temperature on thermal tolerance of zebrafish at different life-history stages. Chin. J. Ecol. 2019, 38, 2133–2137. [Google Scholar] [CrossRef]
- Madeira, D.; Madeira, C.; Costa, P.M.; Vinagre, C.; Pörtner, H.O.; Diniz, M.S. Different sensitivity to heatwaves across the life cycle of fish reflects phenotypic adaptation to environmental niche. Mar. Environ. Res. 2020, 162, 105192. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Xiong, T.; Wang, Y.Q.; Hao, S.X.; Huang, H. Effects of heat treatment temperature on quality and physicochemical properties of muscle protein in Trachinotus ovatus. J. Guangdong Ocean Univ. 2022, 42, 1–10. [Google Scholar]
- Xun, P.; Lin, H.; Wang, R.; Huang, Z.; Zhou, C.; Yu, W.; Huang, Q.; Tan, L.; Wang, Y.; Wang, J. Effects of dietary vitamin b1 on growth performance, intestinal digestion and absorption, intestinal microflora and immune response of juvenile golden pompano (Trachinotus ovatus). Aquaculture 2019, 506, 75–83. [Google Scholar] [CrossRef]
- Liu, Q. Effect of Flow Velocity on Swimming Behavior of Trachinotus ovatus. Master’s Thesis, Guangdong Ocean University, Zhanjiang, China, 2022. [Google Scholar] [CrossRef]
- You, C.; Chen, B.; Wang, M.; Wang, S.; Zhang, M.; Sun, Z.; Juventus, A.J.; Ma, H.; Li, Y. Effects of dietary lipid sources on the intestinal microbiome and health of golden pompano (Trachinotus ovatus). Fish Shellfish Immun. 2019, 89, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhou, C.; Lin, H.; Wang, J.; Wang, Y.; Yu, W. Effect of dietary carbohydrate on glycometabolism of juvenile golden pompano (Trachinotus ovatus) related to growth performance, hepatic histopathology, transcriptome profiles and identification of differentially expressed genes. Aquac. Rep. 2022, 27, 101362. [Google Scholar] [CrossRef]
- Reid, G.K.; Gurney-Smith, H.J.; Marcogliese, D.J.; Knowler, D.; Benfey, T.; Garber, A.F.; Forster, I.; Chopin, T.; Brewer-Dalton, K.; Moccia, R.D.; et al. Climate change and aquaculture: Considering biological response and resources. Aquac. Environ. Interact. 2019, 11, 569–602. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Lin, H.; Huang, Z.; Wang, J.; Wang, Y.; Yu, W. Cloning and expression analysis of c-type lysozyme gene in golden pompano, Trachinotus ovatus. Fish Shellfish Immun. 2016, 54, 580–585. [Google Scholar] [CrossRef]
- Li, G.; Zhao, D.; Huang, L.; Sun, J.; Gao, D.; Wang, H.; Tan, Y.; Liang, L. Identification and phylogenetic analysis of vibrio vulnificus isolated from diseased Trachinotus ovatus in cage mariculture. Aquaculture 2006, 261, 17–25. [Google Scholar] [CrossRef]
- Liu, B.; Guo, H.Y.; Zhu, K.C.; Guo, L.; Liu, B.S.; Zhang, N.; Yang, J.W.; Jiang, S.G.; Zhang, D.C. Growth, physiological, and molecular responses of golden pompano Trachinotus ovatus (linnaeus, 1758) reared at different salinities. Fish Physiol. Biochem. 2019, 45, 1879–1893. [Google Scholar] [CrossRef]
- Zhao, W.; Fang, H.H.; Gao, B.Y.; Dai, C.M.; Liu, Z.Z.; Zhang, C.W.; Niu, J. Dietary Tribonema Sp. Supplementation increased growth performance, antioxidant capacity, immunity and improved hepatic health in golden pompano (Trachinotus ovatus). Aquaculture 2020, 529, 735667. [Google Scholar] [CrossRef]
- Xie, D.; Wang, M.; Wang, S.; You, C.; Li, M.; Ma, Y.; Sun, L.; Wang, Y.; Li, Y. Fat powder can be a feasible lipid source in aquafeed for the carnivorous marine teleost golden pompano, Trachinotus ovatus. Aquac. Int. 2020, 28, 1153–1168. [Google Scholar] [CrossRef]
- Ma, Z.; Guo, H.; Zheng, P.; Wang, L.; Jiang, S.; Zhang, D.; Qin, J.G. Effect of salinity on the rearing performance of juvenile golden pompano Trachinotus ovatus (linnaeus 1758). Aquac. Res. 2016, 47, 1761–1769. [Google Scholar] [CrossRef]
- San, L.; Liu, B.; Liu, B.; Guo, H.; Guo, L.; Zhang, N.; Zhu, K.; Jiang, S.; Zhang, D. Transcriptome analysis of gills provides insights into translation changes under hypoxic stress and reoxygenation in golden pompano, Trachinotus ovatus (linnaeus 1758). Front. Mar. Sci. 2021, 8, 763622. [Google Scholar] [CrossRef]
- Li, J.L.; Chen, G.; Zhang, J.D.; Tang, B.G.; Zhou, H.; Wang, Z.L.; Huang, J.S.; Shi, G.; Chu, Q.Z.; Li, X.M. Effects of temperature and salinity on the respiratory metabolism of derbio (Trachinotus ovatus L.). J. Guangdong Ocean Univ. 2014, 34, 30–36. [Google Scholar]
- Ma, Z.; Zhang, N.; Qin, J.G.; Fu, M.; Jiang, S. Water temperature induces jaw deformity and bone morphogenetic proteins (BMPs) gene expression in golden pompano Trachinotus ovatus larvae. SpringerPlus 2016, 5, 1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q. Effect of temperature on growth, survival and occurrence of skeletal deformity in the golden pompano Trachinotus ovatus larvae. Indian J. Fish. 2016, 63, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Yang, R.; Chen, X.; Fu, Z.; Ma, Z.; Yu, G. Transcriptional response of golden pompano Trachinotus ovatus larvae to cold and heat stress. Aquac. Rep. 2021, 20, 100755. [Google Scholar] [CrossRef]
- Han, M.Y.; Zhou, S.J.; Yang, R.; Hu, J.; Ma, Z.H. Histopathology and molecular characterization of the skeletal tissues of golden pomopano (Trachinotus ovatus) larvae under temperature stress. J. South. Agr. 2021, 52, 3147–3156. [Google Scholar]
- Dettleff, P.; Zuloaga, R.; Fuentes, M.; Gonzalez, P.; Aedo, J.; Estrada, J.M.; Molina, A.; Valdés, J.A. High-temperature stress effect on the red cusk-eel (Geypterus chilensis) liver: Transcriptional modulation and oxidative stress damage. Biology 2022, 11, 990. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.C. Whole Genome Sequencing of Female Pompano (Trachinotus ovatus). 2019. Available online: https://figshare.com/articles/dataset/Whole_genome_sequencing_of_female_pompano_Trachinotus_ovatus_/7570727/3 (accessed on 17 May 2023).
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinkawa, T.; Taoka, M.; Yamauchi, Y.; Ichimura, T.; Kaji, H.; Takahashi, N.; Isobe, T. STEM: A software tool for large-scale proteomic data analyses. J. Proteome Res. 2005, 4, 1826–1831. [Google Scholar] [CrossRef]
- Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From fastq data to high-confidence variant calls: The genome analysis toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 1110. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Sung, Y.Y.; MacRae, T.H. Heat shock proteins and disease control in aquatic organisms. J. Aquac. Res. Dev. 2013, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.X.; Ma, G.S. Biological function of heat shock proteins. Foreign Med. Mol. Biol. Vol. 1993, 15, 275–278. [Google Scholar]
- Ryckaert, J.; Pasmans, F.; Tobback, E.; Duchateau, L.; Decostere, A.; Haesebrouck, F.; Sorgeloos, P.; Bossier, P. Heat shock proteins protect platyfish (Xiphophorus maculatus) from Yersinia ruckeri induced mortality. Fish Shellfish Immun. 2010, 28, 228–231. [Google Scholar] [CrossRef]
- Liu, T.; Daniels, C.K.; Cao, S. Comprehensive review on the HSC70 functions, interactions with related molecules and involvement in clinical diseases and therapeutic potential. Pharmacol. Ther. 2012, 136, 354–374. [Google Scholar] [CrossRef]
- Tsan, M.F.; Gao, B. Cytokine function of heat shock proteins. Am. J. Physiol. Cell Physiol. 2004, 286, C739–C744. [Google Scholar] [CrossRef] [Green Version]
- Subbarao Sreedhar, A.; Kalmár, É.; Csermely, P.; Shen, Y.-F. HSP90 isoforms: Functions, expression and clinical importance. FEBS Lett. 2004, 562, 11–15. [Google Scholar] [CrossRef]
- Burkhardt-Holm, P.; Schmidt, H.; Meier, W. Heat shock protein (HSP70) in brown trout epidermis after sudden temperature rise. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 120, 35–41. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, B.; Xie, J.; Xu, P.; Tsion, H.M.H.; Zhang, Y. The effect of hyperthermia on cell viability, oxidative damage, and heat shock protein expression in hepatic cells of grass carp (Ctenopharyngodon idellus). J. Therm. Biol. 2013, 38, 355–361. [Google Scholar] [CrossRef]
- Shin, M.K.; Park, H.R.; Yeo, W.J.; Han, K.N. Effects of thermal stress on the mRNA expression of SOD, HSP90, and HSP70 in the spotted sea bass (Lateolabrax maculatus). Ocean Sci. J. 2018, 53, 43–52. [Google Scholar] [CrossRef]
- Huang, J.; Li, Y.; Liu, Z.; Kang, Y.; Wang, J. Transcriptomic responses to heat stress in rainbow trout Oncorhynchus mykiss head kidney. Fish Shellfish Immun. 2018, 82, 32–40. [Google Scholar] [CrossRef]
- Topal, A.; Özdemir, S.; Arslan, H.; Çomaklı, S. How does elevated water temperature affect fish brain? (a neurophysiological and experimental study: Assessment of brain derived neurotrophic factor, cFOS, apoptotic genes, heat shock genes, ER-stress genes and oxidative stress genes). Fish Shellfish Immun. 2021, 115, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.B.; Yoon, J.H.; Park, J.Y.; Lee, I.Y.; Lim, H.K. A comparison of the physiological responses to heat stress of juvenile and adult starry flounder (Platichthys stellatus). Isr. J. Aquac. Bamidgeh. 2021, 73, 1–15. [Google Scholar] [CrossRef]
- Manzon, L.A.; Zak, M.A.; Agee, M.; Boreham, D.R.; Wilson, J.Y.; Somers, C.M.; Manzon, R.G. Thermal acclimation alters both basal heat shock protein gene expression and the heat shock response in juvenile lake whitefish (Coregonus clupeaformis). J. Therm. Biol. 2022, 104, 103185. [Google Scholar] [CrossRef] [PubMed]
- Amm, I.; Sommer, T.; Wolf, D.H. Protein quality control and elimination of protein waste: The role of the ubiquitin–proteasome system. Biochim. Biophys. Acta BBA Mol. Cell Res. 2014, 1843, 182–196. [Google Scholar] [CrossRef] [Green Version]
- Pickart, C.M.; Eddins, M.J. Ubiquitin: Structures, functions, mechanisms. Biochim. Biophys. Acta BBA Mol. Cell Res. 2004, 1695, 55–72. [Google Scholar] [CrossRef] [Green Version]
- Tokumoto, M.; Lee, J.Y.; Fujiwara, Y.; Uchiyama, M.; Satoh, M. Inorganic arsenic induces apoptosis through downregulation of UBE2D genes and p53 accumulation in rat proximal tubular cells. J. Toxicol. Sci. 2013, 38, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Fairall, L.; Goult, B.T.; Calkin, A.C.; Hong, C.; Millard, C.J.; Tontonoz, P.; Schwabe, J.W.R. The IDOL–UBE2D complex mediates sterol-dependent degradation of the LDL receptor. Genes Dev. 2011, 25, 1262–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berndsen, C.E.; Wolberger, C. New insights into ubiquitin E3 ligase mechanism. Nat. Struct. Mol. Biol. 2014, 21, 301–307. [Google Scholar] [CrossRef]
- Choi, M.; Choi, Y.M.; An, I.-S.; Bae, S.; Jung, J.H.; An, S. E3 ligase RCHY1 negatively regulates HDAC2. Biochem. Biophys. Res. Commun. 2020, 521, 37–41. [Google Scholar] [CrossRef]
- Windisch, H.S.; Frickenhaus, S.; John, U.; Knust, R.; Pörtner, H.-O.; Lucassen, M. Stress response or beneficial temperature acclimation: Transcriptomic signatures in antarctic fish (Pachycara brachycephalum). Mol. Ecol. 2014, 23, 3469–3482. [Google Scholar] [CrossRef]
- Tang, C.H.; Leu, M.Y.; Shao, K.; Hwang, L.Y.; Chang, W.B. Short-term effects of thermal stress on the responses of branchial protein quality control and osmoregulation in a reef-associated fish, Chromis viridis. Zool. Stud. 2014, 53, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Bai, Y.; Hu, X.; Yang, X.; Xu, S.; Cheng, B. Stress response of rainbow trout fry after exposure to an elevated concentration of carbon dioxide based on transcriptome analysis. Ichthyol. Res. 2022, 70, 215–224. [Google Scholar] [CrossRef]
- Lee, J.Y.; Tokumoto, M.; Hattori, Y.; Fujiwara, Y.; Shimada, A.; Satoh, M. Different regulation of p53 expression by cadmium exposure in kidney, liver, intestine, vasculature, and brain astrocytes. Toxicol. Res. 2016, 32, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Yu, H.; Mim, C.; Matouschek, A. Regulated protein turnover: Snapshots of the proteasome in action. Nat. Rev. Mol. Cell Biol. 2014, 15, 122–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glickman, M.H.; Rubin, D.M.; Fu, H.; Larsen, C.N.; Coux, O.; Wefes, I.; Pfeifer, G.; Cjeka, Z.; Vierstra, R.; Baumeister, W. Functional analysis of the proteasome regulatory particle. Mol. Biol. Rep. 1999, 26, 21–28. [Google Scholar] [CrossRef]
- Ferrell, K.; Wilkinson, C.R.M.; Dubiel, W.; Gordon, C. Regulatory subunit interactions of the 26s proteasome, a complex problem. Trends Biochem. Sci. 2000, 25, 83–88. [Google Scholar] [CrossRef]
- Lee, C.K.; Klopp, R.G.; Weindruch, R.; Prolla, T.A. Gene expression profile of aging and its retardation by caloric restriction. Sci. New Ser. 1999, 285, 1390–1393. [Google Scholar] [CrossRef] [Green Version]
- Finley, D. Recognition and processing of ubiquitin-protein conjugates by the proteasome. Annu. Rev. Biochem. 2009, 78, 477–513. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.S.; Cai, C.F.; Xu, F.; Lin, X.X.; Huang, Y.W.; Dong, J.J.; Zhang, B.T.; Xiao, P.Z. Effects of oxidized fish oil on expression levels of antioxidant stress pathway genes in intestinal mucosa of Grass carp. Acta Hydrobiol. Sin. 2016, 40, 758–766. [Google Scholar] [CrossRef]
- Zhu, R. The Mechanism of Changes on Activity of Ubiquitin Proteasome Pathway after Exercise in Rat Skeletal Muscle. Doctoral Thesis, Beijing Sport University, Beijing, China, 2009. [Google Scholar]
- Xu, N.; Liu, X.L.; Liu, L.; Tang, R.; Li, D.P. Effects of saprolegniasis on blood biochemical parameters and muscular ubiquitin-proteasome system in grass carp (Ctenopharyngodon idellus). Chin. Fish. Qual. Stand. 2021, 11, 44–50. [Google Scholar] [CrossRef]
- Hu, G.; Shu, Y.; Luan, P.; Zhang, T.; Chen, F.; Zheng, X. Genomic analysis of the proteasome subunit gene family and their response to high density and saline-alkali stresses in grass carp. Fishes 2022, 7, 350. [Google Scholar] [CrossRef]
- Todgham, A.E.; Crombie, T.A.; Hofmann, G.E. The effect of temperature adaptation on the ubiquitin-proteasome pathway in notothenioid fishes. J. Exp. Biol. 2016, 220, 145946. [Google Scholar] [CrossRef] [Green Version]
- Olsvik, P.A.; Vikeså, V.; Lie, K.K.; Hevrøy, E.M. Transcriptional responses to temperature and low oxygen stress in atlantic salmon studied with next-generation sequencing technology. BMC Genom. 2013, 14, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Maldonado, S.; Camacho-Cervantes, M. Effect of a temperature gradient on the behaviour of an endangered Mexican topminnow and an invasive freshwater fish. Sci. Rep. 2022, 12, 20584. [Google Scholar] [CrossRef]
- Krzywinska, E.; Stockmann, C. Hypoxia, metabolism and immune cell function. Biomedicines 2018, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Eckle, T.; Köhler, D.; Lehmann, R.; El Kasmi, K.C.; Eltzschig, H.K. Hypoxia-inducible factor-1 is central to cardioprotection: A new paradigm for ischemic preconditioning. Circulation 2008, 118, 166–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivan, M.; Kondo, K.; Yang, H.; Kim, W.; Valiando, J.; Ohh, M.; Salic, A.; Asara, J.M.; Lane, W.S.; Kaelin, W.G., Jr. HIFα targeted for VHL-mediated destruction by proline hydroxylation: Implications for O2 sensing. Sci. New Ser. 2001, 292, 464–468. [Google Scholar] [CrossRef]
- Geng, X.; Feng, J.; Liu, S.; Wang, Y.; Arias, C.; Liu, Z. Transcriptional regulation of hypoxia inducible factors alpha (HIF-α) and their inhibiting factor (FIH-1) of channel catfish (Ictalurus punctatus) under hypoxia. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2014, 169, 38–50. [Google Scholar] [CrossRef]
- Rytkönen, K.T.; Akbarzadeh, A.; Miandare, H.K.; Kamei, H.; Duan, C.; Leder, E.H.; Williams, T.A.; Nikinmaa, M. Subfunctionalization of cyprinid hypoxia-inducible factors for roles in development and oxygen sensing. Evolution 2013, 67, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Ton, C.; Stamatiou, D.; Liew, C.C. Gene expression profile of zebrafish exposed to hypoxia during development. Physiol. Genom. 2003, 13, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Meer, D.L.M.; Van den Thillart, G.E.E.J.M.; Witte, F.; de Bakker, M.A.G.; Besser, J.; Richardson, M.K.; Spaink, H.P.; Leito, J.T.D.; Bagowski, C.P. Gene expression profiling of the long-term adaptive response to hypoxia in the gills of adult zebrafish. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 289, R1512–R1519. [Google Scholar] [CrossRef]
- Baptista, R.B.; Souza-Castro, N.; Almeida-Val, V.M.F. Acute hypoxia up-regulates HIF-1α and VEGF mRNA levels in amazon hypoxia-tolerant oscar (Astronotus ocellatus). Fish Physiol. Biochem. 2016, 42, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wu, F.; Xie, S.; Zhang, L. Acute hypoxia and reoxygenation: Effect on oxidative stress and hypoxia signal transduction in the juvenile yellow catfish (Pelteobagrus fulvidraco). Aquaculture 2021, 531, 735903. [Google Scholar] [CrossRef]
- Pandey, A.; Rajesh, M.; Baral, P.; Sarma, D.; Tripathi, P.H.; Akhtar, M.S.; Ciji, A.; Dubey, M.K.; Pande, V.; Sharma, P.; et al. Concurrent changes in thermal tolerance thresholds and cellular heat stress response reveals novel molecular signatures and markers of high temperature acclimation in rainbow trout. J. Therm. Biol. 2021, 102, 103124. [Google Scholar] [CrossRef]
- Marti, H.H.; Wenger, R.H.; Rivas, L.A.; Straumann, U.; Oigicaylioglu, M.; Henn, V.; Yonekawa, Y.; Bauer, C.; Gassmann, M. Erythropoietin gene expression in human, monkey and murine brain. Eur. J. Neurosci. 1996, 8, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.F.; Tohari, S.; Brenner, S.; Venkatesh, B. Erythropoietin gene from a teleost fish, Fugu rubripes. Blood 2004, 104, 1498–1503. [Google Scholar] [CrossRef] [Green Version]
- Jelkmann, W. Regulation of erythropoietin production. J. Physiol. 2011, 589, 1251–1258. [Google Scholar] [CrossRef]
- Petousi, N.; Croft, Q.P.P.; Cavalleri, G.L.; Cheng, H.-Y.; Formenti, F.; Ishida, K.; Lunn, D.; McCormack, M.; Shianna, K.V.; Talbot, N.P.; et al. Tibetans living at sea level have a hyporesponsive hypoxia-inducible factor system and blunted physiological responses to hypoxia. J. Appl. Physiol. 2014, 116, 893–904. [Google Scholar] [CrossRef] [Green Version]
- Greenald, D.; Jeyakani, J.; Pelster, B.; Sealy, I.; Mathavan, S.; van Eeden, F.J. Genome-wide mapping of HIF-1α binding sites in zebrafish. BMC Genom. 2015, 16, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez-Valls, E.; Flores-Soto, M.E.; Chaparro-Huerta, V.; Torres-Mendoza, B.M.; Gudiño-Cabrera, G.; Rivera-Cervantes, M.C.; Pallas, M.; Camins, A.; Armendáriz-Borunda, J.; Beas-Zarate, C. HIF-1α expression in the hippocampus and peripheral macrophages after glutamate-induced excitotoxicity. J. Neuroimmunol. 2011, 238, 12–18. [Google Scholar] [CrossRef]
- Zhou, J.; Li, C.; Yao, W.; Alsiddig, M.C.; Huo, L.; Liu, H.; Miao, Y.-L. Hypoxia-inducible factor-1α-dependent autophagy plays a role in glycolysis switch in mouse granulosa cells. Biol. Reprod. 2018, 99, 308–318. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Qi, C.; Li, E.; Du, Z.; Qin, J.G.; Chen, L. Metabolic response of nile tilapia (Oreochromis niloticus) to acute and chronic hypoxia stress. Aquaculture 2018, 495, 187–195. [Google Scholar] [CrossRef]
- Sun, J.L.; Zhao, L.L.; Wu, H.; Liu, Q.; Liao, L.; Luo, J.; Lian, W.Q.; Cui, C.; Jin, L.; Ma, J.D.; et al. Acute hypoxia changes the mode of glucose and lipid utilization in the liver of the largemouth bass (Micropterus salmoides). Sci. Total Environ. 2020, 713, 135157. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Liu, C.; Luo, S.; Zhang, Y.; Gao, X.; Wu, X.; Shen, W.; Zhu, J. Transcriptome and physiology analysis identify key metabolic changes in the liver of the large yellow croaker (Larimichthys crocea) in response to acute hypoxia. Ecotoxicol. Environ. Saf. 2020, 189, 109957. [Google Scholar] [CrossRef]
- Ma, F.; Zou, Y.; Ma, L.; Ma, R.; Chen, X. Evolution, characterization, and immune response function of long-chain acyl-coa synthetase genes in rainbow trout (Oncorhynchus mykiss) under hypoxic stress. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2022, 260, 110737. [Google Scholar] [CrossRef]
- Mashek, D.G.; Li, L.O.; Coleman, R.A. Rat long-chain acyl-CoA synthetase mRNA, protein, and activity vary in tissue distribution and in response to diet. J. Lipid Res. 2006, 47, 2004–2010. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhu, H.; Huang, X.; Wang, A.; Xie, D. Molecular characterization, tissue distribution profile, and nutritional regulation of ACSL gene family in golden pompano (Trachinotus ovatus). Int. J. Mol. Sci. 2022, 23, 6437. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Ye, C.X.; Guo, Z.X.; Wang, A.L. Immune and physiological responses of pufferfish (Takifugu obscurus) under cold stress. Fish Shellfish Immun. 2017, 64, 137–145. [Google Scholar] [CrossRef]
- Li, Y.; Pang, Y.; Xiang, X.; Du, J.; Mai, K.; Ai, Q. Molecular cloning, characterization, and nutritional regulation of Elovl6 in large yellow croaker (Larimichthys crocea). Int. J. Mol. Sci. 2019, 20, 1801. [Google Scholar] [CrossRef] [Green Version]
- Geay, F.; Tinti, E.; Mellery, J.; Michaux, C.; Larondelle, Y.; Perpète, E.; Kestemont, P. Cloning and functional characterization of δ6 fatty acid desaturase (FADS2) in eurasian perch (Perca fluviatilis). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 191, 112–125. [Google Scholar] [CrossRef]
- Fadhlaoui, M.; Pierron, F.; Couture, P. Temperature and metal exposure affect membrane fatty acid composition and transcription of desaturases and elongases in fathead minnow muscle and brain. Ecotoxicol. Environ. Saf. 2018, 148, 632–643. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| PFKL | CATGTGTGCTTGGCCTCAAC | GGCCGCAGATTATACCACCA |
| HSP90AB1 | CATTTCTCAGTGGAGGGCCA | GAGATGTTGAGGGGCAGGTC |
| ENO1 | CTGACGACCCTAACCGCTAC | TCACCCACCACCTGGATACT |
| EPO | CTCAGCTTGCTACGGTCCTC | CACCGTCCATGTTCCCTCAA |
| GAPDH | TCGGAGTCGCAAGACAGACA | AACCTTCTTGGAGTGGAAAGCA |
| ALDOCB | TATCCCGCACTGACTCCTGA | TGTGTTCTCAACCCCGATGG |
| MKNK2 | CTGTGGCTGGGAATTAGGGG | GGTTTTTGGCATCCCGAACC |
| PSMB3 | TACATCGAGCCCGTGATTGC | CACACATGCCGTACATCTGC |
| HIF1A | GACACATTGGCATCACGCAG | TGTCTGCGGCTTCTTACTCG |
| PSMC5 | CCCAAGGGTGTGCTGTTGTA | GCACCCTCTCCGATGAACTT |
| PSMD1 | TGGCCTAGCAGTGGGAATTG | AGTGTACATTCCAGAGCGCC |
| Sample | Raw Reads Number | Clean Reads Number | Raw Bases (bp) | Clean Bases (bp) | Error Rate (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|---|
| CT1 | 45,449,954 | 45,392,066 | 6,817,493,100 | 6,779,000,457 | 0.13 | 97.97 | 94.1 | 50.71 |
| CT2 | 44,500,604 | 44,444,572 | 6,675,090,600 | 6,639,750,913 | 0.13 | 98.04 | 94.24 | 50.21 |
| CT3 | 46,646,638 | 46,596,420 | 6,996,995,700 | 6,948,044,496 | 0.11 | 98.02 | 94.18 | 50.19 |
| HT31-1 | 37,526,202 | 37,480,302 | 5,628,930,300 | 5,593,758,917 | 0.12 | 97.81 | 93.71 | 50.67 |
| HT31-2 | 40,355,502 | 40,304,816 | 6,053,325,300 | 6,014,917,041 | 0.13 | 97.88 | 93.92 | 50.12 |
| HT31-3 | 39,904,620 | 39,857,718 | 5,985,693,000 | 5,949,485,580 | 0.12 | 97.99 | 94.16 | 50.78 |
| HT34-1 | 43,571,734 | 43,524,904 | 6,535,760,100 | 6,493,798,692 | 0.11 | 98.11 | 94.38 | 51.23 |
| HT34-2 | 45,659,610 | 45,602,954 | 6,848,941,500 | 6,798,784,900 | 0.12 | 97.94 | 94 | 52.84 |
| HT34-3 | 43,264,302 | 43,214,412 | 6,489,645,300 | 6,448,179,655 | 0.12 | 98.09 | 94.38 | 51.07 |
| Sample | High Quality Clean Reads Number (%) | Unmapped (%) | Unique Mapped (%) | Multiple Mapped (%) | Total Mapped (%) |
|---|---|---|---|---|---|
| CT1 | 45,161,002 (99.49%) | 3,409,069 (7.55%) | 38,298,481 (84.80%) | 3,453,452 (7.65%) | 41,751,933 (92.45%) |
| CT2 | 44,019,362 (99.04%) | 3,303,947 (7.51%) | 37,445,636 (85.07%) | 3,269,779 (7.43%) | 40,715,415 (92.49%) |
| CT3 | 46,521,200 (99.84%) | 2,776,762 (5.97%) | 39,787,437 (85.53%) | 3,957,001 (8.51%) | 43,744,438 (94.03%) |
| HT31-1 | 37,301,710 (99.52%) | 2,670,282 (7.16%) | 31,797,678 (85.24%) | 2,833,750 (7.60%) | 34,631,428 (92.84%) |
| HT31-2 | 40,179,202 (99.69%) | 3,327,393 (8.28%) | 33,573,047 (83.56%) | 3,278,762 (8.16%) | 36,851,809 (91.72%) |
| HT31-3 | 39,515,610 (99.14%) | 3,218,708 (8.15%) | 33,369,322 (84.45%) | 2,927,580 (7.41%) | 36,296,902 (91.85%) |
| HT34-1 | 43,441,596 (99.81%) | 2,913,743 (6.71%) | 37,249,288 (85.75%) | 3,278,565 (7.55%) | 40,527,853 (93.29%) |
| HT34-2 | 41,427,788 (90.84%) | 9,231,820 (22.28%) | 29,895,198 (72.16%) | 2,300,770 (5.55%) | 32,195,968 (77.72%) |
| HT34-3 | 42,779,862 (98.99%) | 3,236,432 (7.57%) | 36,188,522 (84.59%) | 3,354,908 (7.84%) | 39,543,430 (92.43%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.-Q.; Zhang, J.; Wang, H.-Y.; Niu, S.-F.; Wu, R.-X.; Tang, B.-G.; Wang, Q.-H.; Liang, Z.-B.; Liang, Y.-S. Transcriptomic Response of the Liver Tissue in Trachinotus ovatus to Acute Heat Stress. Animals 2023, 13, 2053. https://doi.org/10.3390/ani13132053
Li Q-Q, Zhang J, Wang H-Y, Niu S-F, Wu R-X, Tang B-G, Wang Q-H, Liang Z-B, Liang Y-S. Transcriptomic Response of the Liver Tissue in Trachinotus ovatus to Acute Heat Stress. Animals. 2023; 13(13):2053. https://doi.org/10.3390/ani13132053
Chicago/Turabian StyleLi, Qian-Qian, Jing Zhang, Hong-Yang Wang, Su-Fang Niu, Ren-Xie Wu, Bao-Gui Tang, Qing-Hua Wang, Zhen-Bang Liang, and Yan-Shan Liang. 2023. "Transcriptomic Response of the Liver Tissue in Trachinotus ovatus to Acute Heat Stress" Animals 13, no. 13: 2053. https://doi.org/10.3390/ani13132053