1. Introduction
Whiteleg shrimp,
Litopenaeus vannamei, is probably the most lucrative species in aqua culture, with 5.8 million metric tons of production in 2020 accounting for a value of approximately USD 34 billion [
1]. To satisfy the global market demands for this species, sustainable development of shrimp aquaculture industry is highly required. Challenges such as excessive use of marine ingredients (e.g., fishmeal and fish oil) in diet, high mortality due to improper nutrition and excessive feed prices are major impediments in shrimp aquaculture [
2,
3].
Fishmeal and fish oil are among the best protein and lipid sources in shrimp diets. Fishmeal contains sufficient amounts of essential amino acids and possesses high digestibility and palatability in shrimp nutrition [
4,
5,
6,
7]. However, fishmeal is not sustainably sourced, not available in required quantities and not cheap. To meet the growing demands for shrimp feed, several researchers have examined the use of alternative protein sources in shrimp diet production. These researchers reported the successful use of ingredients originating from plant [
8], animal [
9,
10] and microbial sources [
11] as alternatives or complements of fishmeal in shrimp diets. Microbial protein is obtained from different microbial biomass sources such as yeast bacteria and microalgae, with great potential to partially or even completely replace fishmeal in shrimp diets [
12].
Yeast products obtained from
Saccharomyces cerevisiae (brewer’s yeast) have been used in animal and human diets for decades [
13,
14]. The rich protein, amino acid and energy content of yeast makes it a suitable ingredient for aquatic species including whiteleg shrimp [
15,
16,
17]. Furthermore, yeast is a rich source of nucleotides, beta-glucans, mannan oligosaccharides, chitin and peptides [
12,
18]. These compounds have shown to improve immune response, growth performance and stress resistance in whiteleg shrimp [
19,
20,
21,
22,
23]. However, the yeast cell wall is not easily digested by aquatic animals and low digestibility values are often observed when unprocessed yeast is used in shrimp diets [
17,
24]. Low digestibility can result in poor utilization of yeast nutrients and active compounds by the animal. Another important factor is the price of yeast products after processing. Practical incorporation of yeast products in shrimp diet requires thorough consideration of economic feasibility. With a price almost identical to fishmeal, yeast hydrolysate improved the growth performance and feed utilization of whiteleg shrimp fed a diet with 20% fishmeal [
22]. Moreover, improved defense against low salinity stress by modulating superoxide dismutase, alkaline phosphatase and acid phosphatase activities was observed in shrimp fed 3% yeast hydrolysate [
23]. Nevertheless, a dose-dependent response of whiteleg shrimp to dietary yeast hydrolysate in low-fishmeal diets has not been investigated. Therefore, based on previous literature, the present study aimed at evaluating the effects of graded levels of yeast hydrolysate in low-fishmeal diets (10%) on growth, body composition, non-specific immune responses, hematological parameters, intestinal morphology and disease resistance in juvenile whiteleg shrimp
L. vannamei.
4. Discussion
In the present experiment, the positive control diet was 25% fishmeal, which is comparable to shrimp commercial diets [
15,
22]. Other experimental diets consisted of 10% fishmeal which is considered “low” in Asian shrimp aquaculture. Previous studies have shown the great potential of yeast products in low-fishmeal diets for whiteleg shrimp [
15,
17,
18,
24]. Additionally, the positive effects of yeast hydrolysate on growth, immune responses, antioxidant capacity, stress resistance and gene expression of whiteleg shrimp was proven in previous studies [
22,
23]. However, to the best of our knowledge, a dose-dependent response of dietary yeast hydrolysate in low-fishmeal diets for whiteleg shrimp has not been investigated.
In the present experiment, the FBW, WG, SGR and PER of shrimp fed the positive control diet (25% fishmeal) were higher than all other diets, except for the diet with 4% yeast hydrolysate. This shows that the highest level (4%) of yeast hydrolysate could have compensated for the nutritional insufficiencies in the low-fishmeal diets. In a study, it was shown that fishmeal can be reduced from 25% to 20% in whiteleg shrimp diet when 3% of yeast extract is used [
22]. A straightforward explanation for these observations could be the rich nutritional composition of yeast products, as was shown in several previous studies [
29]. Yeast products, especially those obtained from
Saccharomyces cerevisiae, have a relatively high protein content, balanced amino acid composition and a rich amount of nucleotide [
29]. Nucleotides are intracellular compounds that build up nucleic acids and have key roles in a wide range of biochemical processes [
30]. Fishmeal itself is a rich source of nucleotides and replacement of fishmeal may result in a significant drop in nucleotides, as well as essential amino acids [
31]. Although nucleotides are not essential nutrients, they play critical roles in feeding attraction and stimulation, growth promotion, immunostimulantion, stress responses and disease resistance [
32]. It was reported that dietary nucleotides (0.9%) enhanced immune response, gastrointestinal health and disease resistance in whiteleg shrimp fed a low-fishmeal diet [
33]. The yeast hydrolysate used in our experiment contains 12.17% of nucleic acids [
16], and this can partly explain our growth enhancement results. Another explanation could be related to the high digestibility of yeast products in shrimp diet. Yeast products have a relatively high digestibility in shrimp diet that could be related to the quality of the protein [
15,
17]. Nevertheless, further studies with higher levels of yeast hydrolysate in shrimp diet are required to observe linear additive effects.
Shrimp whole-body proximate composition was not significantly affected as a response to different dietary treatments in this study. Similar to our results, differences in body protein content of whiteleg shrimp when fed different levels of yeast and yeast extract were not observed [
18]. However, an increase in body crude lipid content with higher yeast supplementations was reported [
18], which contradicts our results. In our study, the whole-body crude lipid content of shrimp fed increasing levels of yeast hydrolysate showed a slight increase compared to the negative control group, but differences were not significant. This might be due to the fact that shrimp muscle contains low levels of fat compared to fish [
34], and therefore differences might be negligible. Similar to the body composition results, the hemolymph biochemical parameters, such as aspartate aminotransferase (AST), alanine aminotransferase (ALT) and glucose, were not affected by dietary yeast hydrolysate and/or low fishmeal. AST and ALT are enzymes that are used as indicators of liver health and function [
35] and serum glucose is an indicator of stress in aquatic animals [
36]. A previous study has shown that suboptimal conditions such as induction of hypothermal stress have resulted in increased glucose levels in shrimp hemolymph [
37]. On the contrary, optimum levels of dietary yeast nucleotides can reduce the glucose levels in fish after acute stress [
38]. The stagnant AST, ALT and glucose levels in our experiment might be because the nutrient insufficiencies or experimental conditions were not severe enough to damage the shrimp hepatopancreas. Future studies are needed with a focus on the effects of yeast hydrolysate on hemolymph parameters in shrimp while a stress challenge is induced.
Superoxide dismutase (SOD) enzyme is the part of the antioxidant defense mechanisms that scavenges the superoxide anion (O
2−), and lysozyme is an antimicrobial enzyme that can lyse cell membranes of pathogenic bacteria. SOD and lysozyme are two major indicators of innate immune responses [
39,
40]. Our results showed no significant differences in SOD and lysozyme activities of shrimp hemolymph fed different levels of yeast hydrolysate in low-fishmeal diets. Similar to our results, no changes were observed in SOD activity when 3% of yeast extract was used to replace concentrated cottonseed protein in whiteleg shrimp [
18]. In contrast, in another study, it was reported that dietary yeast hydrolysate increased the SOD activity of shrimp when salinity stress was induced [
23]. It seems that the effects of dietary yeast products on immune responses are more obvious when extreme challenges are exposed. In fish fed graded levels of yeast hydrolysate, it was notably shown that plasma lysozyme, SOD and IgM were only significant after the pathogenic challenge test [
16]; these parameters were not significantly affected by dietary yeast hydrolysate before the challenge test. This might indicate that the positive effects of dietary yeast hydrolysate are more visible when fish/shrimp are challenged or stressed.
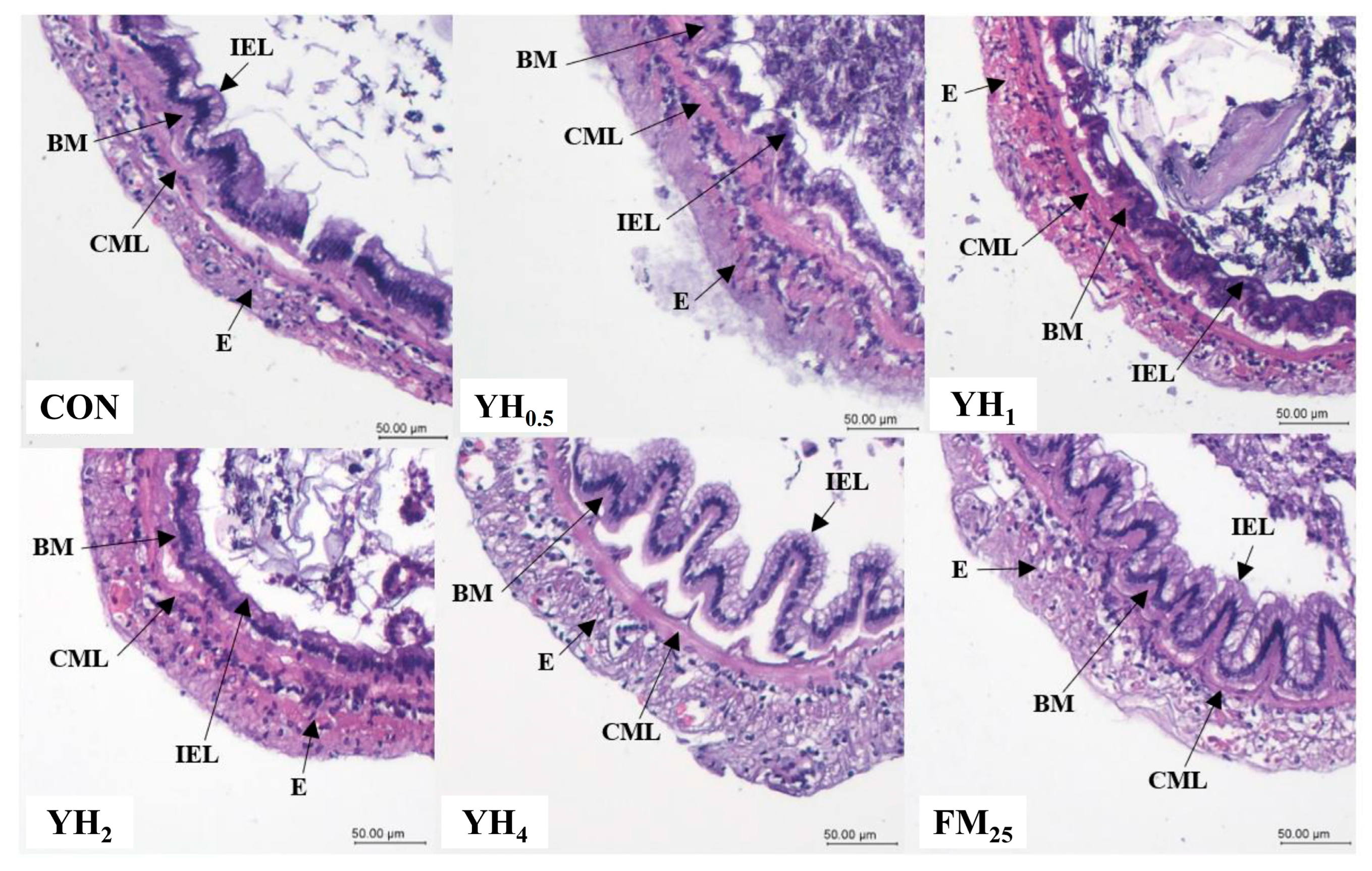
The intestinal morphology of aquatic organisms directly reflects the quality of feed and incorporated ingredients. For example, high levels of unprocessed soybean meal is known to induce intestinal inflammation in fish [
41]. Moreover, intestinal morphological status such as villi length, muscular layer thickness and number of goblet cells play a significant role in digestion and absorption of nutrients [
42]. The results of the present experiment have shown a clear and significant increase in intestinal villi length and muscular layer thickness when 4% yeast hydrolysate was used in shrimp diet. This can partly explain our observations in growth performance parameters. In line with our findings, it was reported that 1.5% dietary baker’s yeast extract improved the structure of hepatocytes and villi in fish [
43]. It was also shown that dietary nucleotides, glucans and mannan oligosaccharides, which are relatively abundant in yeast hydrolysate, enhance intestinal morphometric characteristics [
30,
44]. Other than this, we observed a decrease in villi length in group YH
2 compared to YH
1 that does not seem to have a clear explanation, and further studies in this regard are required.
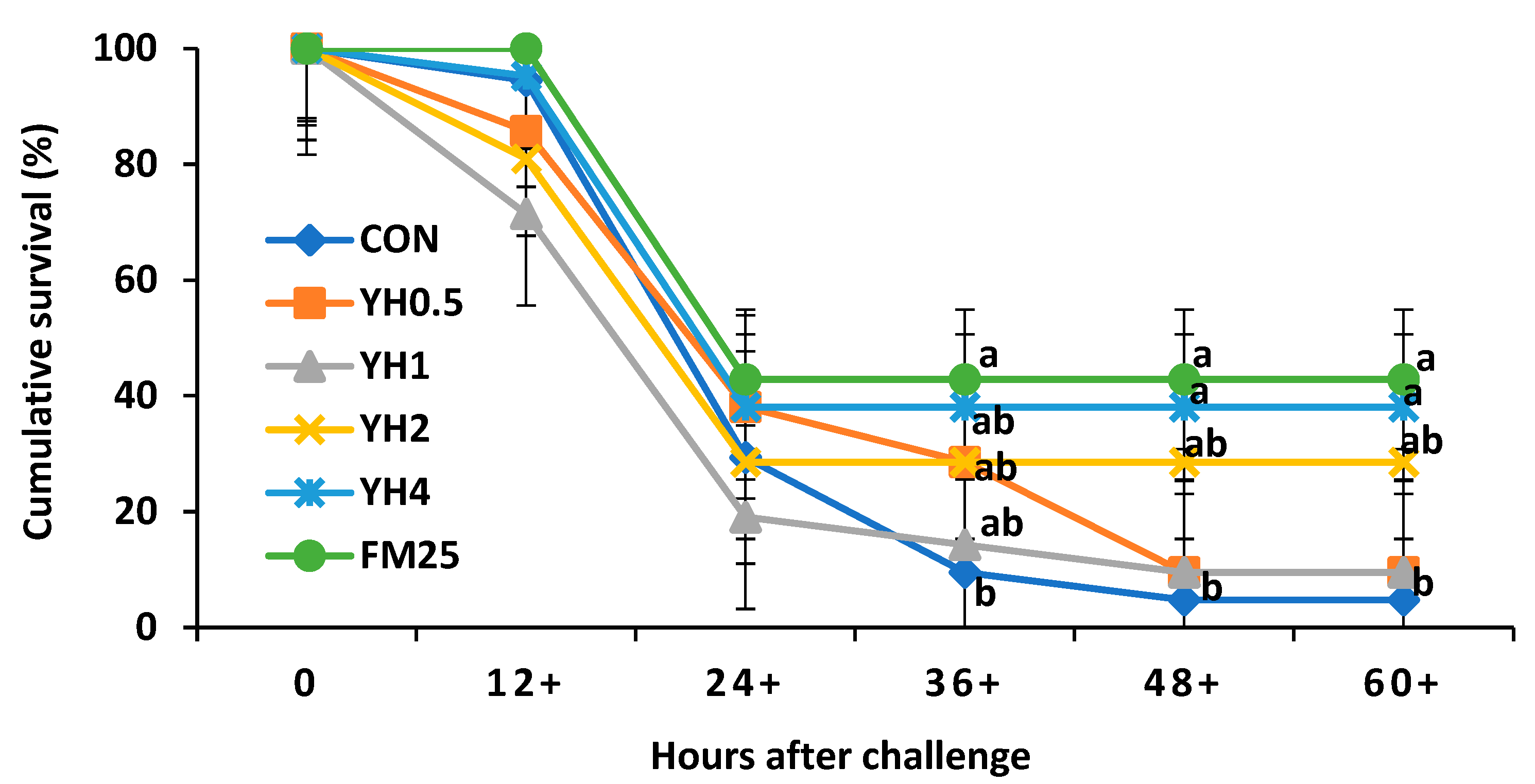
Whiteleg shrimp fed the positive control and 4% yeast hydrolysate showed the highest cumulative survival after the challenge with
Vibrio parahaemolyticus, but with no significant difference with the YH
2 group. Vibriosis is caused by
Vibrio parahaemolyticus, which is a common disease in shrimp farms leading to high mortality and substantial economic losses [
44]. One of the greatest challenges in shrimp farming is the prevention and control of disease outbreaks. Shrimp, unlike teleost fish, lack adaptive immune mechanisms and this highlights the importance of innate immunity [
45,
46]. Yeast and its derivatives have previously been shown to improve the innate immune responses of shrimp, leading to higher survival against
Vibrio parahaemolyticus [
47,
48,
49]. This could be explained by the presence of compounds such as ß–glucan and nucleotides in yeast. These compounds are well-known for their stimulatory effects on the innate immune responses of aquatic animals [
30,
50]. Nevertheless, as mentioned before, our results on lysozyme and SOD activity did not show significant differences as a response to dietary yeast hydrolysate. Moreover, we did not observe the same trends in shrimp survival percentage during the growth trial. It could be possible that the slight (non-significant) differences in shrimp survival during the growth trial were caused by non-nutritional (environmental) factors. Additionally, according to a previous study [
16] and based on the results of the present experiment, it seems that the positive effects of yeast products are more obvious at suboptimal conditions and/or after bacterial challenge. Further studies are required on survival and the changes in innate immune responses of shrimp after bacterial challenge tests.

 ,
, 


{kind=link}
{kind=link}