
Validation of Calcein Violet as a New Marker of Semen Membrane Integrity in Domestic Animals
 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Semen Collection
2.2. Semen Evaluation
2.3. Semen Dilution and Samples Preparation
2.3.1. CaV Validation as a Marker of Viability (Experiment 1)
2.3.2. CaV Validation as a Marker of Acrosomal Membrane Integrity (Experiment 2)
2.4. Fluorochromes and Epifluorescence Evaluation
2.4.1. CaV Validation as a Marker of Viability (Experiment 1)
2.4.2. CaV Validation as a Marker of Acrosomal Membrane Integrity (Experiment 2)
2.5. Statistical Analysis
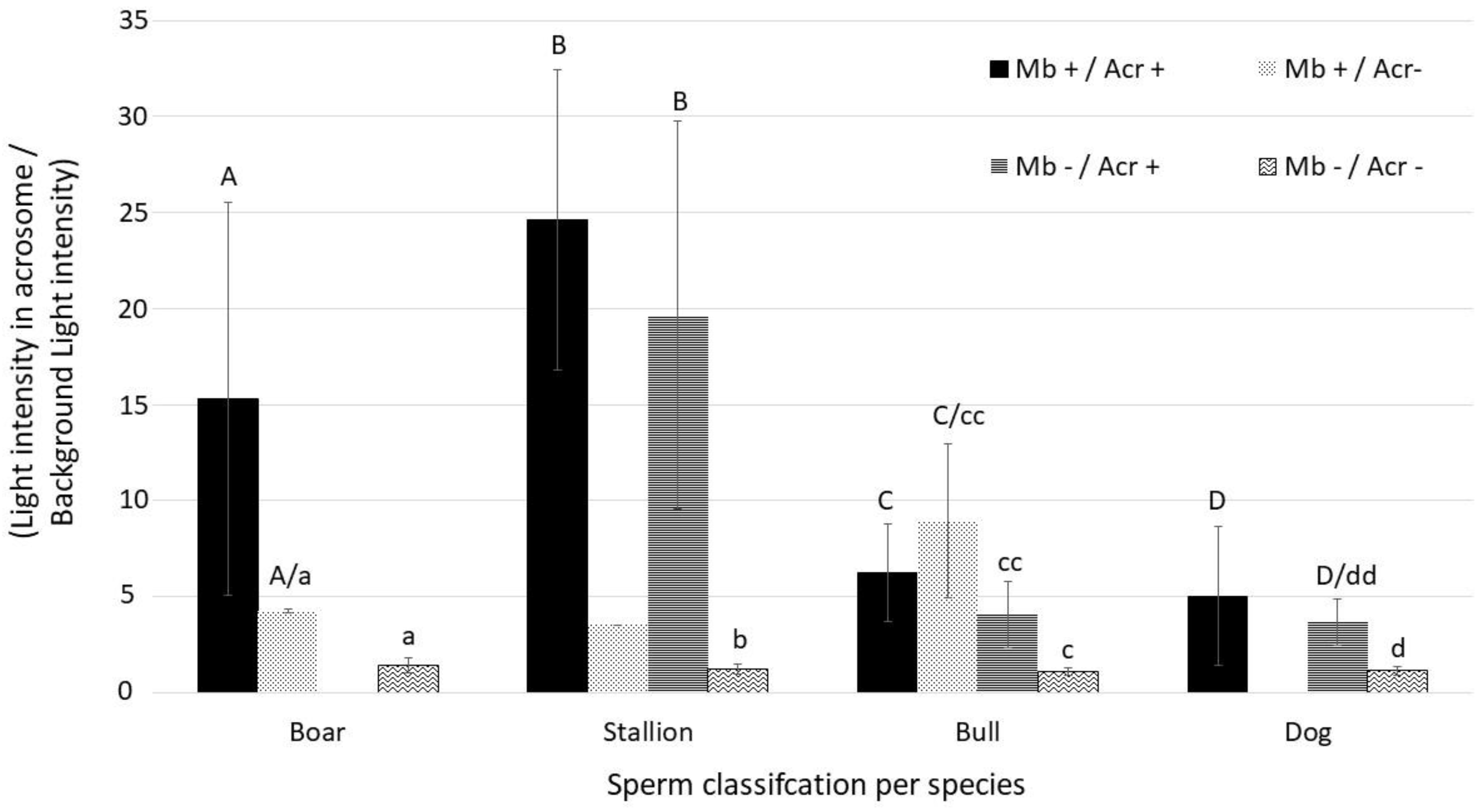
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vincent, P.; Underwood, S.L.; Dolbec, C.; Bouchard, N.; Kroetsch, T.; Blondin, P. Bovine semen quality control in artificial insemination centers. Anim. Reprod. 2012, 9, 153–165. [Google Scholar] [CrossRef]
- Gliozzi, T.M.; Turri, F.; Manes, S.; Cassinelli, C.; Pizzi, F. The combination of kinetic and flow cytometric semen parameters as a tool to predict fertility in cryopreserved bull semen. Animal 2017, 11, 1975–1982. [Google Scholar] [CrossRef] [PubMed]
- Bucher, K.; Malama, E.; Siuda, M.; Janett, F.; Bollwein, H. Multicolor flow cytometric analysis of cryopreserved bovine sperm: A tool for the evaluation of bull fertility. J. Dairy Sci. 2019, 102, 11652–11669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nizański, W.; Partyka, A.; Rijsselaere, T. Use of fluorescent stainings and flow cytometry for canine semen assessment. Reprod. Domest. Anim. 2012, 47, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Ibanescu, I.; Siuda, M.; Bollwein, H. Motile sperm subpopulations in bull semen using different clustering approaches—Associations with flow cytometric sperm characteristics and fertility. Anim. Reprod. Sci. 2020, 215, 106329. [Google Scholar] [CrossRef]
- Jung, M.; Rüdiger, K.; Schulze, M. In Vitro Measures for Assessing Boar Semen Fertility. Reprod. Domest. Anim. 2015, 50, 20–24. [Google Scholar] [CrossRef]
- Wegener, J.; May, T.; Knollmann, U.; Kamp, G.; Müller, K.; Bienefeld, K. In vivo validation of in vitro quality tests for cryopreserved honey bee semen. Cryobiology 2012, 65, 126–131. [Google Scholar] [CrossRef]
- Farrell, P.B.; Presicce, G.A.; Brockett, C.C.; Foote, R.H. Quantification of bull sperm characteristics measured by computer-assisted sperm analysis (CASA) and the relationship to fertility. Theriogenology 1998, 49, 871–879. [Google Scholar] [CrossRef]
- Samper, J.C.; Hellander, J.C.; Crabo, B.G. Relationship between the fertility of fresh and frozen stallion semen and semen quality. J. Reprod. Fertil. Suppl. 1991, 44, 107–114. [Google Scholar]
- Holt, C.; Holt, W.V.; Moore, H.D.M.; Reed, H.C.B.; Curnock, R.M. Objectively Measured Boar Sperm Motility Parameters Correlate with the Outcomes of On-Farm Inseminations: Results of Two Fertility Trials. J. Androl. 1997, 18, 312–323. [Google Scholar] [CrossRef]
- Froman, D.P.; McLean, D.J. Objective measurement of sperm motility based upon sperm penetration of accudenz®. Poult. Sci. 1996, 75, 776–784. [Google Scholar] [CrossRef]
- Tesi, M.; Sabatini, C.; Vannozzi, I.; Di Petta, G.; Panzani, D.; Camillo, F.; Rota, A. Variables affecting semen quality and its relation to fertility in the dog: A retrospective study. Theriogenology 2018, 118, 34–39. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; Björndahl, L., Apolkhin, O., Baldi, E., Barratt, C., Festin, M., Kirkman-Brown, J., Lamb, D., Mbizvo, M., Schlatt, S., Toskin, I., et al., Eds.; World Health Organization: Geneva, Switzerland, 2021; Volume 5, ISBN 978-9-24154-778-9. [Google Scholar]
- Farah, O.I.; Cuiling, L.; Jiaojiao, W.; Zh, H. Use of Fluorescent Dyes for Readily Recognizing Sperm Damage. J. Reprod. Infertil. 2013, 14, 120–125. [Google Scholar] [PubMed]
- Nagy, S.; Jansen, J.; Topper, E.K.; Gadella, B.M. A Triple-Stain Flow Cytometric Method to Assess Plasma-and Acrosome-Membrane Integrity of Cryopreserved Bovine Sperm Immediately after Thawing in Presence of Egg-Yolk Particles 1. Biol. Reprod. 2003, 68, 1828–1835. [Google Scholar] [CrossRef] [PubMed]
- Petrunkina, A.; Harrison, R. Fluorescence technologies for evaluating male gamete (dys)function. Reprod. Domest. Anim. 2013, 48, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Arslan, H.O.; Herrera, C.; Malama, E.; Siuda, M.; Leiding, C.; Bollwein, H. Effect of the addition of different catalase concentrations to a TRIS-egg yolk extender on quality and in vitro fertilization rate of frozen-thawed bull sperm. Cryobiology 2019, 91, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L. Acetylcholinesterase in bull spermatozoa. J. Reprod. Fertil. 1964, 7, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Bryan, J.H.D.; Unnithan, R.R. Non-specific esterase activity in bovine acrosomes. Histochem. J. 1972, 4, 413–419. [Google Scholar] [CrossRef]
- Bradford, M.M.; McRorie, R.A.; Williams, W.L. A role for esterases in the fertilization process. J. Exp. Zool. 1976, 197, 297–301. [Google Scholar] [CrossRef]
- Meizel, S.; Boggs, D.; Cotham, J. Electrophoretic studies of esterases of bull spermatozoa, cytoplasmic droplets and seminal plasma. J. Histochem. Cytochem. 1971, 19, 226–231. [Google Scholar] [CrossRef]
- Salazar, P.; Graham, J.K.; Parrish, J.J.; Susko-Parrish, J.; Squires, E.L. Indirect determination of stallion sperm capacitation based on esterase release from spermatozoa challenged with lysophosphatidylcholine. J. Reprod. Fertil. Suppl. 2000, 56, 407–414. [Google Scholar]
- Graham, J.K.; Mocé, E. Fertility evaluation of frozen/thawed semen. Theriogenology 2005, 64, 492–504. [Google Scholar] [CrossRef]
- Sirivaidyapong, S.; Cheng, F.P.; Marks, A.; Voorhout, W.F.; Bevers, M.M.; Colenbrander, B. Effect of sperm diluents on the acrosome reaction in canine sperm. Theriogenology 2000, 53, 789–802. [Google Scholar] [CrossRef]
- Fazeli, A.; Hage, W.J.; Cheng, F.P.; Voorhout, W.F.; Marks, A.; Bevers, M.M.; Colenbrander, B. Acrosome-intact boar spermatozoa initiate binding to the homologous zona pellucida in vitro. Biol. Reprod. 1997, 56, 430–438. [Google Scholar] [CrossRef]
- Cross, N.L.; Watson, S.K. Assessing acrosomal status of bovine sperm using fluoresceinated lectins. Theriogenology 1994, 42, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.-P.; Fazeli, A.; Voorhout, W.F.; Marks, A.; Bevers, M.M.; Colenbrander, B. Use of Peanut to Assess the Acrosomal Status and the Zona Pellucida-Induced Acrosome Reactin in Stallion Spermatozoa. J. Androl. 1996, 17, 674–682. [Google Scholar] [CrossRef]
- Srivastava, N.; Pande, M. Determination of Capacitation-Related changes using fluorescent stains. In Protocols in Semen Biology (Comparing Assays); Srivastava, N., Pande, M., Eds.; Springer: Singapore, 2017; pp. 185–203. ISBN 978-981-10-5199-9. [Google Scholar]
- Paynter, E.; Baer-imhoof, B.; Linden, M.; Lee-pullen, T.; Heel, K.; Rigby, P.; Baer, B. Flow Cytometry as a Rapid and Reliable Method to Quantify Sperm Viability in the Honeybee Apis mellifera. Cytometry. 2014, 85, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Paillard, M.; Rousseau, A.; Giovenazzo, P.; Bailey, J.L. Preservation of Domesticated Honey Bee (Hymenoptera: Apidae) Drone Semen. J. Econ. Entomol. 2017, 110, 1412–1418. [Google Scholar] [CrossRef]
- Yániz, J.; Palacín, I.; Santolaria, P. Effect of chamber characteristics, incubation, and diluent on motility of honey bee (Apis mellifera) drone sperm. Apidologie 2019, 50, 472–481. [Google Scholar] [CrossRef]
- Vidament, M.; Magistrini, M.; Le Foll, Y.; Levillain, N.; Yvon, J.M.; Duchamp, G.; Blesbois, E. Temperatures from 4 to 15 °C are suitable for preserving the fertilizing capacity of stallion semen stored for 22 h or more in INRA96 extender. Theriogenology 2012, 78, 297–307. [Google Scholar] [CrossRef]
- Deleuze, S.; Brutinel, F.; Ponthier, J.; Egyptien, S. Evaluation of short-term storage of canine semen at room temperature. In Reproduction in Domestic Animals; Constantinos, B., Aristotelis, G.L., Eds.; NLM (Medline): Thessaloniki, Greece, 2022; Volume 57, p. 51. [Google Scholar]
- Ahmad, M.; Ahmad, N.; Riaz, A.; Anzar, M. Sperm survival kinetics in different types of bull semen: Progressive motility, plasma membrane integrity, acrosomal status and reactive oxygen species generation. Reprod. Fertil. Dev. 2015, 27, 784–793. [Google Scholar] [CrossRef]
- Bresciani, C.; Bianchera, A.; Bettini, R.; Buschini, A.; Marchi, L.; Cabassi, C.S.; Sabbioni, A.; Righi, F.; Mazzoni, C.; Parmigiani, E. Long-term liquid storage and reproductive evaluation of an innovative boar semen extender (Formula12®) containing a non-reducing disaccharide and an enzymatic agent. Anim. Reprod. Sci. 2017, 180, 10–16. [Google Scholar] [CrossRef]
- Lake, P.; Ravie, O. An exploration of cryoprotective compounds for fowl spermatozoa. Br. Poult. Sci. 1984, 25, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Hermansson, U.; Johannisson, A.; Axnér, E. Cryopreservation of dog semen in a Tris extender with two different 1% soybean preparations compared with a Tris egg yolk extender. Vet. Med. Sci. 2021, 7, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, B.K.; Herr, C.; Sheppard, W.S. Sequential generations of honey bee (Apis mellifera) queens produced using cryopreserved semen. Reprod. Fertil. Dev. 2012, 24, 1079–1083. [Google Scholar] [CrossRef]
- LIVE/DEADTM Sperm Viability Kit. Available online: https://www.thermofisher.com/order/catalog/product/L7011 (accessed on 15 December 2021).
- Janosikova, M.; Petricakova, K.; Ptacek, M.; Savvulidi, F.G.; Rychtarova, J.; Fulka, J.J. New approaches for long-term conservation of rooster spermatozoa. Poult. Sci. 2023, 102, 102386. [Google Scholar] [CrossRef] [PubMed]
- Lensky, Y.; Ben-david, E.; Schindler, H. Ultrastructure of the Spermatozoon of the Mature Drone Honeybee. J. Apic. Res. 1979, 18, 264–271. [Google Scholar] [CrossRef]
- Chłopik, A.; Wysokińska, A. Canine spermatozoa—What do we know about their morphology and physiology? An overview. Reprod. Domest. Anim. 2020, 55, 113–126. [Google Scholar] [CrossRef]
- Hancock, J.L. The morphology of Boar Spermatozoa. J. Microsc. 1956, 76, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, H.B.; Zülkadir, U. The Correlation Between Bull Sperm Head Dimensions and Mitochondrial Helix Length. J. Anim. Vet. Adv. 2010, 9, 1169–1172. [Google Scholar] [CrossRef]
- Gravance, C.G.; Liu, I.K.M.; Davis, R.O.; Hughes, J.P.; Casey, P.J. Quantification of normal head morphometry of stallion spermatozoa. J. Reprod. Fertil. 1996, 108, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.C.; DeRuiter, M.C. Fluorescent Nuclei Measurements Macro (FNMM), a tool for automated cell quantification in ImageJ. Softw. Impacts 2020, 6, 100030. [Google Scholar] [CrossRef]
- Muratori, M.; Forti, G.; Baldi, E. Comparing Flow Cytometry and Fluorescence Microscopy for Analyzing Human Sperm DNA Fragmentation by TUNEL Labeling. Cytometry 2008, 73, 785–787. [Google Scholar] [CrossRef]
- Munkhbat, B.; Wersäll, M.; Baranov, D.G.; Antosiewicz, T.J.; Shegai, T. Suppression of photo-oxidation of organic chromophores by strong coupling to plasmonic nanoantennas. Sci. Adv. 2018, 4, eaas9552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probes, M. SYBRTM Green I. Available online: https://www.thermofisher.com/order/catalog/product/S7585 (accessed on 29 March 2022).
- Love, C.C.; Thompson, J.A.; Brinsko, S.P.; Rigby, S.L.; Blanchard, T.L.; Lowry, V.K.; Varner, D.D. Relationship between stallion sperm motility and viability as detected by two fluorescence staining techniques using flow cytometry. Theriogenology 2003, 60, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Špaleková, E.; Makarevich, A.; Kubovičová, E.; Olexíková, L.; Pivko, J. Relationship between motility and viability parameters of frozen-thawed bull spermatozoa. J. Microbiol. Biotechnol. Food Sci. 2013, 2, 1580–1591. [Google Scholar]
- Reis, L.S.L.S.; Ramos, A.A.; Camargos, A.S.; Oba, E. Integrity of the plasma membrane, the acrosomal membrane, and the mitochondrial membrane potential of sperm in Nelore bulls from puberty to sexual maturity. Arq. Bras. Med. Vet. Zootec. 2016, 68, 620–628. [Google Scholar] [CrossRef]
- Garner, D.L.; Johnson, L.A. Viability Assessment of Mammalian Sperm Using SYBR-14 and Propidium Iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Câmara, D.R.; Ibanescu, I.; Siuda, M.; Bollwein, H. Mitoquinone does not improve sperm cryo-resistance in bulls. Reprod. Domest. Anim. 2022, 57, 10–18. [Google Scholar] [CrossRef]
- Keles, E.; Malama, E.; Bozukova, S.; Siuda, M.; Wyck, S.; Witschi, U.; Bauersachs, S.; Bollwein, H. The micro-RNA content of unsorted cryopreserved bovine sperm and its relation to the fertility of sperm after sex-sorting. BMC Genom. 2021, 22, 30. [Google Scholar] [CrossRef]
- Sung Woo, K.; Jae-Yeong, L.; Chan-Lan, K.; Yeong Gyu, K.; Bongki, K. Evaluation of rooster semen quality using CBB dye based staining method. J. Anim. Reprod. Biotechnol. 2022, 37, 55–61. [Google Scholar] [CrossRef]
- Angrimani, D.S.R.; Bicudo, L.C.; Luceno, N.L.; Leemans, B.; Nichi, M.; Vannucchi, C.I.; Van Soom, A. A triple stain method in conjunction with an in-depth screening of cryopreservation effects on post-thaw sperm in dogs. Cryobiology 2022, 105, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Yániz, J.L.; Silvestre, M.A.; Santolaria, P. Sperm quality assessment in honey bee drones. Biology 2020, 9, 174. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PI Nucleus (RED) | SYBR Nucleus (GREEN) | CaV Cytoplasm (BLUE) | PE-PNAacrosome (ORANGE) | CaV Acrosome (BLUE) | |
|---|---|---|---|---|---|
| Intact plasma membrane/intact acrosome (Mb+/Acr+) | - | + | + | - | + |
| Intact plasma membrane/damaged acrosome (Mb+/Acr-) | - | + | + | + | - |
| Damaged plasma membrane/intact acrosome (Mb-/Acr+) | + | - | - | - | + |
| Damaged plasma membrane/damaged acrosome (Mb-/Acr-) | + | - | - | + | - |
| Species | Motility (%) | Progressive Motility (%) | Concentration (×106/mL) |
|---|---|---|---|
| Boar | 84 ±12 | 49 ±12 | 184.1 ±32 |
| Bull | 39 ±9 | 29 ±10 | 87 ±47.9 |
| Dog | 95 ±0 | 87 ±3 | 623.6 ±210.8 |
| Stallion | 22 ±17 | 16 ±15 | 177 ±114 |
| Drone | 25 ±0 | Nonevaluated | 4800 ±3394.1 |
| Rooster | 68 ±38 | 28 ±4 | 214 ±192.3 |
| Species | PE-PNA - | CaV + |
|---|---|---|
| Boar | 89.9 ±7.2 | 89.9 ±7.2 * |
| Bull | 74 ±8.1 | 68.7 ±17.2 * |
| Dog | 87.5 ±21.7 | 87.5 ±21.7 * |
| Stallion | 84.7 ±5.1 | 71.8 ±17.4 * |
| Non-Heat-Treated Samples | Heat-Treated Samples | ||||
|---|---|---|---|---|---|
| Species | SYBR+ PI- | CaV + | Species | SYBR+ PI- | CaV + |
| Boar | 62.4 ±11.4 | 62.4 ±11.4 * | Boar | 0 ±0 | 0 ±0 |
| Bull | 51.7 ±12/0.2 | 48.9 ±9.4 * | Bull | 0.6 ±1.1 | 0 ±0 |
| Dog | 73.8 ±5.5 | 72.4 ±4.8 * | Dog | 0 ±0 | 0 ±0 |
| Stallion | 30.2 ±20.9 | 29.8 ±21 * | Stallion | 0 ±0 | 0 ±0 |
| Drone | 66.3 ±5.7 | 66.3 ±5.7 * | Drone | 6.5 ±0.7 | 0 ±0 |
| Rooster | 63.6 ±7.4 | 63.6 ±7.4 * | Rooster | 9.5 ±0.7 | 0 ±0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egyptien, S.; Dewals, B.; Ectors, F.; Brutinel, F.; Ponthier, J.; Deleuze, S. Validation of Calcein Violet as a New Marker of Semen Membrane Integrity in Domestic Animals. Animals 2023, 13, 1874. https://doi.org/10.3390/ani13111874
Egyptien S, Dewals B, Ectors F, Brutinel F, Ponthier J, Deleuze S. Validation of Calcein Violet as a New Marker of Semen Membrane Integrity in Domestic Animals. Animals. 2023; 13(11):1874. https://doi.org/10.3390/ani13111874
Chicago/Turabian StyleEgyptien, Sophie, Benjamin Dewals, Fabien Ectors, Flore Brutinel, Jérôme Ponthier, and Stéfan Deleuze. 2023. "Validation of Calcein Violet as a New Marker of Semen Membrane Integrity in Domestic Animals" Animals 13, no. 11: 1874. https://doi.org/10.3390/ani13111874