
Simultaneous Occurrence of Hypospadias and Bilateral Cleft Lip and Jaw in a Crossbred Calf: Clinical, Computer Tomographic, and Genomic Characterization
 , ,
, ,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Statement of Ethics
2.2. Animal
2.3. Computer Tomography (CT) Analyzes
2.4. Whole Genome Sequencing
3. Results
3.1. Clinical Findings
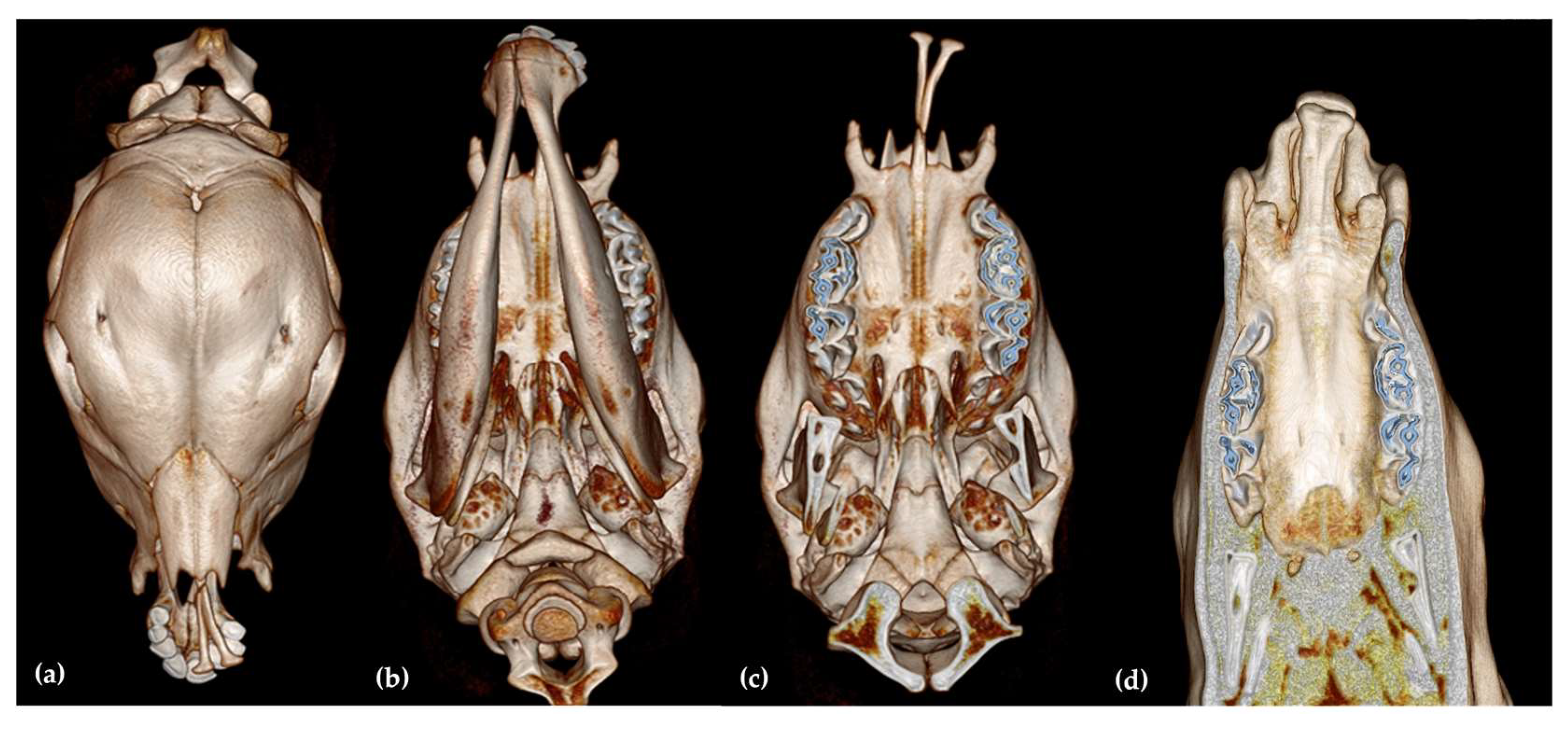
3.2. Computed Tomography Findings
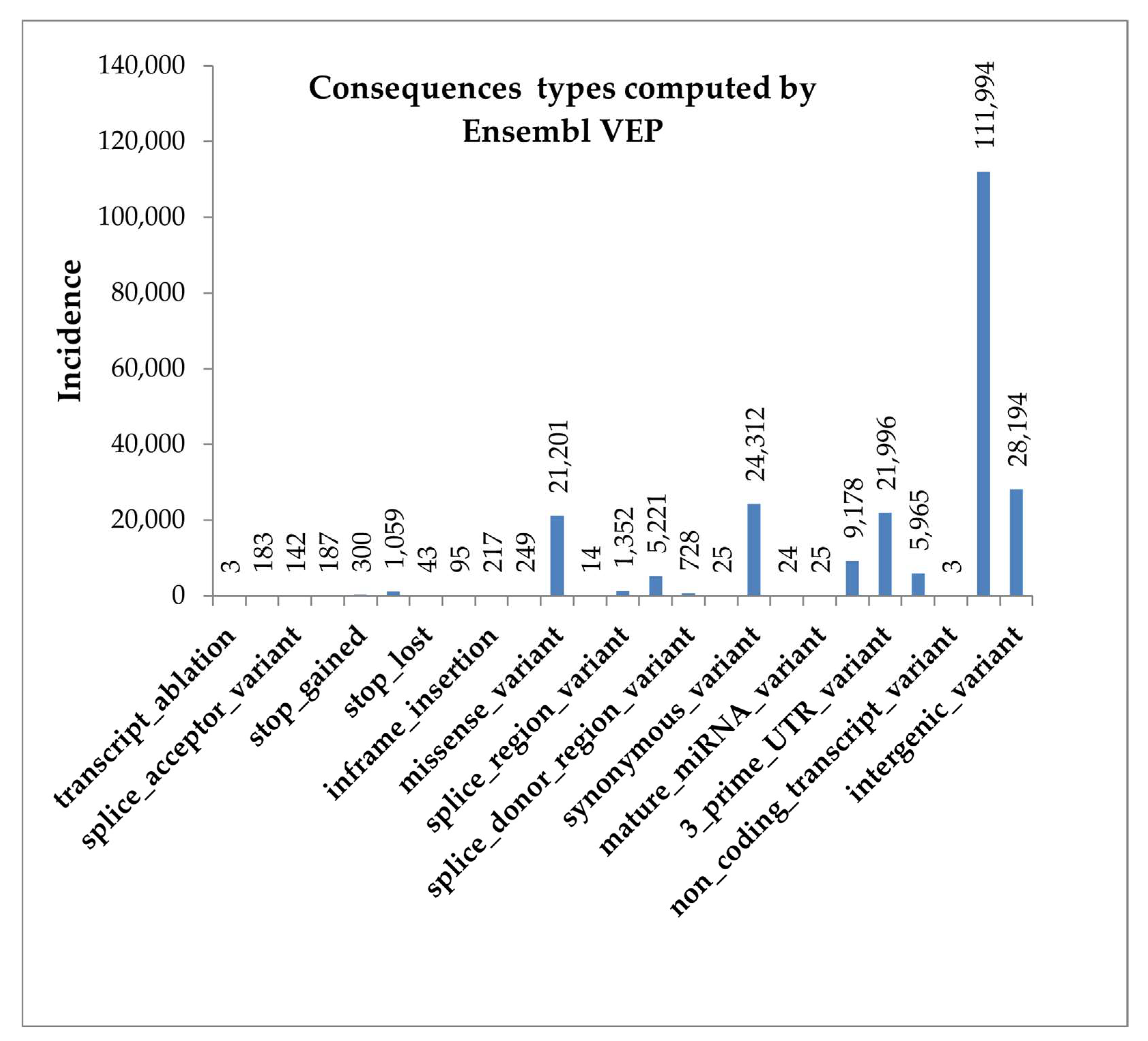
3.3. Whole Genome Sequencing Findings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGeady, T.A.; Quinn, P.J.; FitzPatrick, E.S.; Ryan, M.T.; Kilroy, D.; Lonergan, P. Veterinary Embryology; Wiley Blackwell: Hoboken, NJ, USA, 2017; p. 400. [Google Scholar]
- Moura, E.; Pimpão, C.T. Cleft Lip and Palate in the Dog: Medical and Genetic Aspects; InTech: Toyama, Japan, 2017. [Google Scholar]
- Usui, K.; Tokita, M. Creating diversity in mammalian facial morphology: A review of potential developmental mechanisms. EvoDevo 2018, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Łobodzinska, A.; Gruszczynska, J.; Max, A.; Jan Bartyzel, B.; Mikula, M.; Mikula, J.I.; Grzegrzółka, B. Cleft palate in the domestic dog Canis Lupus Familiaris- etiology, pathophtsiology, diagnosis, prevention and treatment. Acta. Sci. Polonorum. Zootech. 2014, 13, 5–28. [Google Scholar]
- Leslie, E.J.; Liu, H.; Carlson, J.; Shaffer, J.R.; Feingold, E.; Wehby, G.; Laurie, C.A.; Jain, D.; Laurie, C.C.; Doheny, K.F.; et al. A Genome-wide Association Study ofS Nonsyndromic Cleft Palate Identifies an Etiologic Missense Variant in GRHL3. Am. J. Hum. Genet. 2016, 98, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Bush, J.O.; Lidral, A.C. Development of the upper lip: Morphogenetic and molecular mechanisms. Dev. Dyn. 2006, 235, 1152–1166. [Google Scholar] [CrossRef] [PubMed]
- Juriloff, D.M.; Harris, M.J. Mouse genetic models of cleft lip with or without cleft palate. Birth Defects Res. Part A Clin. Mol. Teratol. 2008, 82, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Gebuijs, I.G.E.; Raterman, S.T.; Metz, J.R.; Swanenberg, L.; Zethof, J.; Van den Bos, R.; Carels, C.E.L.; Wagener, F.A.D.T.G.; Von den Hoff, J.W. Fgf8a mutation affects craniofacial development and skeletal gene expression in zebrafish larvae. Biology Open 2019, 8, bio039834. [Google Scholar] [CrossRef]
- Kazempoor, R.; Akbarinejad, V.; Mardjanmehr, S.H.; Shojaei, M.; Soroori, S.; Amini, M. Bilateral cleft lip, jaw, and palate in a female Holstein calf. Comp. Clin. Pathol. 2012, 21, 349–352. [Google Scholar] [CrossRef]
- Reinartz, S.; Hellige, M.; Feige, K.; Wenning, P.; Distl, O. Phenotypic classification of variability of non-syndromic congenital cleft lip and jaw in Vorderwald × Montbéliarde cattle. Acta Vet. Scand. 2015, 57, 87. [Google Scholar] [CrossRef]
- Koul, R. Describing Cleft Lip and Palate Using a New Expression System. Cleft Palate-Craniofacial J. 2007, 44, 585–589. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, M.-L.; Li, Z.-J.; Bai, X.-F.; Wang, X.-K.; Lu, L.; Wang, Y.-X. A Simple and Precise Classification for Cleft Lip and Palate: A Five-Digit Numerical Recording System. Cleft Palate-Craniofacial J. 2007, 44, 465–468. [Google Scholar] [CrossRef]
- Guo, H.L.; Zhao, L.; Liu, Q.; Li, Z.J.; Zhang, B.; Lu, L. Clinical evaluation of the five-digit numerical recording system for classification of cleft lip and palate deformities. Chin. J. Stomatol. 2013, 48, 15–17. [Google Scholar]
- Saleem, K.; Zaib, T.; Sun, W.; Fu, S. Assessment of candidate genes and genetic heterogeneity in human non syndromic orofacial clefts specifically non syndromic cleft lip with or without palate. Heliyon 2019, 5, e03019. [Google Scholar] [CrossRef] [PubMed]
- Lupp, B.; Reinhardt, M.; Maus, F.; Hellige, M.; Feige, K.; Distl, O. Right-sided cleft lip and jaw in a family of Vorderwald×Montbéliarde cattle. Vet. J. 2012, 192, 520–522. [Google Scholar] [CrossRef]
- Shaw, S.D.; Norman, T.E.; Arnold, C.E.; Coleman, M.C. Clinical characteristics of horses and foals diagnosed with cleft palate in a referral population: 28 cases (1988–2011). Can Vet. J. 2015, 56, 756–760. [Google Scholar] [PubMed]
- Oryan, A.; Shirian, S.; Samadian, M.R. Congenital craniofacial and skeletal defects with arthrogryposis in two newborn male Holstein Friesian calves. Comp. Clin. Pathol. 2011, 20, 43–46. [Google Scholar] [CrossRef]
- Moritomo, Y.; Tsuda, T.; Miyamoto, H. Craniofacial Skeletal Abnormalities in Anomalous Calves with Clefts of the Face. J. Vet. Med. Sci. 1999, 61, 1147–1152. [Google Scholar] [CrossRef]
- Stuppia, L.; Capogreco, M.; Marzo, G.; La Rovere, D.; Antonucci, I.; Gatta, V.; Palka, G.; Mortellaro, C.; Tetè, S. Genetics of Syndromic and Nonsyndromic Cleft Lip and Palate. J. Craniofacial Surg. 2011, 22, 1722–1726. [Google Scholar] [CrossRef]
- Zucchero, T.M.; Cooper, M.E.; Maher, B.S.; Daack-Hirsch, S.; Nepomuceno, B.; Ribeiro, L.; Caprau, D.; Christensen, K.; Suzuki, Y.; Machida, J.; et al. Interferon Regulatory Factor 6 (IRF6) Gene Variants and the Risk of Isolated Cleft Lip or Palate. N. Engl. J. Med. 2004, 351, 769–780. [Google Scholar] [CrossRef]
- Dixon, M.J.; Marazita, M.L.; Beaty, T.H.; Murray, J.C. Cleft lip and palate: Understanding genetic and environmental influences. Nat. Rev. Genet. 2011, 12, 167–178. [Google Scholar] [CrossRef]
- Sylvester, B.; Brindopke, F.; Suzuki, A.; Giron, M.; Auslander, A.; Maas, R.L.; Tsai, B.; Gao, H.; Magee, W.; Cox, T.C.; et al. A Synonymous Exonic Splice Silencer Variant in IRF6 as a Novel and Cryptic Cause of Non-Syndromic Cleft Lip and Palate. Genes 2020, 11, 903. [Google Scholar] [CrossRef]
- Robledo, R.F.; Rajan, L.; Li, X.; Lufkin, T. The Dlx5 and Dlx6 homeobox genes are essential for craniofacial, axial, and appendicular skeletal development. Genes Dev. 2002, 16, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Hristov, K.; Stoimenov, A. Hypospadias in small ruminants: A case report. Tradit. Mod. Vet. Med. 2020, 5, 58–61. [Google Scholar] [CrossRef]
- Usta, Z.; Distl, O. Atresia Ani, Hypospadia and Rudimentary External Genitalia in Two German Holstein Calves. Atatürk Üniversitesi Vet. Bil. Derg. 2017, 12, 195–202. [Google Scholar] [CrossRef]
- Radostits, O.; Gay, C.; Hinchcliff, K.; Constable, P. Veterinary Medicine, a Textbook of the Diseases of Cattle, Horses, Sheep, Pig and Goats, 10th ed.; Elsevier Saunders London: London, UK, 2007. [Google Scholar]
- Piñeyro-Ruiz, C.; Serrano, H.; Pérez-Brayfield, M.R.; Jorge, J.C. New frontiers on the molecular underpinnings of hypospadias according to severity. Arab. J. Urol. 2020, 18, 257–266. [Google Scholar] [CrossRef]
- Chang, J.; Wang, S.; Zheng, Z. Etiology of Hypospadias: A Comparative Review of Genetic Factors and Developmental Processes Between Human and Animal Models. Res. Rep. Urol. 2020, 12, 673–686. [Google Scholar] [CrossRef]
- Almubarak, A.M.; Abdelghafar, R.M.; Gameel, A.A.; Osman, N.M. Penile Urethral Hypospadias with Two Fistulae and Diverticulum in a Saanen Kid. Case Rep. Vet. Med. 2016, 2016, 6534062. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Braun, M.; Genualdo, V.; Perucatti, A.; Reinartz, S.; Proios, I.; Heppelmann, M.; Rehage, J.; Hülskötter, K.; Beineke, A.; et al. Clinical, cytogenetic and molecular genetic characterization of a tandem fusion translocation in a male Holstein cattle with congenital hypospadias and a ventricular septal defect. PLoS ONE 2020, 15, e0227117. [Google Scholar] [CrossRef]
- Harrison, J.; Corley, K.; Kearney, C.; Mushtaq, I. Single stage urethroplasty for perineal hypospadias in a foal. Equine Vet. Educ. 2018, 30, 527–530. [Google Scholar] [CrossRef]
- Abd-El-Hady, A.A.A.; Mohi Eldin, M.M. Hypospadia and urethral diverticulum in a female pseudohermaphrodite calf. Sch. J. Agric. Vet. Sci. 2014, 1, 288–292. [Google Scholar]
- Hoque, M. A Case Report on Surgical Management of Hypospadias in a Day-Old Goat Kid. Int. J. Livest. Res. 2021, 11, 175–178. [Google Scholar] [CrossRef]
- Switonski, M.; Dzimira, S.; Aleksiewicz, R.; Szczerbal, I.; Nowacka-Woszuk, J.; Krzeminska, P.; Deska, T.; Nizanski, W. Hypospadias Is Not Rare in Dogs: Five New Cases, a Retrospective Study, and a Review of the Literature. Sex. Dev. 2018, 12, 244–250. [Google Scholar] [CrossRef]
- Meyers-Wallen, V.N. Gonadal and sex differentiation abnormalities of dogs and cats. Sex. Dev. 2012, 6, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T. Anatomical Examination of Hypospadias in Cattle. J. Jpn. Vet. Med. Assoc. 2008, 61, 931–935. [Google Scholar] [CrossRef]
- Kumi-Diaka, J.; Osori, D.I.K. Perineal hypospadias in two related bull calves, a case report. Theriogenology 1979, 11, 163–164. [Google Scholar] [CrossRef] [PubMed]
- van der Zanden, L.F.M.; van Rooij, I.A.L.M.; Feitz, W.; Franke, B.; Knoers, N.V.A.M.; Roeleveld, N. Aetiology of hypospadias: A systematic review of genes and environment. Hum. Reprod. Update 2012, 18, 260–283. [Google Scholar] [CrossRef]
- Raghavan, R.; Romano, M.E.; Karagas, M.R.; Penna, F.J. Pharmacologic and Environmental Endocrine Disruptors in the Pathogenesis of Hypospadias: A Review. Curr. Environ. Health Rep. 2018, 5, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Bouty, A.; Ayers, K.L.; Pask, A.; Heloury, Y.; Sinclair, A.H. The Genetic and Environmental Factors Underlying Hypospadias. Sex. Dev. 2015, 9, 239–259. [Google Scholar] [CrossRef]
- Chen, Z.-Z.; Gao, Y.-Q.; Xie, H.; Huang, Y.-C.; Chen, F.; Lei, Y.-P.; Zhu, Y.-Q. Transcription factors dysregulated in three complex birth defects datasets. Reprod. Dev. Med. 2022, 06, 79–85. [Google Scholar] [CrossRef]
- Murashima, A.; Kishigami, S.; Thomson, A.; Yamada, G. Androgens and mammalian male reproductive tract development. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2015, 1849, 163–170. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, H.; Pask, A.J.; Fujiyama, A.; Suzuki, Y.; Sugano, S.; Shaw, G.; Renfree, M.B. Hormone-responsive genes in the SHH and WNT/β-catenin signaling pathways influence urethral closure and phallus growth. Biol. Reprod. 2018, 99, 806–816. [Google Scholar] [CrossRef]
- Govers, L.C.; Phillips, T.R.; Mattiske, D.M.; Rashoo, N.; Black, J.R.; Sinclair, A.; Baskin, L.S.; Risbridger, G.P.; Pask, A.J. A critical role for estrogen signaling in penis development. FASEB J. 2019, 33, 10383–10392. [Google Scholar] [CrossRef] [PubMed]
- Baskin, L.; Cao, M.; Sinclair, A.; Li, Y.; Overland, M.; Isaacson, D.; Cunha, G.R. Androgen and estrogen receptor expression in the developing human penis and clitoris. Differentiation 2020, 111, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Lin, X.; Wang, Y.; Xie, H.; Chen, F. Dysregulated expression of androgen metabolism genes and genetic analysis in hypospadias. Mol. Genet. Genom. Med. 2020, 8, e1346. [Google Scholar] [CrossRef]
- Yucel, S.; Cavalcanti, A.G.; Desouza, A.; Wang, Z.; Baskin, L.S. The effect of oestrogen and testosterone on the urethral seam of the developing male mouse genital tubercle. BJU Int. 2003, 92, 1016–1021. [Google Scholar] [CrossRef] [PubMed]
- Vottero, A.; Minari, R.; Viani, I.; Tassi, F.; Bonatti, F.; Neri, T.M.; Bertolini, L.; Bernasconi, S.; Ghizzoni, L. Evidence for Epigenetic Abnormalities of the Androgen Receptor Gene in Foreskin from Children with Hypospadias. J. Clin. Endocrinol. Metab. 2011, 96, E1953–E1962. [Google Scholar] [CrossRef]
- Ohsako, S.; Aiba, T.; Miyado, M.; Fukami, M.; Ogata, T.; Hayashi, Y.; Mizuno, K.; Kojima, Y. Expression of Xenobiotic Biomarkers CYP1 Family in Preputial Tissue of Patients with Hypospadias and Phimosis and Its Association with DNA Methylation Level of SRD5A2 Minimal Promoter. Arch. Environ. Contam. Toxicol. 2018, 74, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. Babraham Bioinformatics. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 3 August 2022).
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Null, D.J.; Vanraden, P.M.; Rosen, B.D.; O’Connell, J.R.; Bickhart, D.M. Using the ARS-UCD1.2 reference genome in U.S. evaluations. Interbull Bull. 2019, 55, 30–34. [Google Scholar]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013. [Google Scholar] [CrossRef]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef]
- van der Auwera, G.A.; O’Connor, B.D. Genomics in the Cloud; O’Reilly Media, Inc.: Sebastopol, CA, USA, 2020. [Google Scholar]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 5 August 2022).
- Fernández, N.; Pabon, J.; Ayala, P.; Perez, J.; Ortiz, A.M.; Zarante, I. Description of a novel variant in the MAMLD1 gene in isolated hypospadias. Urol. J. 2018, 27, 105–109. [Google Scholar] [CrossRef]
- Xu, H.; Yan, F.; Hu, R.; Suzuki, A.; Iwaya, C.; Jia, P.; Iwata, J.; Zhao, Z. CleftGeneDB: A resource for annotating genes associated with cleft lip and cleft palate. Sci. Bull. 2021, 66, 2340. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.; Shepherd, L. AnnotationHub: Client to Access AnnotationHub Resources. R Package Version 3.4.0. Available online: https://bioconductor.org/packages/release/bioc/html/AnnotationHub.html (accessed on 10 August 2022).
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Lipscomb, C.E. Medical Subject Headings (MeSH). Bull Med. Libr. Assoc. 2000, 88, 265–266. [Google Scholar] [PubMed]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Yu, G. Using meshes for MeSH term enrichment and semantic analyses. Bioinformatics 2018, 34, 3766–3767. [Google Scholar] [CrossRef]
- Yu, G.; He, Q.-Y. ReactomePA: An R/Bioconductor package for reactome pathway analysis and visualization. Mol. BioSystems 2016, 12, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Pagès, H.; Carlson, M.; Falcon, S.; Li, N. Bioconductor. Available online: https://bioconductor.org/packages/release/bioc/html/AnnotationDbi.html (accessed on 2 September 2022).
- Yan, F.; Dai, Y.; Iwata, J.; Zhao, Z.; Jia, P. An integrative, genomic, transcriptomic and network-assisted study to identify genes associated with human cleft lip with or without cleft palate. BMC Med. Genom. 2020, 13, 39. [Google Scholar] [CrossRef] [PubMed]
- Krzemińska, P.; D’Anza, E.; Ciotola, F.; Paciello, O.; Restucci, B.; Peretti, V.; Albarella, S.; Switonski, M. Polymorphisms of MAMLD1, SRD5A2 and AR Candidate Genes in Seven Dogs (78,XY.; SRY-Positive) Affected by Hypospadias or Cryptorchidism. Sex. Dev. 2019, 13, 92–98. [Google Scholar] [CrossRef]
- Switonski, M.; Payan-Carreira, R.; Bartz, M.; Nowacka-Woszuk, J.; Szczerbal, I.; Colaço, B.; Pires, M.A.; Ochota, M.; Nizanski, W. Hypospadias in a Male (78,XY.; SRY-Positive) Dog and Sex Reversal Female (78,XX.; SRY-Negative) Dogs: Clinical, Histological and Genetic Studies. Sex. Dev. 2012, 6, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Miyado, M.; Nakamura, M.; Miyado, K.; Morohashi, K.-I.; Sano, S.; Nagata, E.; Fukami, M.; Ogata, T. Mamld1 deficiency significantly reduces mRNA expression levels of multiple genes expressed in mouse fetal Leydig cells but permits normal genital and reproductive development. Endocrinology 2012, 153, 6033–6040. [Google Scholar] [CrossRef]
- Peycelon, M.; Mansour-Hendili, L.; Hyon, C.; Collot, N.; Houang, M.; Legendre, M.; Chabaud, M.; Bouvier, M.D.; Audry, G.; Amselem, S.; et al. Recurrent Intragenic Duplication within the NR5A1 Gene and Severe Proximal Hypospadias. Sex. Dev. 2017, 11, 293–297. [Google Scholar] [CrossRef]
- Song, Y.; Chen, W.; Zhu, B.; Ge, W. Disruption of Epidermal Growth Factor Receptor but Not EGF Blocks Follicle Activation in Zebrafish Ovary. Front. Cell Dev. Biol. 2022, 9, 750888. [Google Scholar] [CrossRef]
- Miettinen, P.J.; Chin, J.R.; Shum, L.; Slavkin, H.C.; Shuler, C.F.; Derynck, R.; Werb, Z. Epidermal growth factor receptor function is necessary for normal craniofacial development and palate closure. Nat. Genet. 1999, 22, 69–73. [Google Scholar] [CrossRef]
- Paiva, K.B.S.; Maas, C.S.; Santos, P.M.d.; Granjeiro, J.M.; Letra, A. Extracellular Matrix Composition and Remodeling: Current Perspectives on Secondary Palate Formation, Cleft Lip/Palate, and Palatal Reconstruction. Front. Cell Dev. Biol. 2019, 7, 340. [Google Scholar] [CrossRef]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016, baw100. [Google Scholar] [CrossRef]
- Noda, K.; Mishina, Y.; Komatsu, Y. Constitutively active mutation of ACVR1 in oral epithelium causes submucous cleft palate in mice. Dev. Biol. 2016, 415, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Duffhues, G.; Williams, E.; Goumans, M.-J.; Heldin, C.-H.; ten Dijke, P. Bone morphogenetic protein receptors: Structure, function and targeting by selective small molecule kinase inhibitors. Bone 2020, 138, 115472. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Y.; Lin, M.; Yuan, G.; Yang, G.; Zheng, Y.; Chen, Y. Augmented BMPRIA-Mediated BMP Signaling in Cranial Neural Crest Lineage Leads to Cleft Palate Formation and Delayed Tooth Differentiation. PLoS ONE 2013, 8, e66107. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.; Siegel, S.; Ellis, D. The role of EGF in testosterone-induced reproductive tract differentiation. Dev. Biol. 1991, 146, 106–116. [Google Scholar] [CrossRef]
- Chen, T.; Li, Q.; Xu, J.; Ding, K.; Wang, Y.; Wang, W.; Li, S.; Shen, Y. Mutation screening of BMP4, BMP7, HOXA4 and HOXB6 genes in Chinese patients with hypospadias. Eur. J. Hum. Genet. 2007, 15, 23–28. [Google Scholar] [CrossRef]
- de la Garza, G.; Schleiffarth, J.R.; Dunnwald, M.; Mankad, A.; Weirather, J.L.; Bonde, G.; Butcher, S.; Mansour, T.A.; Kousa, Y.A.; Fukazawa, C.F.; et al. Interferon Regulatory Factor 6 Promotes Differentiation of the Periderm by Activating Expression of Grainyhead-Like 3. J. Investig. Dermatol. 2013, 133, 68–77. [Google Scholar] [CrossRef]
- Girousi, E.; Muerner, L.; Parisi, L.; Rihs, S.; von Gunten, S.; Katsaros, C.; Degen, M. Lack of IRF6 Disrupts Human Epithelial Homeostasis by Altering Colony Morphology, Migration Pattern, and Differentiation Potential of Keratinocytes. Front. Cell Dev. Biol. 2021, 9, 718066. [Google Scholar] [CrossRef]
- Knight, A.S.; Schutte, B.C.; Jiang, R.; Dixon, M.J. Developmental expression analysis of the mouse and chick orthologues of IRF6: The gene mutated in Van der Woude syndrome. Dev. Dyn. 2006, 235, 1441–1447. [Google Scholar] [CrossRef]
- Jugessur, A.; Rahimov, F.; Lie, R.T.; Wilcox, A.J.; Gjessing, H.K.; Nilsen, R.M.; Nguyen, T.T.; Murray, J.C. Genetic variants in IRF6 and the risk of facial clefts: Single-marker and haplotype-based analyses in a population-based case-control study of facial clefts in Norway. Genet. Epidemiol. 2008, 32, 413–424. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, J.; Zhao, H.; Ievlev, V.; Zhong, W.; Huang, W.; Cornell, R.A.; Lin, J.; Chen, F. Functional Characterization of a Novel IRF6 Frameshift Mutation From a Van Der Woude Syndrome Family. Front. Genet. 2020, 11, 562. [Google Scholar] [CrossRef]
- Gurramkonda, V.B.; Syed, A.H.; Murthy, J.; Lakkakula, B.V.K.S. IRF6 rs2235375 single nucleotide polymorphism is associated with isolated non-syndromic cleft palate but not with cleft lip with or without palate in South Indian population. Braz. J. Otorhinolaryngol. 2018, 84, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Kousa, Y.A.; Moussa, D.; Schutte, B.C. IRF6 expression in basal epithelium partially rescues Irf6 knockout mice. Dev. Dyn. 2017, 246, 670–681. [Google Scholar] [CrossRef]
- Isono, K.-I.; Fujimura, Y.-I.; Shinga, J.; Yamaki, M.; O-Wang, J.; Takihara, Y.; Murahashi, Y.; Takada, Y.; Mizutani-Koseki, Y.; Koseki, H. Mammalian polyhomeotic homologues Phc2 and Phc1 act in synergy to mediate polycomb repression of Hox genes. Mol. Cell Biol. 2005, 25, 6694–6706. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.T.; Wang, Y.; Connor, B.; Daiger, S.P.; Blanton, S.H. Nonsyndromic cleft lip and palate: No evidence of linkage to HLA or factor 13A. Am. J. Hum. Genet. 1993, 52, 1230–1233. [Google Scholar] [PubMed]
- Krapels, I.P.C.; Raijmakers-Eichhorn, J.; Peters, W.H.M.; Roelofs, H.M.J.; Ras, F.; Steegers-Theunissen, R.P.M.; the Eurocran Gene–Environment Interaction, G. The I105V polymorphism in glutathione S-transferase P1, parental smoking and the risk for nonsyndromic cleft lip with or without cleft palate. Eur. J. Hum. Genet. 2008, 16, 358–366. [Google Scholar] [CrossRef]
- Wang, P.; Yan, F.; Li, Z.; Yu, Y.; Parnell, S.E.; Xiong, Y. Impaired plasma membrane localization of ubiquitin ligase complex underlies 3-M syndrome development. J. Clin. Investig. 2019, 129, 4393–4407. [Google Scholar] [CrossRef]
- Hanson, D.; Murray, P.G.; O’Sullivan, J.; Urquhart, J.; Daly, S.; Bhaskar, S.S.; Biesecker, L.G.; Skae, M.; Smith, C.; Cole, T.; et al. Exome sequencing identifies CCDC8 mutations in 3-M syndrome, suggesting that CCDC8 contributes in a pathway with CUL7 and OBSL1 to control human growth. Am. J. Hum Genet 2011, 89, 148–153. [Google Scholar] [CrossRef]
- Selvanathan, A.; Nixon, C.Y.; Zhu, Y.; Scietti, L.; Forneris, F.; Moreno Uribe, L.M.; Lidral, A.C.; Jezewski, P.A.; Mulliken, J.B.; Murray, J.C.; et al. CDH1 Mutation Distribution and Type Suggests Genetic Differences between the Etiology of Orofacial Clefting and Gastric Cancer. Genes 2020, 11, 391. [Google Scholar] [CrossRef]
- Frebourg, T.; Oliveira, C.; Hochain, P.; Karam, R.; Manouvrier, S.; Graziadio, C.; Vekemans, M.; Hartmann, A.; Baert-Desurmont, S.; Alexandre, C.; et al. Cleft lip/palate and CDH1/E-cadherin mutations in families with hereditary diffuse gastric cancer. J. Med. Genet 2006, 43, 138–142. [Google Scholar] [CrossRef]
- Katsonis, P.; Wilhelm, K.; Williams, A.; Lichtarge, O. Genome interpretation using in silico predictors of variant impact. Hum. Genet. 2022, 141, 1549–1577. [Google Scholar] [CrossRef]
- Richard, M.A.; Sok, P.; Canon, S.; Nembhard, W.N.; Brown, A.L.; Peckham-Gregory, E.C.; Ton, M.; Ehli, E.A.; Kallsen, N.A.; Peyton, S.A.; et al. Altered mechanisms of genital development identified through integration of DNA methylation and genomic measures in hypospadias. Sci. Rep. 2020, 10, 12715. [Google Scholar] [CrossRef] [PubMed]
- Kaefer, M.; Rink, R.; Misseri, R.; Winchester, P.; Proctor, C.; Ben Maamar, M.; Beck, D.; Nilsson, E.; Skinner, M.K. Role of epigenetics in the etiology of hypospadias through penile foreskin DNA methylation alterations. Sci. Rep. 2023, 13, 555. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Type of Variant | Variant a | Existing Variant ID | Consequence of the Variant | Zygosity b |
|---|---|---|---|---|---|
| ACVR1 | deletion | 2:g38466740delC | NA | frameshift | HET |
| ADGRA2 | substitution | 27:g.33105944C>T | rs109168787 | stop gained | HOM |
| BHMT2 | substitution | 10:g.10153010G>A | rs384114512 | splice acceptor | HET |
| BMPR1B | substitution | 6:g.29483476G>A | rs133060728 | frameshift | HET |
| CCDC8 | insertion | 18:g.53693827_53693828 ins CAGACAA | rs1118258295 | frameshift | HET |
| CCDC8 | insertion | 18:g53693828-53693829 ins CAGAGGGCAGAGGCCCC | NA | frameshift | HET |
| CDH1 | insertion | 18:36101086-36101087insT | NA | frameshift, splice region, intron variant | HET |
| EGF | substitution | 6:g.15450370G>A | rs136048352 | Stop gained | HOM |
| F13A1 | deletion | 23:48853544delA | NA | frameshift, splice donor region variant | HOM |
| GSTP1 | deletion | 29:45444664delG | NA | frameshift | HOM |
| IRF6 | insertion | 16:g73512380-73512381insT | NA | frameshift | HOM |
| MMP14 | insertion | 10:g21978036-21978037insA | NA | frameshift | HET |
| MYBPHL | deletion | 3:g.34117074delC | rs480413110 | frameshift | HET |
| PHC2 | substitution | 2:120387448C>T | NA | Stop gained | HET |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marc, S.; Mizeranschi, A.E.; Paul, C.; Otavă, G.; Savici, J.; Sicoe, B.; Torda, I.; Huțu, I.; Mircu, C.; Ilie, D.E.; et al. Simultaneous Occurrence of Hypospadias and Bilateral Cleft Lip and Jaw in a Crossbred Calf: Clinical, Computer Tomographic, and Genomic Characterization. Animals 2023, 13, 1709. https://doi.org/10.3390/ani13101709
Marc S, Mizeranschi AE, Paul C, Otavă G, Savici J, Sicoe B, Torda I, Huțu I, Mircu C, Ilie DE, et al. Simultaneous Occurrence of Hypospadias and Bilateral Cleft Lip and Jaw in a Crossbred Calf: Clinical, Computer Tomographic, and Genomic Characterization. Animals. 2023; 13(10):1709. https://doi.org/10.3390/ani13101709
Chicago/Turabian StyleMarc, Simona, Alexandru Eugeniu Mizeranschi, Cristina Paul, Gabriel Otavă, Jelena Savici, Bogdan Sicoe, Iuliu Torda, Ioan Huțu, Călin Mircu, Daniela Elena Ilie, and et al. 2023. "Simultaneous Occurrence of Hypospadias and Bilateral Cleft Lip and Jaw in a Crossbred Calf: Clinical, Computer Tomographic, and Genomic Characterization" Animals 13, no. 10: 1709. https://doi.org/10.3390/ani13101709