
Molecular Epidemiology of Pasteurella multocida Associated with Bovine Respiratory Disease Outbreaks
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pasteurella Multocida Isolates
2.2. DNA Samples
2.3. Capsular and Lipopolysaccharide Typing
2.4. Virulence-Associated Gene Typing
2.5. Multilocus Sequence Typing (MLST)
2.6. Pulsed-Field Gel Electrophoresis (PFGE)
2.7. Data Analysis
3. Results
3.1. Capsular and LPS Genotypes
3.2. Virulotypes
3.3. MLST
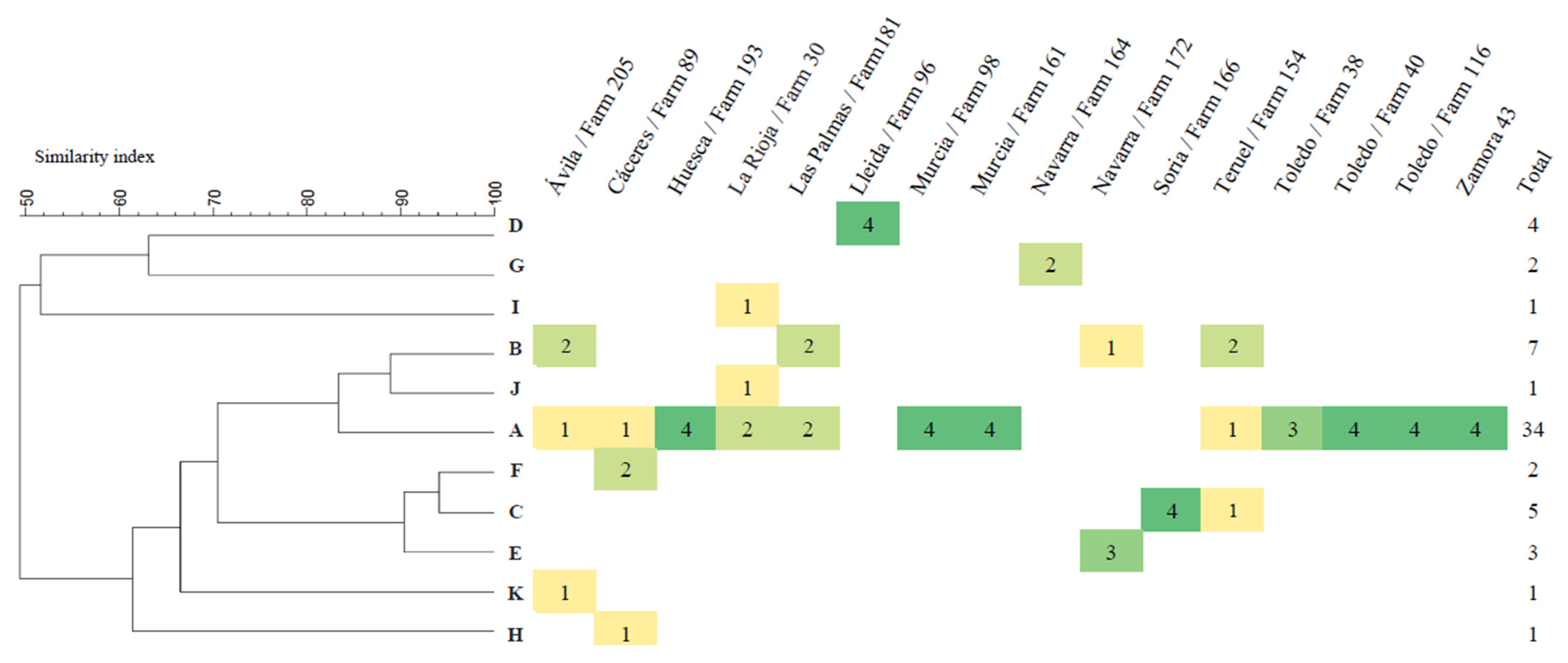
3.4. PFGE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, B.J.; Larson, B.L. Impact of bovine respiratory disease in U.S. beef cattle. Anim. Health Res. Rev. 2020, 21, 132–134. [Google Scholar] [CrossRef] [PubMed]
- Pardon, B.; Callens, J.; Maris, J.; Allais, L.; Van Praet, W.; Deprez, P.; Ribbens, S. Pathogen-specific risk factors in acute outbreaks of respiratory disease in calves. J. Dairy Sci. 2020, 103, 2556–2566. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, A.; Hodnik, A.; Cirone, F.; Capozza, P.; Trotta, A.; Corrente, M.; Balestrieri, A.; Buonavoglia, C. Bovine respiratory disease in beef calves supported long transport stress: An epidemiological study and strategies for control and prevention. Res. Vet. Sci 2020, 135, 450–455. [Google Scholar] [CrossRef]
- Smith, R.A.; Step, D.L.; Woolums, A.R. Bovine respiratory disease looking back and looking forward, what do we see? Vet. Clin. Food Anim. 2020, 36, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Fulton, R.W. Viruses in bovine respiratory disease in North America knowledge advances using genomic testing. Vet. Clin. Food Anim. 2020, 36, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Pardon, B.; Buczinski, S. Bovine respiratory disease diagnosis what progress has been made in infectious diagnosis? Vet. Clin. Food Anim. 2020, 36, 425–444. [Google Scholar] [CrossRef]
- Snyder, E.; Credille, B. Mannheimia haemolytica and Pasteurella multocida in bovine respiratory disease. How are they changing in response to efforts to control them? Vet. Clin. Food Anim. 2020, 36, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Timsit, E.; Hallewell, J.; Booker, C.; Tison, N.; Amat, S.; Alexander, T.W. Prevalence and antimicrobial susceptibility of Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni isolated from the lower respiratory tract of healthy feedlot cattle and those diagnosed with bovine respiratory disease. Vet. Microbiol. 2017, 208, 118–125. [Google Scholar] [CrossRef]
- Yaman, T.; Büyükbayram, H.; Özyıldız, Z.; Terzi, F.; Uyar, A.; Keles, Ö.F.; Özsoy, Ş.Y.; Yener, Z. Detection of bovine respiratory syncytial virus, Pasteurella multocida, and Mannheimia haemolytica by immunohistochemical method in naturally-infected Cattle. J. Vet. Res. 2018, 62, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, M.; Choudhary, B.K.; Chandra Ghosh, R.; Bhoyar, S.; Chaudhari, S.; Barbuddhe, S.B. Cultivable microbiota and pulmonary lesions in polymicrobial bovine pneumonia. Microb. Pathog. 2019, 134, 103577. [Google Scholar] [CrossRef]
- Crawshaw, W.M.; Caldow, G.L. Field study of pneumonia in vaccinated cattle associated with incorrect vaccination and Pasteurella multocida infection. Vet. Rec. 2015, 176, 17. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.D.; Harper, M.; Wilkie, I.W.; Adler, B. Pasteurella. In Pathogenesis of Bacterial Infections in Animals, 4th ed.; Gyles, C.L., Prescott, J.F., Songer, J.G., Thoen, C.O., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 325–346. [Google Scholar]
- Harper, M.; John, M.; Turni, C.; Edmunds, M.; Michael, F.S.; Adler, B.; Blackall, P.J.; Cox, A.D.; Boyce, J.D. Development of a rapid multiplex PCR assay to genotype Pasteurella multocida strains by use of the lipopolysaccharide outer core biosynthesis locus. J. Clin. Microbiol. 2015, 53, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Liu, J.; Liang, W.; Wang, F.; Wang, L.; Wang, X.; Hua, L.; Chen, H.; Wilson, B.A.; Wang, J.; et al. Development of an online tool for Pasteurella multocida genotyping and genotypes of Pasteurella multocida from different hosts. Front. Vet. Sci. 2021, 8, 771157. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.M.; Boyce, J.D.; Chung, J.Y.; Frost, A.J.; Adler, B. Genetic organization of Pasteurella multocida cap loci and development of a multiplex capsular PCR typing system. J. Clin. Microbiol. 2001, 39, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.; Miller, E.; Aguayo, J.M.; Figueroa, C.F.; Nezworski, J.; Studniski, M.; Wileman, B.; Johnson, T. Genomic diversity and molecular epidemiology of Pasteurella multocida. PLoS ONE 2021, 16, e0249138. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, X.; Zhou, R.; Chen, H.; Wilson, B.A.; Wu, B. Pasteurella multocida: Genotypes and genomics. Microbiol. Mol. Biol. Rev. 2019, 83, 00014–00019. [Google Scholar] [CrossRef]
- Ewers, C.; Lübke-Becker, A.; Bethe, A.; Kießling, S.; Filter, M.; Wieler, L.H. Virulence genotype of Pasteurella multocida strains isolated from different hosts with various disease status. Vet. Microbiol. 2006, 114, 304–317. [Google Scholar] [CrossRef]
- Hotchkiss, E.J.; Hodgson, J.C.; Lainson, F.A.; Zadoks, R.N. Multilocus sequence typing of a global collection of Pasteurella multocida isolates from cattle and other host species demonstrates niche association. BMC Microbiol. 2011, 11, 115. [Google Scholar] [CrossRef] [Green Version]
- Hotchkiss, E.J.; Hodgson, J.C.; Schmitt-van de Leemput, E.; Dagleish, M.P.; Zadoks, R.N. Molecular epidemiology of Pasteurella multocida in dairy and beef calves. Vet. Microbiol. 2011, 151, 329–335. [Google Scholar] [CrossRef]
- García-Alvarez, A.; Chaves, F.; Fernández, A.; Sanz, C.; Borobia, M.; Cid, D. An ST11 clone of Pasteurella multocida, widely spread among farmed rabbits in the Iberian Peninsula, demonstrates respiratory niche association. Infect. Genet. Evol. 2015, 34, 81–87. [Google Scholar] [CrossRef]
- García-Alvarez, A.; Vela, A.I.; San Martín, E.; Chaves, F.; Fernández-Garayzábal, J.F.; Domínguez, L.; Cid, D. Characterization of Pasteurella multocida associated with ovine pneumonia using multi-locus sequence typing (MLST) and virulence associated gene profile analysis and comparison with porcine isolates. Vet. Microbiol. 2017, 204, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Massacci, F.R.; Magistrali, C.F.; Cucco, L.; Curcio, L.; Bano, L.; Mangili, P.; Scoccia, E.; Bisgaard, M.; Aalbæk, B.; Christensen, H. Characterization of Pasteurella multocida involved in rabbit infections. Vet. Microbiol. 2018, 213, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Turni, C.; Singh, R.; Blackall, P. Genotypic diversity of Pasteurella multocida isolates from pigs and poultry in Australia. Austral. Vet. J. 2018, 96, 390–394. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, H.; Liang, W.; Chen, Y.; Tang, X.; Chen, H.; Wu, B. A capsule/lipopolysaccharide/MLST genotype D/L6/ST11 of Pasteurella multocida is likely to be strongly associated with swine respiratory disease in China. Arch. Microbiol. 2018, 200, 107–118. [Google Scholar] [CrossRef]
- Peng, Z.; Liang, W.; Wang, F.; Xu, Z.; Xie, Z.; Lian, Z.; Hua, L.; Zhou, R.; Chen, H.; Wu, B. Genetic and phylogenetic characteristics of Pasteurella multocida isolates from different host species. Front. Microbiol. 2018, 9, 1408. [Google Scholar] [CrossRef] [PubMed]
- Karthik, K.; Devi, R.P.; Chitra, M.A.; Govindan, K.; Bharathi, R.; Sridhar, R. Virulence genes and enterobacterial repetitive intergenic consensus region (ERIC) profiling reveals highly diverse genetic population among avian strains of Pasteurella multocida. Microb. Pathog. 2021, 161, 105303. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Sharma, M.; Katoch, S.; Verma, L.; Kumar, S.; Dogra, V.; Chahota, R.; Dhar, P.; Singh, G. Profiling of virulence associated genes of Pasteurella multocida isolated from cattle. Vet. Res. Commun. 2013, 37, 83–89. [Google Scholar] [CrossRef]
- Jamali, H.; Rezagholipour, M.; Fallah, S.; Dadrasnia, A.; Chelliah, S.; Velappan, R.D.; Wei, K.S.C.; Ismail, S. Prevalence, characterization and antibiotic resistance of Pasteurella multocida isolated from bovine respiratory infection. Vet. J. 2014, 202, 381–383. [Google Scholar] [CrossRef]
- Aski, H.S.; Tabatabaei, M. Occurrence of virulence-associated genes in Pasteurella multocida isolates obtained from different hosts. Microb Pathog. 2016, 96, 52e57. [Google Scholar] [CrossRef]
- Katsuda, K.; Hoshinoo, K.; Ueno, Y.; Kohmoto, M.; Mikami, O. Virulence genes and antimicrobial susceptibility in Pasteurella multocida isolates from calves. Vet. Microbiol. 2013, 167, 737–741. [Google Scholar] [CrossRef]
- Gülaydin, Ö.; Gürtürk, K. Investigation of biotype, genotype and virulence associated genes in Pasteurella multocida capsular type A strains from the respiratory tract of cattle and their relationship with disease cases. Turkish. J. Vet. Anim. Sci. 2020, 44, 503–510. [Google Scholar] [CrossRef]
- Townsend, K.M.; Frost, A.J.; Lee, C.W.; Papadimitriou, J.M.; Dawkins, H.J.S. Development of PCR assays for species-and type-specific identification of Pasteurella multocida isolates. J. Clin. Microbiol. 1998, 36, 1096–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Zhao, Z.; Hu, J.; Wu, B.; Cai, X.; He, Q.; Chen, H. Isolation, antimicrobial resistance, and virulence genes of Pasteurella multocida strains from swine in China. J. Clin. Microbiol. 2009, 47, 951–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doughty, S.W.; Ruffolo, C.G.; Adler, B. The type 4 fimbrial subunit gene of Pasteurella multocida. Vet. Microbiol. 2000, 72, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Subaaharan, S.; Blackall, L.L.; Blackall, P.J. Development of a multi-locus sequence typing scheme for avian isolates of Pasteurella multocida. Vet. Microbiol. 2010, 141, 354–361. [Google Scholar] [CrossRef]
- Gharib Mombeni, E.; Gharibi, D.; Ghorbanpoor, M.; Jabbari, A.R.; Cid, D. Toxigenic and non-toxigenic Pasteurella multocida genotypes, based on capsular, LPS, and virulence profile typing, associated with pneumonic pasteurellosis in Iran. Vet. Microbiol. 2021, 257, 109077. [Google Scholar] [CrossRef]
- García-Alvarez, A.; Fernández-Garayzábal, J.F.; Chaves, F.; Pinto, C.; Cid, D. Ovine Mannheimia haemolytica isolates from lungs with and without pneumonic lesions belong to similar genotypes. Vet. Microbiol. 2018, 219, 80–86. [Google Scholar] [CrossRef]
- Griffin, D.; Chengappa, M.M.; Kuszak, J.; McVey, D.S. Bacterial pathogens of the bovine respiratory disease complex. Vet. Clin. Food Anim. 2010, 26, 381–394. [Google Scholar] [CrossRef]
- Turni, C.; Dayao, D.; Aduriz, G.; Cortabarria, N.; Tejero, C.; Ibabe, J.C.; Singh, R.; Blackall, P. A Pasteurella multocida strain affecting nulliparous heifers and calves in different ways. Vet. Microbiol. 2016, 195, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Sánchez del Rey, V.; Fernández-Garayzábal, J.F.; Domínguez, L.; Gottschalk, M.; Vela, A.I. Screening of virulence-associated genes as a molecular typing method for characterization of Streptococcus suis isolates recovered from wild boars and pigs. Vet. J. 2016, 209, 108-12. [Google Scholar] [CrossRef]
- Khamesipour, F.; Momtaz, H.; Mamoreh, M.A. Occurrence of virulence factors and antimicrobial resistance in Pasteurella multocida strains isolated from slaughter cattle in Iran. Front. Microbiol. 2014, 5, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Canovas, L.; Martinez Benitez, M.B.; Herrera Isidron, J.A.; Flores Soto, E. Pulsed Field Gel Electrophoresis: Past, present, and future. Anal. Biochem. 2019, 573, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Fulton, R.W.; Dabo, S.M.; Lehenbauer, T.W.; Confer, A.W. Comparison of genotypic and phenotypic characterization methods for Pasteurella multocida isolates from fatal cases of bovine respiratory disease J. Vet. Diagn. Invest. 2010, 22, 366–375. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene | Primer | Sequence (5′-3′) | Size (bp) | Reference |
|---|---|---|---|---|
| Identification and Typing | ||||
| KMT | kmt1T7 | ATCCGCTATTTACCCAGTGG | 460 | [33] |
| kmt1SP6 | GCTGTAAACGAACTCGCCAC | |||
| Capsular type A | hyaD/hyaC-Fw | TGCCAAAATCGCAGTCAG | 1044 | [15] |
| hyaD/hyaC-Rv | TTGCCATCATTGTCAGTG | |||
| Capsular type B | bcbD-Fw | CATTTATCCAAGCTCCACC | 758 | [15] |
| bcbD-Rv | GCCCGAGAGTTTCAATCC | |||
| Capsular type D | dcbF-Fw | TTACAAAAGAAAGACTAGGAGCCC | 647 | [15] |
| dcbF-Rv | CATCTACCCACTCAACCATATCAG | |||
| Capsular type E | ecbJ-Fw | TCCGCAGAAAATTATTGACTC | 512 | [15] |
| ecbJ-Rv | GCTTGCTGCTTGATTTTGTC | |||
| Capsular type F | fcbD-Fw | AATCGGAGAACGCAGAAATCAG | 852 | [15] |
| fcbD-Rv | TTCCGCCGTCAATTACTCTG | |||
| LPS genotype 1 | BAP6119 | ACATTCCAGATAATACACCCG | 1307 | [13] |
| BAP6120 | ATTGGAGCACCTAGTAACCC | |||
| LPS genotype 2 | BAP6121 | CTTAAAGTAACACTCGCTATTGC | 810 | [13] |
| BAP6122 | TTTGATTTCCCTTGGGATAGC | |||
| LPS genotype 3 | BAP7213 | TGCAGGCGAGAGTTGATAAACCATC | 474 | [13] |
| BAP7214 | CAAAGATTGGTTCCAAATCTGAATGGA | |||
| LPS genotype 4 | BAP6125 | TTTCCATAGATTAGCAATGCCG | 550 | [13] |
| BAP6126 | CTTTATTTGGTCTTTATATATACC | |||
| LPS genotype 5 | BAP6129 | AGATTGCATGGCGAAATGGC | 1175 | [13] |
| BAP6130 | CAATCCTCGTAAGACCCCC | |||
| LPS genotype 6 | BAP7292 | TCTTTATAATTATACTCTCCCAAGG | 668 | [13] |
| BAP7293 | AATGAAGGTTTAAAAGAGATAGCTGGAG | |||
| LPS genotype 7 | BAP6127 | CCTATATTTATATCTCCTCCCC | 931 | [13] |
| BAP6128 | CTAATATATAAACCATCCAACGC | |||
| LPS genotype 8 | BAP6133 | GAGAGTTACAAAAATGATCGGC | 255 | [13] |
| BAP6134 | TCCTGGTTCATATATAGGTAGG | |||
| Virulence Associated Genes (VAGs) | ||||
| Superoxide dismutase A | sodA-Fw | TACCAGAATTAGGCTACGC | 361 | [18] |
| sodA-Rv | GAAACGGGTTGCTGCCGCT | |||
| Outer membrane protein H | ompH-Fw | CGCGTATGAAGGTTTAGGT | 438 | [18] |
| ompH-Rv | TTTAGATTGTGCGTAGTCAAC | |||
| Transferrin binding protein A | tbpA-Fw | TTGGTTGGAAACGGTAAAGC | 728 | [18] |
| tbpA-Rv | TAACGTGTACGGAAAAGCCC | |||
| Putative tight adherence protein D | tadD-Fw | TCTACCCATTCTCAGCAAGGC | 416 | [34] |
| tadD-Rv | ATCATTTCGGGCATTCACC | |||
| Autotransporter adhesion | hsf1-Fw | TTGAGTCGGCTGTAGAGTTCG | 654 | [34] |
| hsf1-Rv | ACTCTTTAGCAGTGGGGACAACCTC | |||
| B hemoglobin-binding protein | hgbB-Fw | ACCGCGTTGGAATTATGATTG | 788 | [18] |
| hgbB-Rv | CATTGAGTACGGCTTGACAT | |||
| Filamentous hemagglutinin | pfhA-Fw | AGCTGATCAAGTGGTGAAC | 275 | [18] |
| pfhA-Rv | TGGTACATTGGTGAATGCTG | |||
| Type 4 fimbriae | ptfA-Fw | TGTGGAATTCAGCATTTTAGTGTGTC | 488 | [35] |
| ptfA-Rv | TCATGAATTCTTATGCGCAAAATCCTGCTGG | |||
| Dermonecrotic toxin | toxA-Fw | CTTAGATGAGCGACAAGGTT | 865 | [18] |
| toxA-Rv | GGAATGCCACACCTCTATA | |||
| Multi-Locus Sequence Typing – RIRDC scheme | ||||
| Adenylate kinase | adk-Fw | TTTTTCGTCCCGTCTAAGC | 570 | [36] |
| adk-Rv | GGGGAAAGGGACACAAGC | |||
| Esterase | est-Fw | TCTGGCAAAAGATGTTGTCG | 641 | [36] |
| est-Rv | CCAAATTCTTGGTTGGTTGG | |||
| Mannose-6-phosphate isomerase | pmi-Fw | TGCCTTGAGACAGGGTAAGC | 739 | [36] |
| pmi-Rv | GCCTTAACAAGTCCCATTCG | |||
| Glucose-6-phosphate dehydrogenase | zwf1-Fw | AATCGGTCGTTTGACTGAGC | 808 | [36] |
| zwf1-Rv | TGCTTCACCTTCAACTGTGC | |||
| Malate dehydrogenase | mdh-Fw | ATTTCGGGATCAGGGTTAGC | 620 | [36] |
| mdh-Rv | GGAAAACCGGTAATGGAAGG | |||
| Glutamate dehydrogenase | gdh-Fw | ATCGACTTCTTCCGCAGACC | 702 | [36] |
| gdh-Rv | GCGGGTGATATTGGTGTAGG | |||
| Phosphor glucose isomerase | pgi-Fw | ACCACGCTATTTTTGGTTGC | 784 | [36] |
| pgi-Rv | ATGGCACAACCTCTTTCACC |
| Virulence Profile (VP) | Virulence Associated Genes 1 | No. (%) of Isolates of Capsular Type and LipopolysacCharide (LPS) Genotype | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pfhA | ptfaA | tadD | tbpA | ompH | hfs1 | hgbB | A:L3 | A:L6 | B:L2 | F:L3 | Total | |
| n = 166 | n = 2 | n = 1 | n = 1 | n = 170 | ||||||||
| VP1 | + | + | + | + | - | - | - | 141 (84.9) | 141 (82.9) | |||
| VP2 | + | + | + | + | + | - | - | 22 (13.3) | 22 (12.9) | |||
| VP3 | - | + | + | + | - | - | - | 2 (100) | 2 (1.2) | |||
| VP4 | + | - | + | + | - | - | - | 2 (1.2) | 2 (1.2) | |||
| VP5 | + | + | - | - | - | - | - | 1 (100) | 1 (0.6) | |||
| VP6 | + | + | - | - | + | + | + | 1 (100) | 1 (0.6) | |||
| VP7 | + | + | + | - | + | - | - | 1 (0.6) | 1 (0.6) | |||
| Capsular and LPS Genotype | Virulence Profile | Isolates Analyzed 1 | No. (%) of Isolates of Sequence Type (ST) | ||||
|---|---|---|---|---|---|---|---|
| ST79 | ST13 | ST206 | ST9 | ST322 | |||
| A:L3 | VP1 | 30 | 17 (56.7) | 13 (43.3) | |||
| VP2 | 12 | 12 (100) | |||||
| VP4 | 1 | 1 (100) | |||||
| VP7 | 1 | 1 (100) | |||||
| A:L6 | VP3 | 2 | 2 (100) | ||||
| F:L3 | VP6 | 1 | 1 (100) | ||||
| B:L2 | VP5 | 1 | 1 (100) | ||||
| Total | 48 | 30 (62.5) | 14 (29.2) | 2 (4.2) | 1 (2.1) | 1 (2.1) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calderón Bernal, J.M.; Fernández, A.; Arnal, J.L.; Sanz Tejero, C.; Fernández-Garayzábal, J.F.; Vela, A.I.; Cid, D. Molecular Epidemiology of Pasteurella multocida Associated with Bovine Respiratory Disease Outbreaks. Animals 2023, 13, 75. https://doi.org/10.3390/ani13010075
Calderón Bernal JM, Fernández A, Arnal JL, Sanz Tejero C, Fernández-Garayzábal JF, Vela AI, Cid D. Molecular Epidemiology of Pasteurella multocida Associated with Bovine Respiratory Disease Outbreaks. Animals. 2023; 13(1):75. https://doi.org/10.3390/ani13010075
Chicago/Turabian StyleCalderón Bernal, Johan Manuel, Ana Fernández, José Luis Arnal, Celia Sanz Tejero, José Francisco Fernández-Garayzábal, Ana I. Vela, and Dolores Cid. 2023. "Molecular Epidemiology of Pasteurella multocida Associated with Bovine Respiratory Disease Outbreaks" Animals 13, no. 1: 75. https://doi.org/10.3390/ani13010075