
An Epidemiological Investigation and Drug-Resistant Strain Isolation of Nematodirus oiratianus in Sheep in Inner Mongolia, China
 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Epidemiological Investigation of Nematodiriasis in Sheep in Major Pastures in Inner Mongolia, China
2.3. Isolation and Identification of Naturally Occurring Nematodirus
2.3.1. Morphological Identification
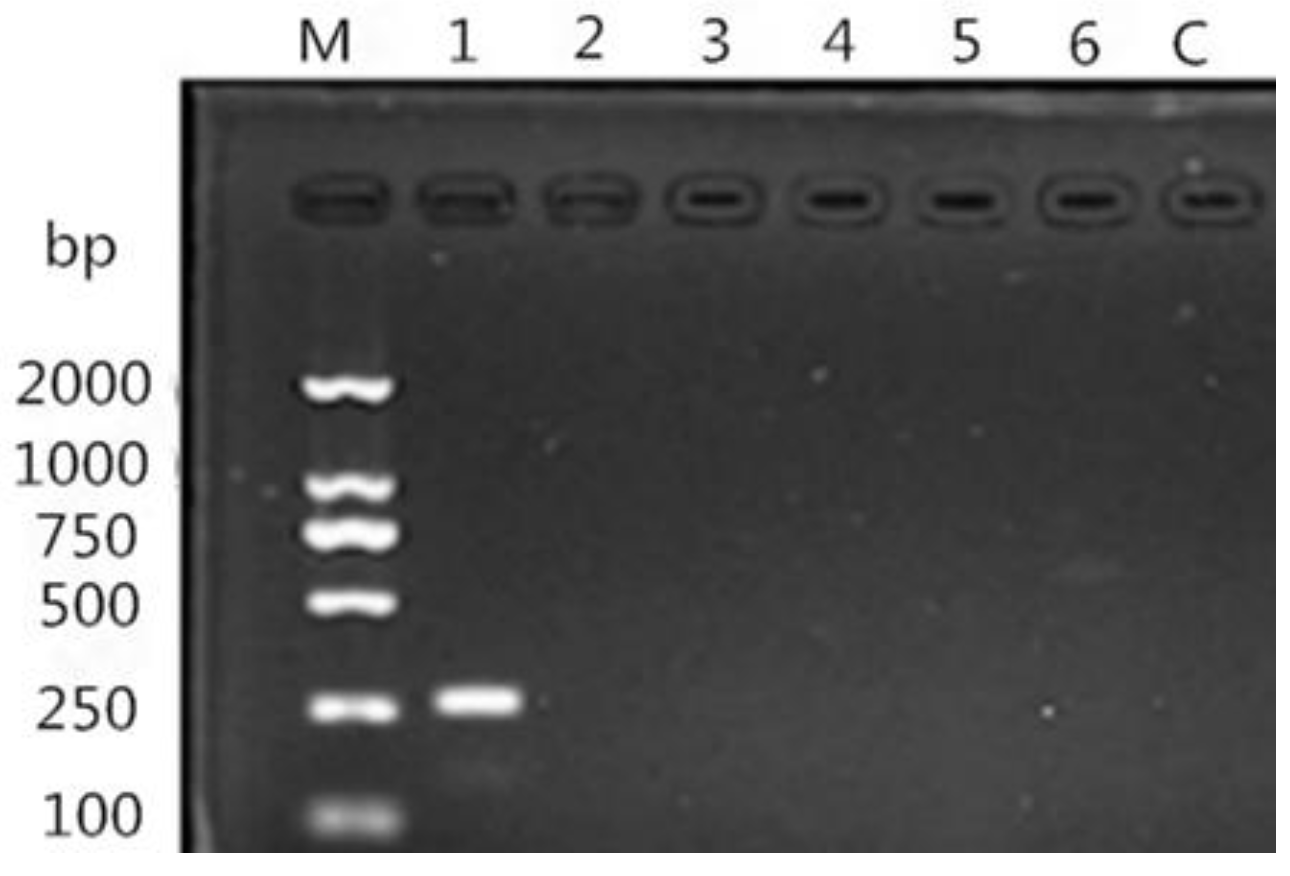
2.3.2. DNA Extraction, PCR, and Sequencing
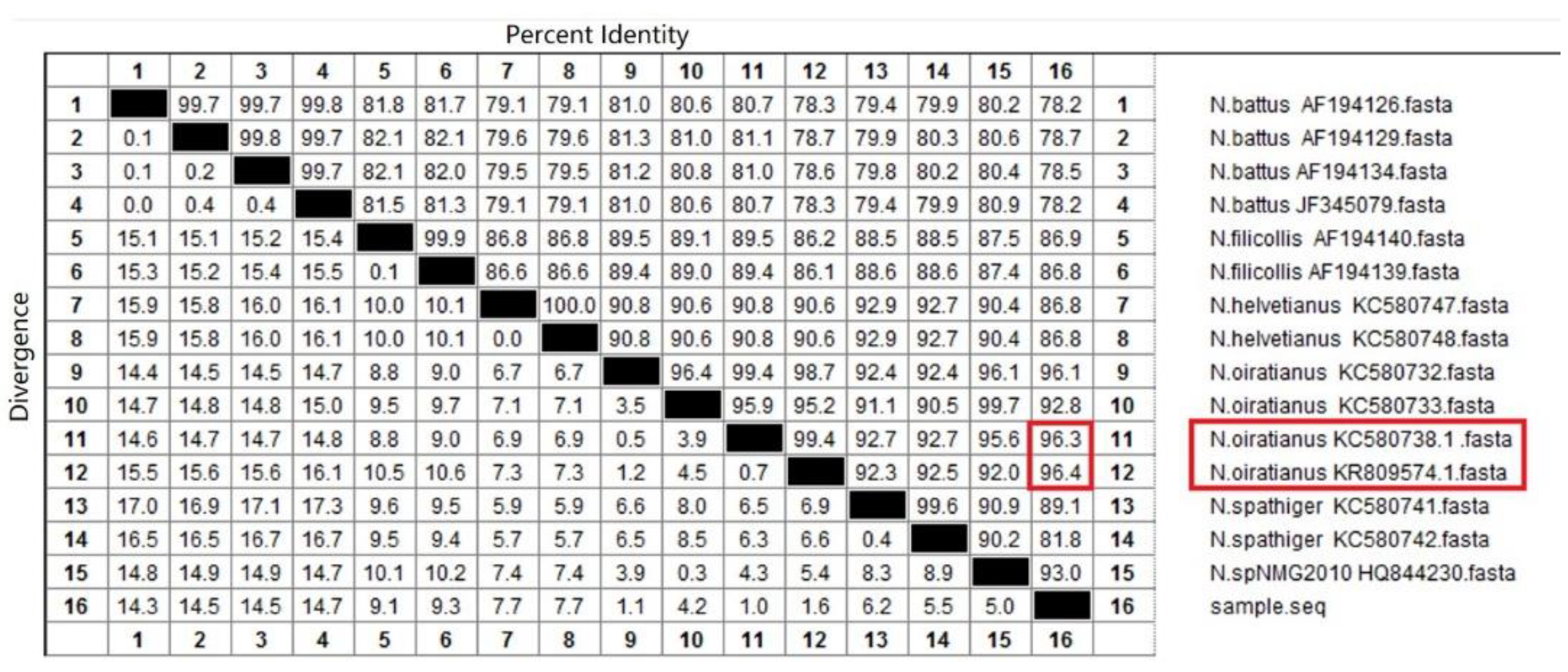
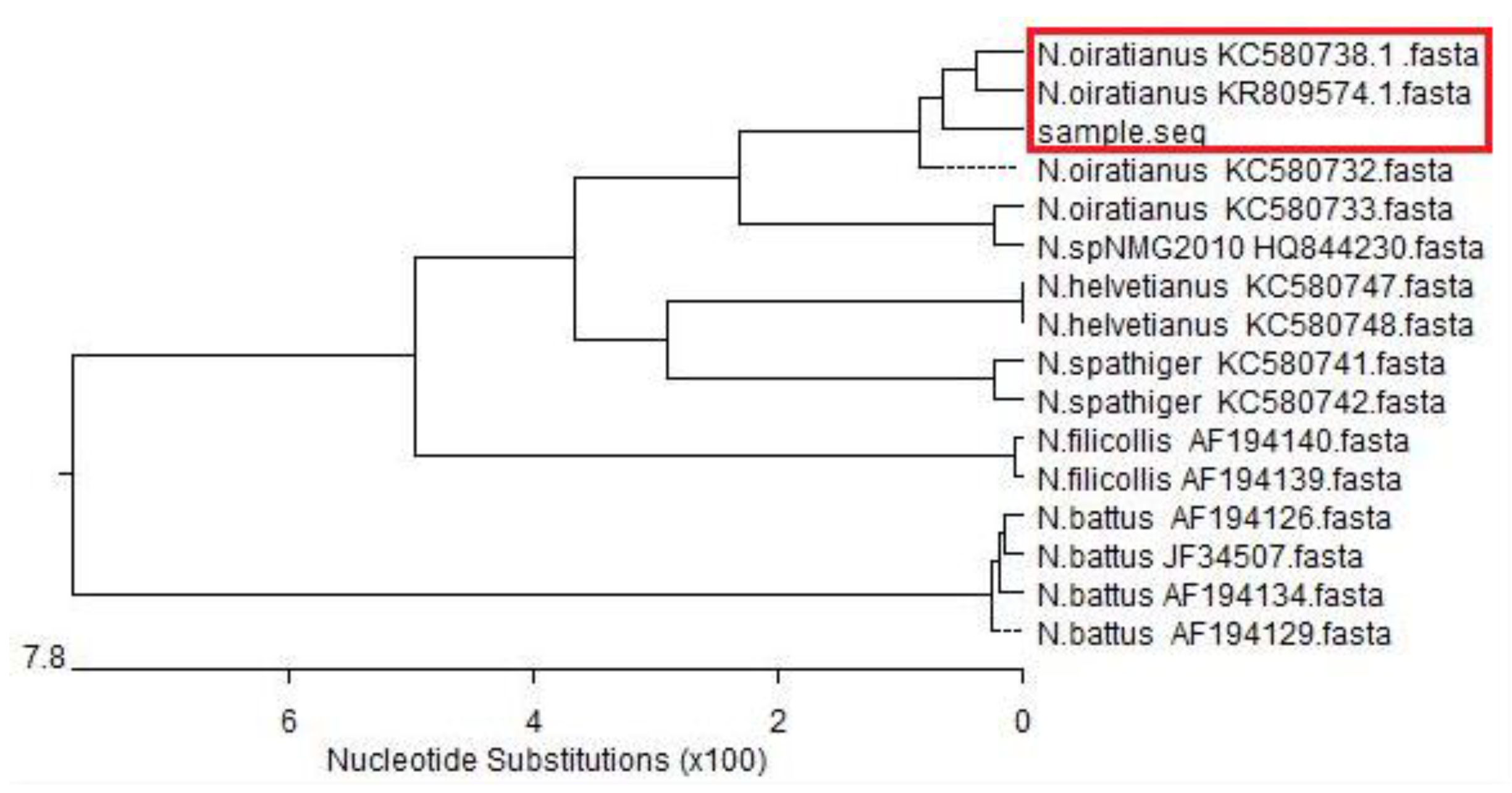
2.3.3. Sequence Analysis
2.4. Determination of Anthelmintic Resistance in Nematodirus
3. Results
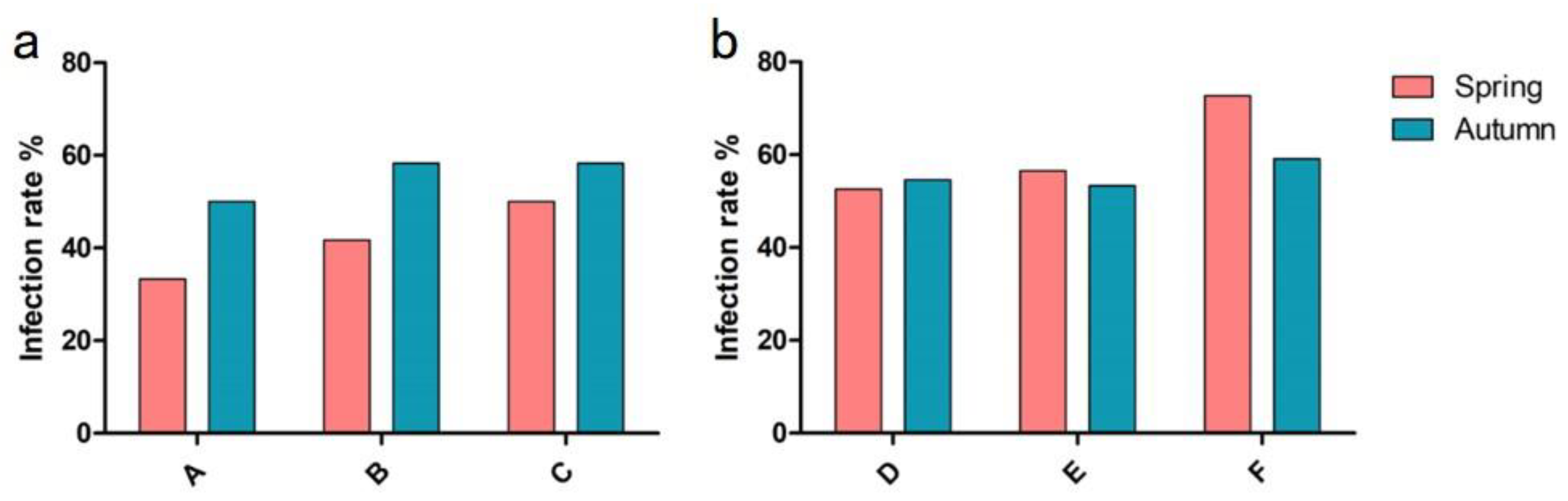
3.1. Investigation of Nematodirus Infection in Sheep
3.2. Isolation and Identification of Nematodirus
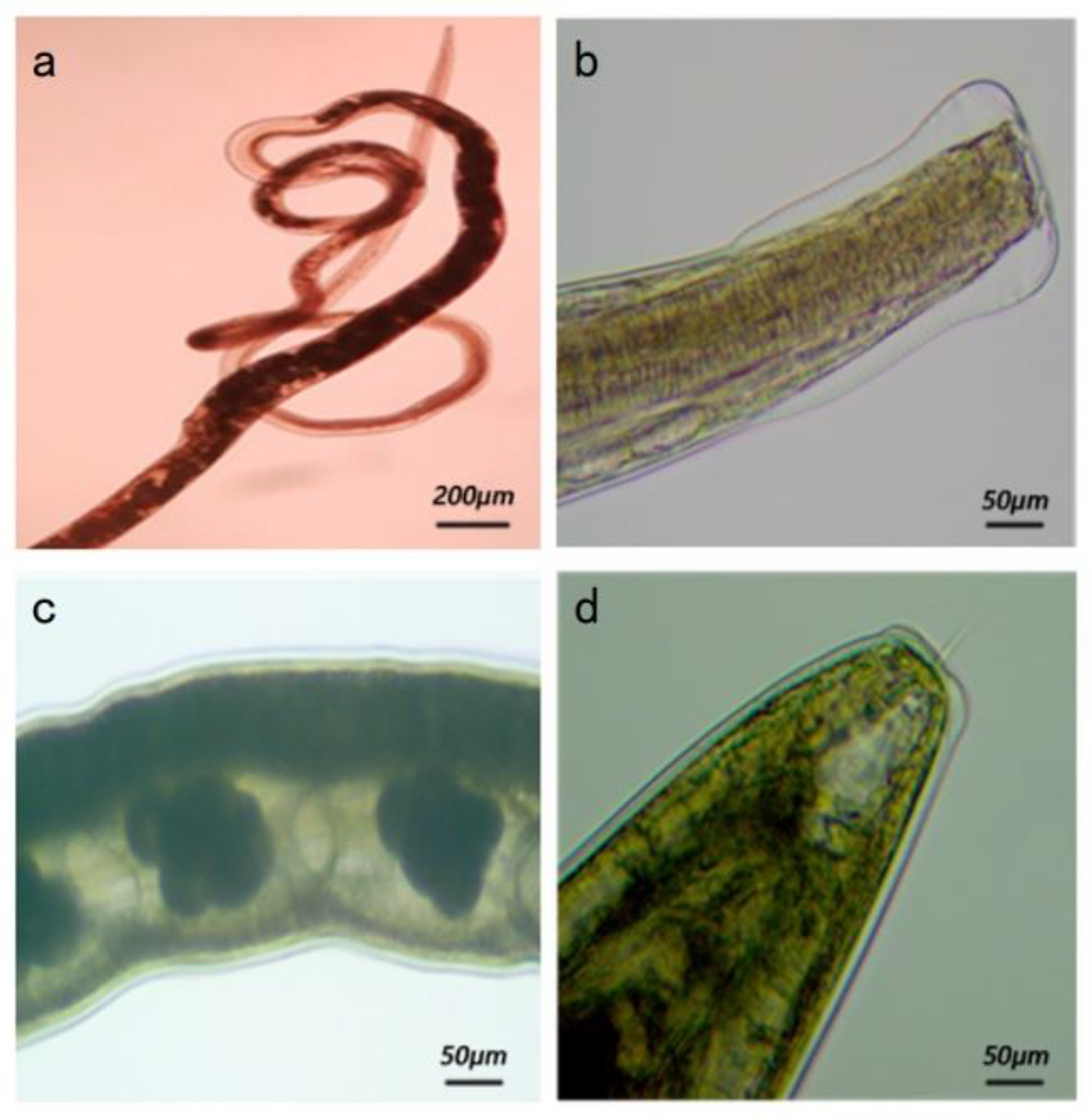
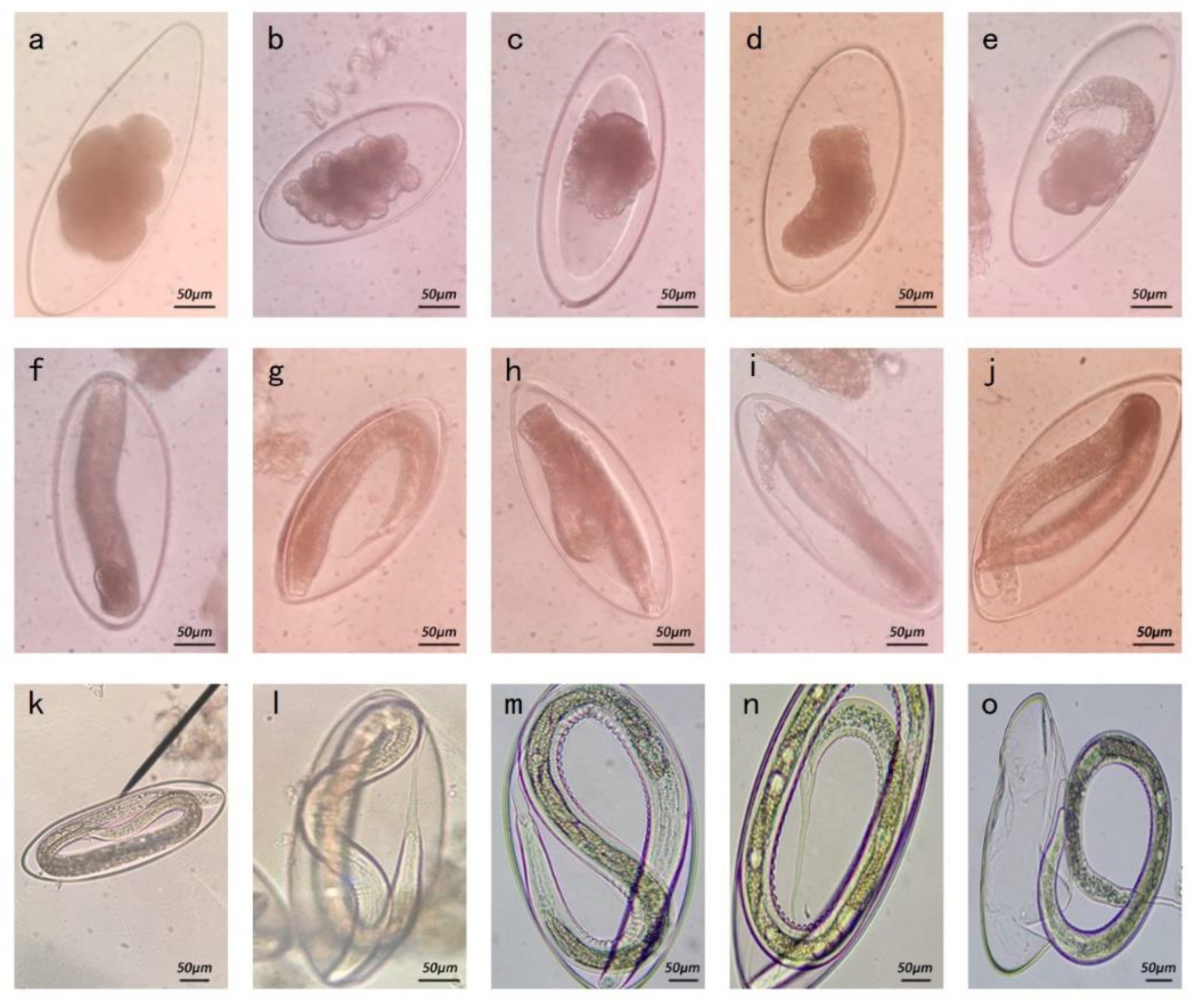
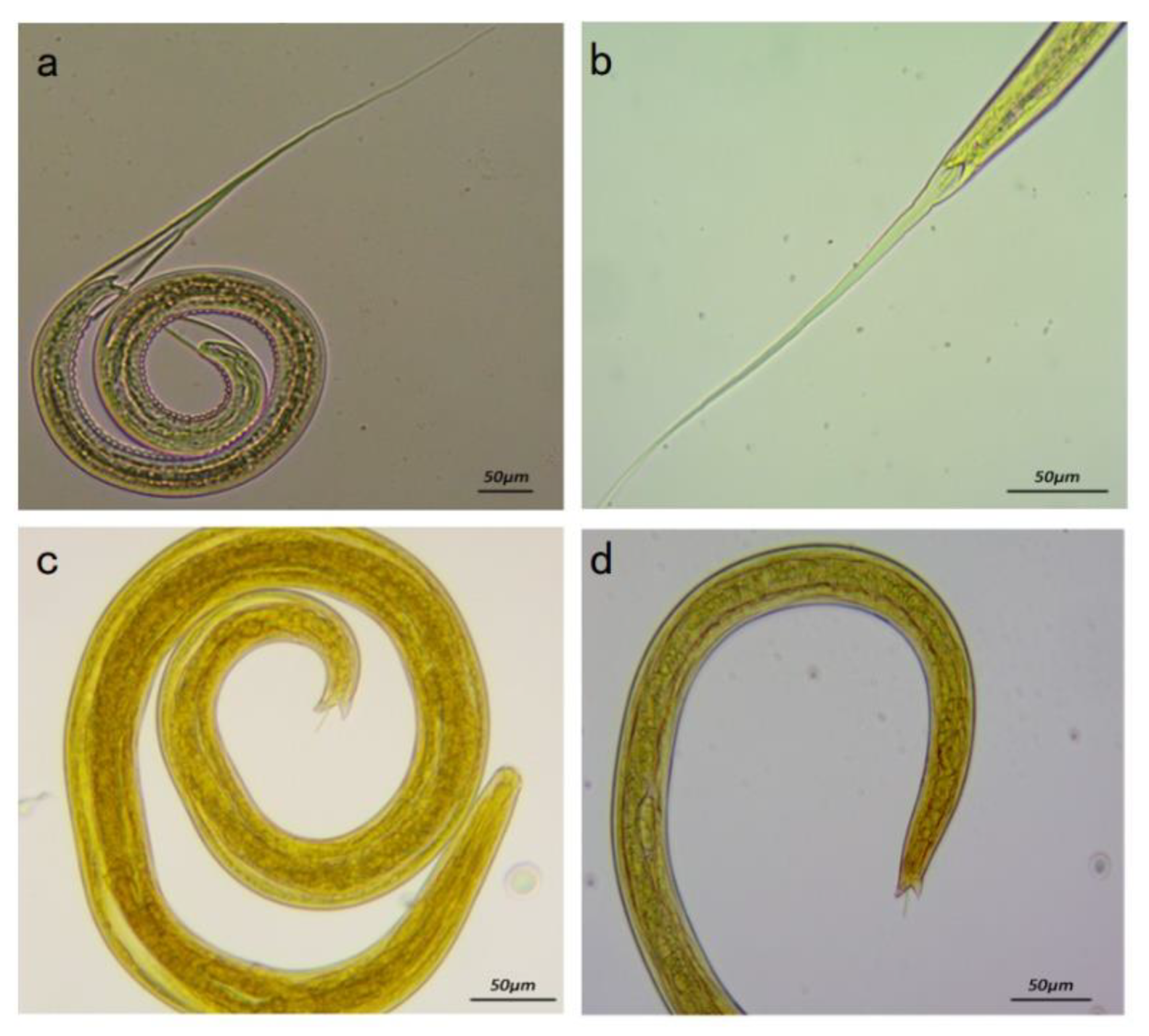
3.2.1. Morphology
3.2.2. Subsubsection
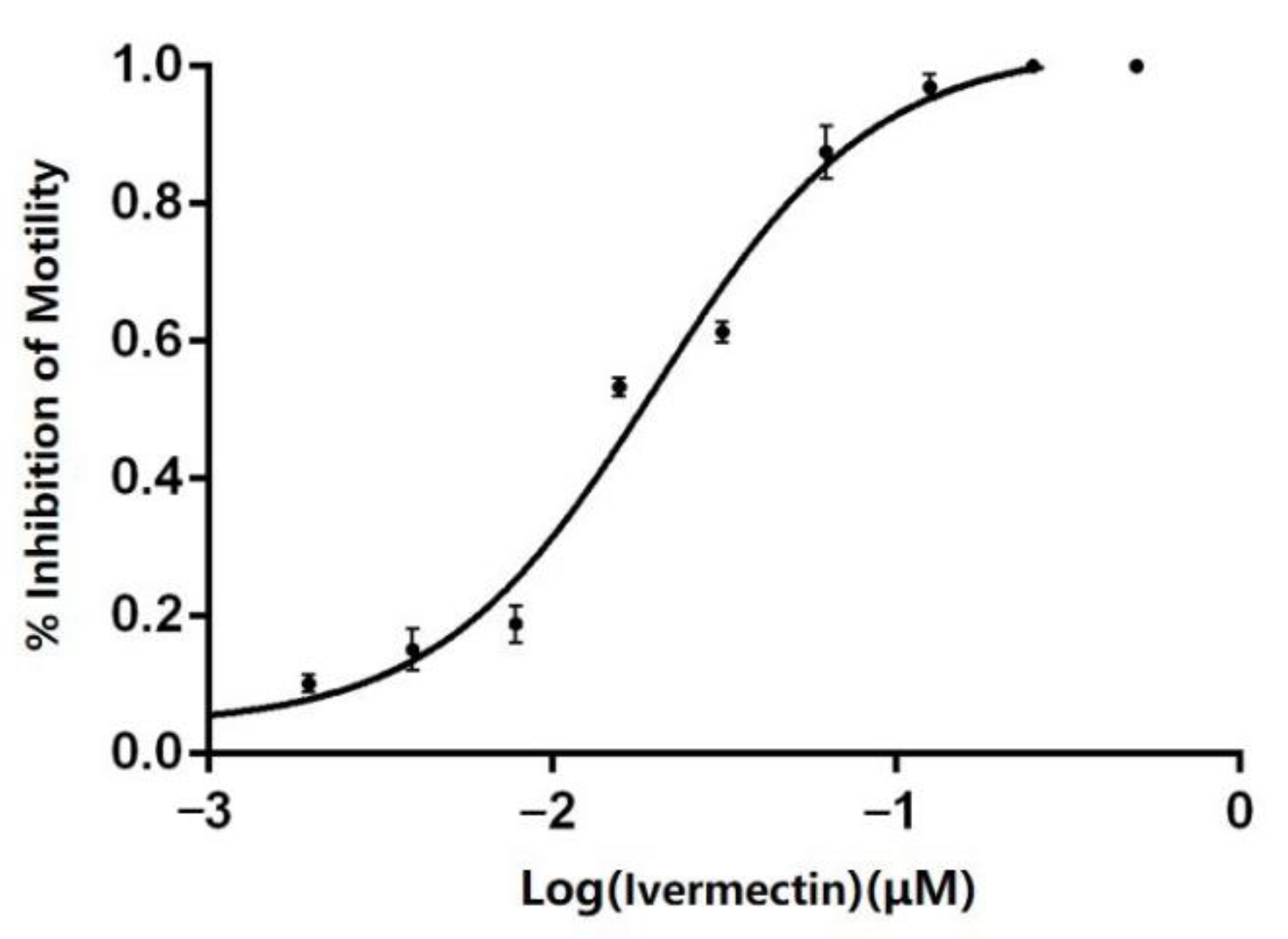
3.3. Anthelmintic Resistance Testing (LMIT)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliver, A.M.; Leathwick, D.; Pomroy, W. A survey of the prevalence of Nematodirus spathiger and N. filicollis on farms in the North and South Islands of New Zealand. N. Z. Vet. J. 2014, 62, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Hyuga, A.; Matsumoto, J. A survey of gastrointestinal parasites of alpacas (Vicugna pacos) raised in Japan. J. Vet. Med. Sci. 2016, 78, 719–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelinski, M.; Gilleard, J.; Rocheleau, L.; Royan, G.; Waldner, C. Epidemiology of gastrointestinal nematode infections in grazing yearling beef cattle in Saskatchewan. Can. Vet. J. 2017, 58, 1044–1050. [Google Scholar]
- Eye, L.; Cao, Z.F.; Hai, Y.; Miqid; Maliya, Q.; Hasi, S. Studies on infections and drug resistance of gastrointestinal nematodesin sheep in Uxin Banner. Chin. Vet. Sci. 2018, 48, 735–742. [Google Scholar]
- Oliver, A.M.; Pomroy, W.; Leathwick, D. Benzimidazole resistance in Nematodirus spathiger and N. filicollis in New Zealand. N. Z. Vet. J. 2016, 64, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Wang, M.; Zhang, G.; Han, D.; Li, X.; Liu, G.; Li, X.; Wang, Z. Gastrointestinal nematodes infections and anthelmintic resistance in grazing sheep in the Eastern Inner Mongolia in China. Acta Parasitol. 2017, 62, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, M.T.; Morenikeji, O.A.; Emikpe, B.O.; Jarikre, T.A. Interactions between gastrointestinal parasitism and pneumonia in Nigerian goats. J. Parasit. Dis. 2017, 41, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Demeler, J.; Kleinschmidt, N.; Küttler, U.; Koopmann, R.; von Samson-Himmelstjerna, G. Evaluation of the egg hatch assay and the larval migration inhibition assay to detect anthelmintic resistance in cattle parasitic nematodes on farms. Parasitol. Int. 2012, 61, 614–618. [Google Scholar] [CrossRef]
- Luo, X.; Shi, X.; Yuan, C.; Ai, M.; Ge, C.; Hu, M.; Feng, X.; Yang, X. Genome-wide SNP analysis using 2b-RAD sequencing identifies the candidate genes putatively associated with resistance to ivermectin in Haemonchus contortus. Parasites Vectors 2017, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Choubisa, S.L.; Jaroli, V.J. Gastrointestinal parasitic infection in diverse species of domestic ruminants inhabiting tribal rural areas of southern Rajasthan, India. J. Parasit. Dis. 2013, 37, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Gasser, R.B.; Chilton, N.B.; Hoste, H.; Beveridge, I. Rapid sequencing of rDNA from single worms and eggs of parasitic helminths. Nucleic. Acids. Res. 1993, 21, 2525–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.Q. Molecular Identification and Genetic Variation in Nematodirus. Master’s Thesis, Northwest A&F University, Xi’an, China, 2014. [Google Scholar]
- Demeler, J.; Küttler, U.; El-Abdellati, A.; Stafford, K.; Rydzik, A.; Varady, M.; Kenyon, F.; Coles, G.; Höglund, J.; Jackson, F.; et al. Standardization of the larval migration inhibition test for the detection of resistance to ivermectin in gastrointestinal nematodes of ruminants. Vet. Parasitol. 2010, 174, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Shen, J. Classification Map of Chinese Livestock and Poultry Parasite Morphology; China Agricultural Science and Technology Press: Beijing, China, 2006. [Google Scholar]
- Ge, P. Morphological Identification of Parasitic Nematodes in Cattle and Sheep; Xinjiang Uygur Autonomous Region Veterinary Epidemic Prevention Station: Urumqi, China, 1983. [Google Scholar]
- Tetley, J.H. Studies on Nematodirus species in sheep in Manawatu district, New Zealand. J. Helminthol. 1935, 13, 41–58. [Google Scholar] [CrossRef]
- van Wyk, J.A.; Cabaret, J.; Michael, L.M. Morphological identification of nematode larvae of small ruminants and cattle simplified. Vet. Parasitol. 2004, 119, 277–306. [Google Scholar] [CrossRef]
- van Wyk, J.A.; Mayhew, E. Morphological identification of parasitic nematode infective larvae of small ruminants and cattle: A practical lab guide. Onderstepoort J. Vet. Res. 2013, 80, E1–E14. [Google Scholar] [CrossRef] [Green Version]
- Douch, P.G.; Morum, P.E. The effects of anthelmintics on ovine larval nematode parasite migration in vitro. Int. J. Parasitol. 1994, 24, 321–326. [Google Scholar] [CrossRef]
- Boulenger, C.L. The life history of Nematodirus filicollis Rud. a nematode parasite of the sheep’s intestine. Parasitology 1915, 8, 133–155. [Google Scholar] [CrossRef] [Green Version]
- Dikmans, G.; Andrews, J.S. A comparative morphological study of the infective larvae of the common nematodes parasitic in the alimentary tract of sheep. Trans. Am. Microsc. Soc. 1933, 52, 1–25. [Google Scholar] [CrossRef]
- Wang, F.H.; Wang, B.B.; Cai, K.Z. A simple method to establish a monospecific isolate of Haemonchus contortus. Acta Agric. Boreali-Occident. Sin. 2017, 26, 1085–1089. [Google Scholar]
- Wang, F.H.; Wang, B.B.; Cai, K.Z. Establishment of a monospecific isolate of Trichostrongylus colubriformis. J. Gansu Agric.Univ. 2017, 52, 7–12. [Google Scholar]
- Zhao, S.H.; Zheng, R.K.; Geng, W.H.; Gao, H.Y. Establishment of a model of single-species Nematodirus infection in sheep. Inn. Mong. J. Anim. Sci. Prod. 1995, 16, 36–37. [Google Scholar]
- Peng, D.L. Chinese Nematology Research; China Agricultural Science and Technology Press: Beijing, China, 2016. [Google Scholar]
- Healey, A.; Mitchell, R.; Upcroft, J.A.; Boreham, P.F.; Upcroft, P. Complete nucleotide sequence of the ribosomal RNA tandem repeat unit from Giardia intestinalis. Nucleic Acids Res. 1990, 18, 4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C. Differentiation of Bunostomum phlebotomum and B. trigonocephalum by PCR-RFLP and Studies of Their Complete Mitochondrial Genome Sequences. Ph.D. Thesis, Jilin Agricultural University, Changchun, China, 2013. [Google Scholar]
- Zhao, G.H.; Jia, Y.Q.; Bian, Q.Q.; Nisbet, A.J.; Cheng, W.Y.; Liu, Y.; Fang, Y.Q.; Ma, X.T.; Yu, S.K. Molecular approaches to differentiate three species of Nematodirus in sheep and goats from China based on internal transcribed spacer rDNA sequences. J. Helminthol. 2015, 89, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, M.; Berry, K.; Graciano, S.; Becker, B.; Reuter, J.D. False-positive results after environmental pinworm PCR testing due to Rhabditid nematodes in Corncob bedding. J. Am. Assoc. Lab. Anim. Sci. 2014, 3, 717–724. [Google Scholar]
- Coles, G.C.; Jackson, F.; Pomroy, W.E.; Prichard, R.K.; von Samson-Himmelstjerna, G.; Silvestre, A.; Taylor, M.A.; Vercruysse, J. The detection of anthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 2006, 136, 167–185. [Google Scholar] [CrossRef] [PubMed]
- Middelberg, A.; McKenna, P.B. Oxfendazole resistance in Nematodirus spathiger. N. Z. Vet. J. 1983, 31, 65–68. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Longitudinal Diameter of Eggs (μm) | Transverse Diameter of Eggs (μm) | Reference |
|---|---|---|---|
| Isolated strain | 261 (218–312) | 111 (102–121) | Present study |
| N. oiratianus | 255–272 | 119–153 | [15] |
| N. spathiger | 255–272 | 90–105 | [16] |
| N. filicollis | 140–165 | 70–85 | [16] |
| N. abnormalis | 130–220 | 90–119 | [15] |
| N. helvatianus | 160–230 | 85–121 | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, P.; Wang, R.; Li, J.; Zhai, B.; Luo, X.; Yang, X. An Epidemiological Investigation and Drug-Resistant Strain Isolation of Nematodirus oiratianus in Sheep in Inner Mongolia, China. Animals 2023, 13, 30. https://doi.org/10.3390/ani13010030
Liu Y, Wang P, Wang R, Li J, Zhai B, Luo X, Yang X. An Epidemiological Investigation and Drug-Resistant Strain Isolation of Nematodirus oiratianus in Sheep in Inner Mongolia, China. Animals. 2023; 13(1):30. https://doi.org/10.3390/ani13010030
Chicago/Turabian StyleLiu, Yang, Penglong Wang, Rui Wang, Junyan Li, Bintao Zhai, Xiaoping Luo, and Xiaoye Yang. 2023. "An Epidemiological Investigation and Drug-Resistant Strain Isolation of Nematodirus oiratianus in Sheep in Inner Mongolia, China" Animals 13, no. 1: 30. https://doi.org/10.3390/ani13010030