
Aggression in Group-Housed Male Mice: A Systematic Review

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Search
2.2. Inclusion and Exclusion Criteria
2.3. Data Extraction
2.4. Data Analysis
3. Results
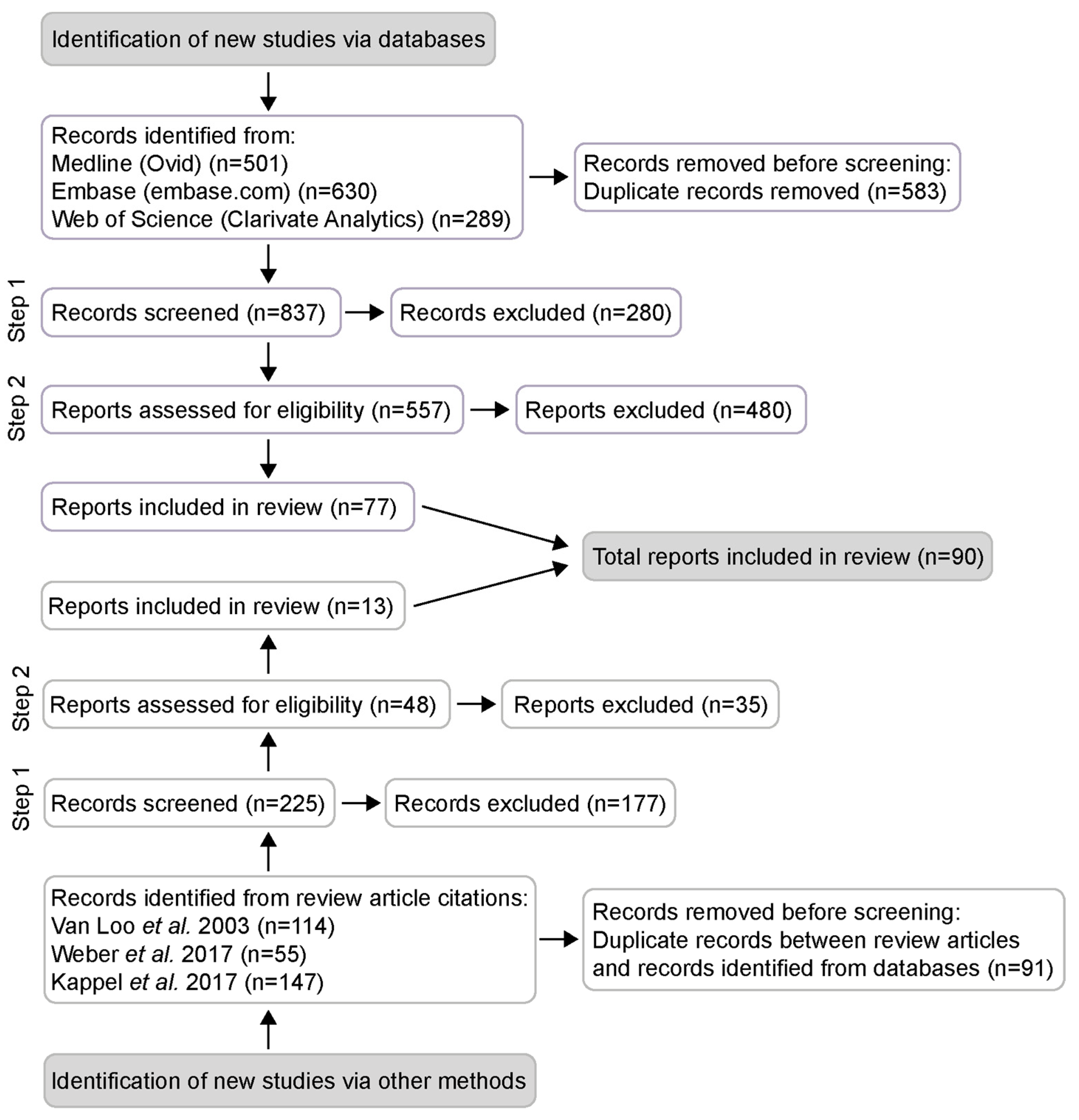
3.1. Search and Study Selection
3.2. Description of Data Set
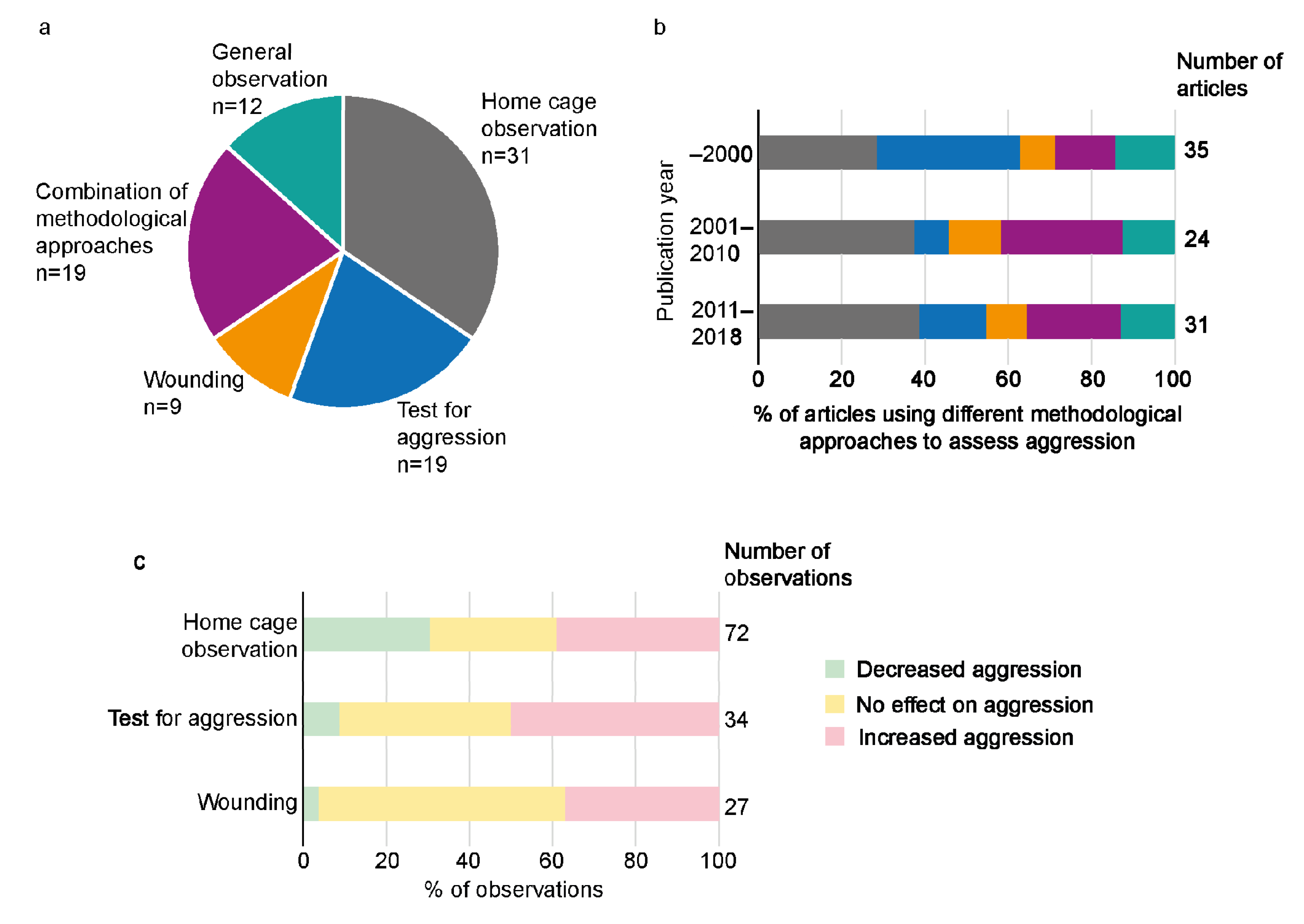
3.3. Different Methodological Approaches Used to Study Aggression
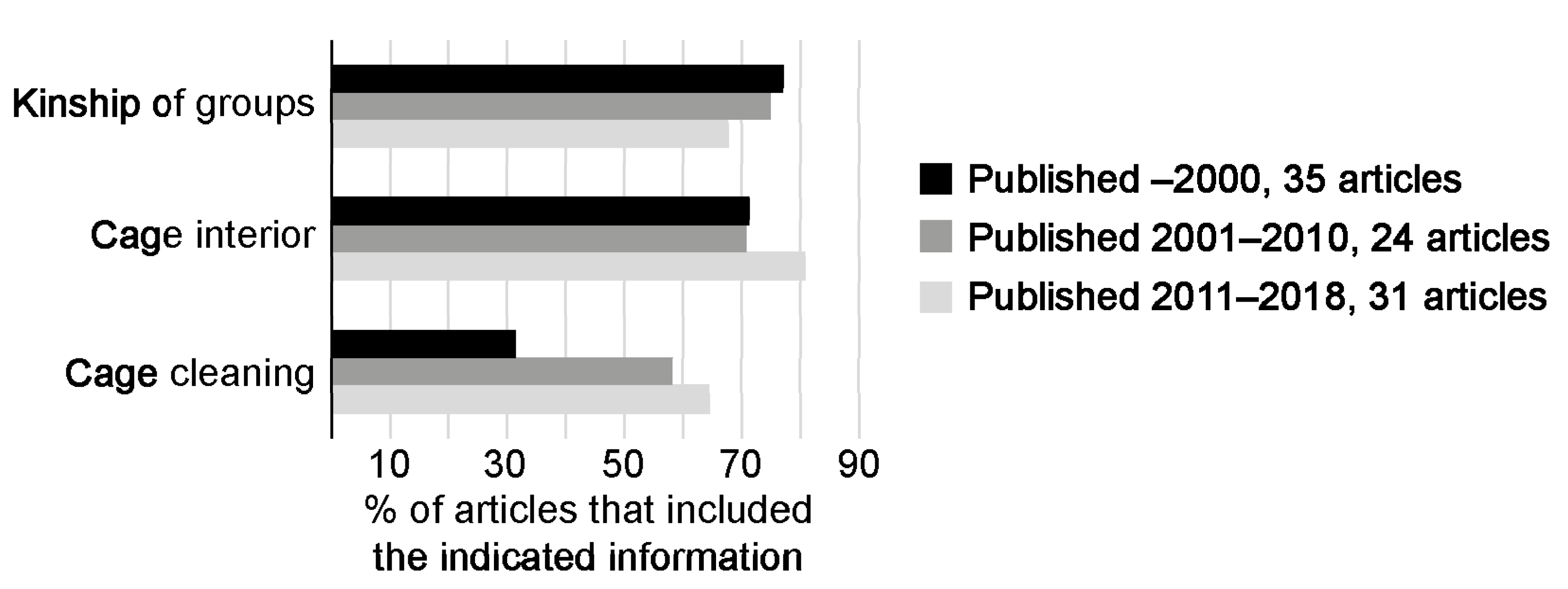
3.4. Details and Description of Housing and Husbandry
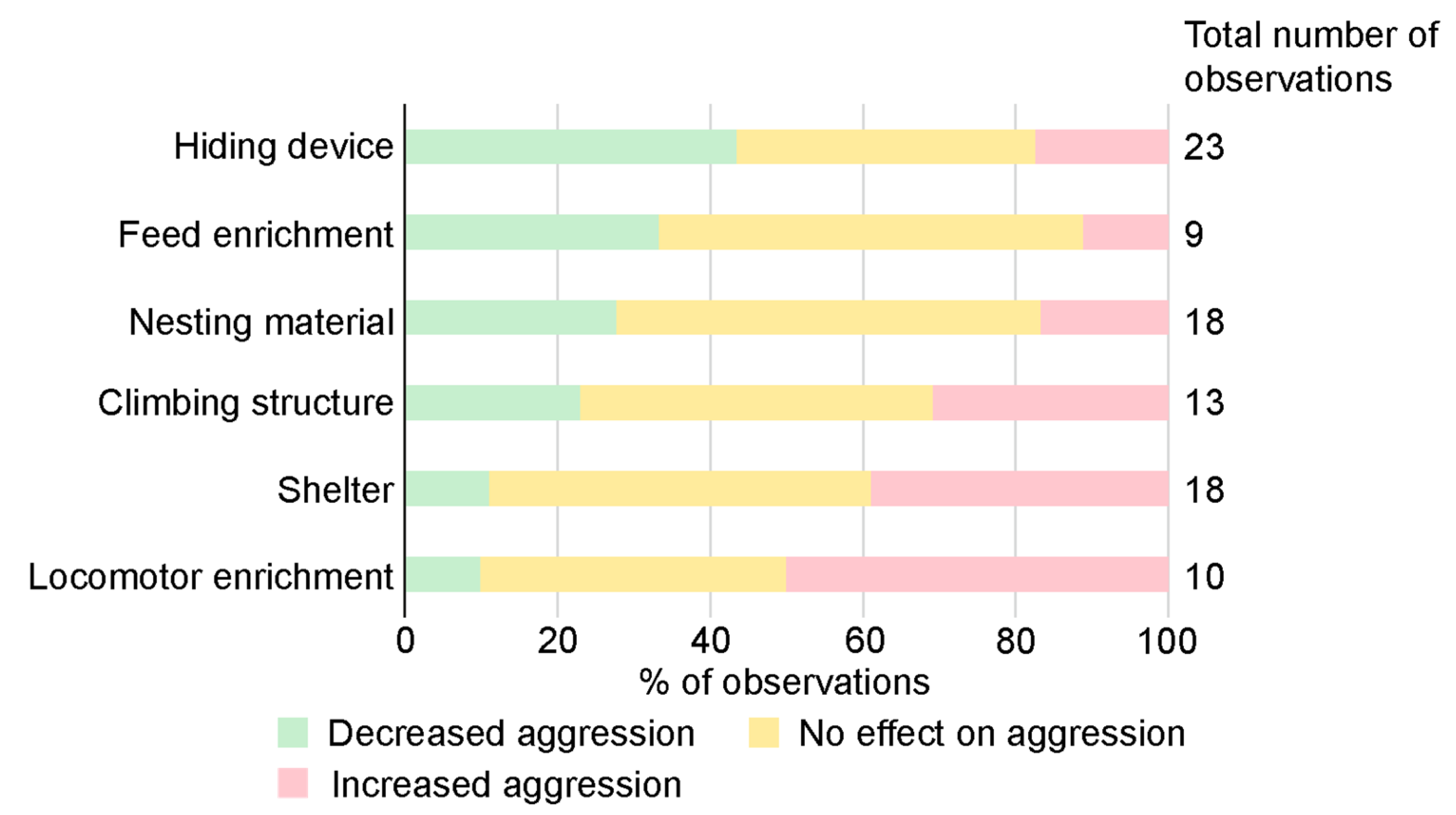
3.5. The Effect of Enrichment on Aggression
3.6. Aggression Observed over Time
3.7. Strain Differences in Aggression
3.8. Size and Group Composition
3.9. Housing Conditions and Male Mouse Aggression
4. Discussion
4.1. Methods Used for Assessment of Aggressive Behaviour
4.2. Environmental Enrichment
4.3. Strain
4.4. Group Formation
4.5. Recommendations Based on Results from the Systematic Review
- Enrichment: Use nesting material and other enrichment items that the mice can manipulate and work with. Use hiding devices where the mice can hide. Be careful when using enrichments that can be monopolized by one or a few individuals Note that environmental enrichment seem to be even more important in non-sibling groups.
- Strain: Use less aggressive strains whenever possible. This literature review indicates that C57BL/6 and BALB/c show low prevalence of aggression, in line with recently published advice [3].
- Solutions may vary: Diverging and varying opinions exist between research facilities concerning what works to prevent aggression. Try different solutions and adjust to specific needs in your lab. Be aware that the same solution might not work for all strains and research projects.
- Methods used to study aggression: Be aware that the method used to study aggression could influence the relevance of the results for home cage aggression. It is therefore important to evaluate the method used to study aggression before drawing conclusions on whether or not the reported results should be implemented in daily practice. Recent studies using home cage observations could provide more reliable information.
4.6. Important Notes for Future Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kappel, S.; Hawkins, P.; Mendl, M.T. To Group or Not to Group? Good Practice for Housing Male Laboratory Mice. Anim. Open Access J. MDPI 2017, 7, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, E.M.; Dallaire, J.A.; Gaskill, B.N.; Pritchett-Corning, K.R.; Garner, J.P. Aggression in group-housed laboratory mice: Why can’t we solve the problem? Lab Anim. 2017, 46, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lidster, K.; Owen, K.; Browne, W.J.; Prescott, M.J. Cage aggression in group-housed laboratory male mice: An international data crowdsourcing project. Sci. Rep. 2019, 9, 15211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zidar, J.; Weber, E.M.; Ewaldsson, B.; Tjäder, S.; Lilja, J.; Mount, J.; Svensson, C.; Svensk, E.; Udén, E.; Törnqvist, A.E. Group and Single Housing of Male Mice: Collected Experiences from Research Facilities in Sweden. Animals 2019, 9, 1010. [Google Scholar] [CrossRef] [Green Version]
- Theil, J.H.; Ahloy-Dallaire, J.; Weber, E.M.; Gaskill, B.N.; Pritchett-Corning, K.R.; Felt, S.A.; Garner, J.P. The epidemiology of fighting in group-housed laboratory mice. Sci. Rep. 2020, 10, 16649. [Google Scholar] [CrossRef]
- EN. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Off. J. Eur. Union 2010, 276, 33–76. [Google Scholar]
- National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [CrossRef]
- UK Home Office Code of Practice for the Housing and Care of Animals Bred, Supplied or Used for Scientific Purposes Rodents and Rabbits. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/389807/44389_unact_animals_Rodents_and_Rabbits.pdf (accessed on 23 December 2022).
- Van Loo, P.L.P.; Van Zutphen, L.F.M.; Baumans, V. Male management: Coping with aggression problems in male laboratory mice. Lab. Anim. 2003, 37, 300–313. [Google Scholar] [CrossRef] [Green Version]
- Gaskill, B. Aggression in Laboratory Mice: Potential Influences and How to Manage It. Enrich. Rec. 2014, 18, 22–25. [Google Scholar]
- Koolhaas, J.M.; Coppens, C.M.; de Boer, S.F.; Buwalda, B.; Meerlo, P.; Timmermans, P.J. The resident-intruder paradigm: A standardized test for aggression, violence and social stress. J. Vis. Exp. JoVE 2013, e4367. [Google Scholar] [CrossRef] [Green Version]
- Golden, S.A.; Covington, H.E.; Berton, O.; Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 2011, 6, 1183–1191. [Google Scholar] [CrossRef]
- Terranova, M.L.; Laviola, G.; de Acetis, L.; Alleva, E. A description of the ontogeny of mouse agonistic behavior. J. Comp. Psychol. 1998, 112, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Lindzey, G.; Winston, H.; Manosevitz, M. Social dominance in inbred mouse strains. Nature 1961, 191, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, C.C.; Akaeze, H.; Ndlebe, I.; Goodwin, N.; Eagle, A.L.; Moon, K.; Bender, A.R.; Golden, S.A.; Robison, A.J. Quantitative standardization of resident mouse behavior for studies of aggression and social defeat. Neuropsychopharmacology 2021, 46, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Gaskill, B.N.; Stottler, A.; Pritchett-Corning, K.R.; Wong, L.K.; Geronimo, J.; Garner, J.P. He’s getting under my skin! Comparing the sensitivity and specificity of dermal vs subcuticular lesions as a measure of aggression in mice. Appl. Anim. Behav. Sci. 2016, 183, 77–85. [Google Scholar] [CrossRef]
- Olsson, I.A.; Dahlborn, K. Improving housing conditions for laboratory mice: A review of “environmental enrichment”. Lab. Anim. 2002, 36, 243–270. [Google Scholar] [CrossRef] [Green Version]
- Van Loo, P.L.P.; Kruitwagen, C.; Koolhaas, J.M.; Van de Weerd, H.A.; Van Zutphen, L.F.M.; Baumans, V. Influence of cage enrichment on aggressive behaviour and physiological parameters in male mice. Appl. Anim. Behav. Sci. 2002, 76, 65–81. [Google Scholar] [CrossRef] [Green Version]
- Howerton, C.L.; Garner, J.P.; Mench, J.A. Effects of a running wheel-igloo enrichment on aggression, hierarchy linearity, and stereotypy in group-housed male CD-1 (ICR) mice. Appl. Anim. Behav. Sci. 2008, 115, 90–103. [Google Scholar] [CrossRef]
- Van Loo, P.L.P.; Van der Meer, E.; Kruitwagen, C.; Koolhaas, J.M.; Van Zutphen, L.F.M.; Baumans, V. Strain-specific aggressive behavior of male mice submitted to different husbandry procedures. Aggress. Behav. 2003, 29, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Giles, J.M.; Whitaker, J.W.; Moy, S.S.; Fletcher, C.A. Effect of environmental enrichment on aggression in BALB/cJ and BALB/cByJ mice monitored by using an automated system. J. Am. Assoc. Lab. Anim. Sci. 2018, 57, 236–243. [Google Scholar] [CrossRef]
- Swetter, B.J.; Karpiak, C.P.; Cannon, J.T. Separating the effects of shelter from additional cage enhancements for group-housed BALB/cJ mice. Neurosci. Lett. 2011, 495, 205–209. [Google Scholar] [CrossRef]
- Abramov, U.; Puussaar, T.; Raud, S.; Kurrikoff, K.; Vasar, E. Behavioural differences between C57BL/6 and 129S6/SvEv strains are reinforced by environmental enrichment. Neurosci. Lett. 2008, 443, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, N.; Morton, D.B. The Use of Cage Enrichment to Reduce Male Mouse Aggression. J. Appl. Anim. Welf. Sci. 2000, 3, 117–125. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Bloomsmith, M.A.; Brent, L.Y.; Schapiro, S.J. Guidelines for developing and managing an environmental enrichment program for nonhuman primates. Lab. Anim. Sci. 1991, 41, 372–377. [Google Scholar] [PubMed]
- Armstrong, K.R.; Clark, T.R.; Peterson, M.R. Use of Corn-Husk Nesting Material to Reduce Aggression in Caged Mice. Contemp. Top Lab. Anim. Sci. 1998, 37, 64–66. [Google Scholar]
- Lima, F.B.; Spinelli de Oliveira, E. What is the impact of low testosterone levels on the anatomical and behavioral repertoire of long-term enriched housing of male mice? Behav. Process. 2014, 108, 57–64. [Google Scholar] [CrossRef]
- Nadiah, M.Y.N.; Saadiah, M.B.H.; Nurdiana, S. Effects of Different Resource Distribution on Behaviour and Corticosterone Level of Male Mice (Mus musculus). APCBEE Procedia 2014, 8, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Clipperton-Allen, A.E.; Ingrao, J.C.; Ruggiero, L.; Batista, L.; Ovari, J.; Hammermueller, J.; Armstrong, J.N.; Bienzle, D.; Choleris, E.; Turner, P.V. Long-Term Provision of Environmental Resources Alters Behavior but not Physiology or Neuroanatomy of Male and Female BALB/c and C57BL/6 Mice. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2015, 54, 718–730. [Google Scholar]
- Nakamura, Y.; Ueno, A.; Nunomura, Y.; Nakagaki, K.; Takeda, S.; Suzuki, K. Effects of inducing exercise on growing mice by means of three-dimensional structure in rearing environment. Exp. Anim. 2016, 65, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Haemisch, A.; Gartner, K. The cage design affects intermale aggression in small groups of male laboratory mice: Strain specific consequences on social organization, and endocrine activations in two inbred strains (DBA/2J and CBA/J). J. Exp. Anim. Sci. 1994, 36, 101–116. [Google Scholar]
- Eskola, S.; Kaliste-Korhonen, E. Aspen wood-wool is preferred as a resting place, but does not affect intracage fighting of male BALB/c and C57BL/6J mice. Lab. Anim. 1999, 33, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marashi, V.; Barnekow, A.; Ossendorf, E.; Sachser, N. Effects of different forms of environmental enrichment on behavioral, endocrinological, and immunological parameters in male mice. Horm. Behav. 2003, 43, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Marashi, V.; Barnekow, A.; Sachser, N. Effects of environmental enrichment on males of a docile inbred strain of mice. Physiol. Behav. 2004, 82, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, E.; Van Loo, P.L.P.; Baumans, V. Short-term effects of a disturbed light-dark cycle and environmental enrichment on aggression and stress-related parameters in male mice. Lab. Anim. 2004, 38, 376–383. [Google Scholar] [CrossRef]
- Kaliste, E.K.; Mering, S.M.; Huuskonen, H.K. Environmental modification and agonistic behavior in NIH/S male mice: Nesting material enhances fighting but shelters prevent it. Comp. Med. 2006, 56, 202–208. [Google Scholar]
- Hunt, C.; Hambly, C. Faecal corticosterone concentrations indicate that separately housed male mice are not more stressed than group housed males. Physiol. Behav. 2006, 87, 519–526. [Google Scholar] [CrossRef]
- Workman, J.L.; Fonken, L.K.; Gusfa, J.; Kassouf, K.M.; Nelson, R.J. Post-weaning environmental enrichment alters affective responses and interacts with behavioral testing to alter nNOS immunoreactivity. Pharmacol. Biochem. Behav. 2011, 100, 25–32. [Google Scholar] [CrossRef]
- Lockworth, C.R.; Kim, S.-J.; Liu, J.; Palla, S.L.; Craig, S.L. Effect of Enrichment Devices on Aggression in Manipulated Nude Mice. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2015, 54, 731–736. [Google Scholar]
- Gaskill, B.N.; Stottler, A.M.; Garner, J.P.; Winnicker, C.W.; Mulder, G.B.; Pritchett-Corning, K.R. The effect of early life experience, environment, and genetic factors on spontaneous home-cage aggression-related wounding in male C57BL/6 mice. Lab Anim. 2017, 46, 176–184. [Google Scholar] [CrossRef]
- Barnard, C.J.; Behnke, J.M.; Sewell, J. Environmental enrichment, immunocompetence, and resistance to Babesia microti in male mice. Physiol. Behav. 1996, 60, 1223–1231. [Google Scholar] [CrossRef]
- McGregor, P.K.; Ayling, S.J. Varied cages result in more aggression in male CFLP mice. Appl. Anim. Behav. Sci. 1990, 26, 277–281. [Google Scholar] [CrossRef]
- Haemisch, A.; Gartner, K. Effects of cage enrichment on territorial aggression and stress physiology in male laboratory mice. Acta Physiol. Scand. Suppl. 1997, 640, 73–76. [Google Scholar]
- McQuaid, R.J.; Audet, M.C.; Anisman, H. Environmental enrichment in male CD-1 mice promotes aggressive behaviors and elevated corticosterone and brain norepinephrine activity in response to a mild stressor. Stress-Int. J. Biol. Stress 2012, 15, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Gresa, P.; Perez-Martinez, A.; Redolat, R. Environmental enrichment improves novel object recognition and enhances agonistic behavior in male mice. Aggress. Behav. 2013, 39, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Leduc, R.Y.M.; Rauw, G.; Baker, G.B.; McDermid, H.E. What goes around can come around: An unexpected deleterious effect of using mouse running wheels for environmental enrichment. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 194–201. [Google Scholar]
- Haemisch, A.; Voss, T.; Gartner, K. Effects of environmental enrichment on aggressive behavior, dominance hierarchies, and endocrine states in male DBA/2J mice. Physiol. Behav. 1994, 56, 1041–1048. [Google Scholar] [CrossRef]
- McQuaid, R.J.; Audet, M.-C.; Jacobson-Pick, S.; Anisman, H. Environmental enrichment influences brain cytokine variations elicited by social defeat in mice. Psychoneuroendocrinology 2013, 38, 987–996. [Google Scholar] [CrossRef]
- McQuaid, R.J.; Audet, M.-C.; Jacobson-Pick, S.; Anisman, H. The differential impact of social defeat on mice living in isolation or groups in an enriched environment: Plasma corticosterone and monoamine variations. Int. J. Neuropsychopharmacol. 2013, 16, 351–363. [Google Scholar] [CrossRef]
- McQuaid, R.J.; Dunn, R.; Jacobson-Pick, S.; Anisman, H.; Audet, M.-C. Post-weaning Environmental Enrichment in Male CD-1 Mice: Impact on Social Behaviors, Corticosterone Levels and Prefrontal Cytokine Expression in Adulthood. Front. Behav. Neurosci. 2018, 12, 145. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, P.; Militzer, K.; Buttner, D. Environmental enrichment and aggressive behaviour: Influence on body weight and body fat in male inbred HLG mice. J. Exp. Anim. Sci. 1995, 37, 69–78. [Google Scholar]
- Page, D.L.; Glenner, G.G. Social interaction and wounding in the genesis of “spontaneous” murine amyloidosis. Am. J. Pathol. 1972, 67, 555–567. [Google Scholar]
- Poole, T.B.; Morgan, H.D.R. Differences in aggressive behaviour between male mice (Mus musculus L.) in colonies of different sizes. Anim. Behav. 1973, 21, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Poole, T.B.; Morgan, H.D.R. Social and territorial behaviour of laboratory mice (Mus musculus L.) in small complex areas. Anim. Behav. 1976, 24, 476–480. [Google Scholar] [CrossRef]
- Bishop, M.J.; Chevins, P.F.D. Urine odours and marking patterns in territorial laboratory mice (Mus musculus). Behav. Process. 1987, 15, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Burright, R.G.; Freeman, M.J.; Donovick, P.J. Repeated tests of intermale aggression in mice (Mus musculus) are influenced by housing and test conditions. J. Comp. Psychol. 1988, 102, 303–305. [Google Scholar] [CrossRef]
- Hilakivi-Clarke, L.; Dickson, R.B. Stress influence on development of hepatocellular tumors in transgenic mice overexpressing TGF alpha. Acta Oncol. 1995, 34, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Grimm, M.S.; Emerman, J.T.; Weinberg, J. Effects of social housing condition and behavior on growth of the Shionogi mouse mammary carcinoma. Physiol. Behav. 1996, 59, 633–642. [Google Scholar] [CrossRef]
- Vaughan, L.M.; Dawson, J.S.; Porter, P.R.; Whittaker, A.L. Castration promotes welfare in group-housed male Swiss outbred mice maintained in educational institutions. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2014, 53, 38–43. [Google Scholar]
- Bove, M.; Ike, K.; Eldering, A.; Buwalda, B.; de Boer, S.F.; Morgese, M.G.; Schiavone, S.; Cuomo, V.; Trabace, L.; Kas, M.J.H. The Visible Burrow System: A behavioral paradigm to assess sociability and social withdrawal in BTBR and C57BL/6J mice strains. Behav. Brain Res. 2018, 344, 9–19. [Google Scholar] [CrossRef]
- Svare, B.B.; Leshner, A.I. Behavioral correlates of intermale aggression and grouping in mice. J. Comp. Physiol. Psychol. 1973, 85, 203–210. [Google Scholar] [CrossRef]
- O’Donnell, V.; Blanchard, R.J.; Blanchard, D.C. Mouse aggression increases after 24 hours of isolation or housing with females. Behav. Neural Biol. 1981, 32, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kareem, A.M.; Barnard, C.J. The importance of kinship and familiarity in social interactions between mice. Anim. Behav. 1982, 30, 594–601. [Google Scholar] [CrossRef]
- Fairless, A.H.; Katz, J.M.; Vijayvargiya, N.; Dow, H.C.; Kreibich, A.S.; Berrettini, W.H.; Abel, T.; Brodkin, E.S. Development of home cage social behaviors in BALB/cJ vs. C57BL/6J mice. Behav. Brain Res. 2013, 237, 338–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwirner, P.P.; Porsolt, R.D.; Loew, D.M. Inter-group aggression in mice: A new method for testing the effects of centrally active drugs. Psychopharmacologia 1975, 45, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Terranova, M.L.; Laviola, G.; Alleva, E. Ontogeny of amicable social behavior in the mouse: Gender differences and ongoing isolation outcomes. Dev. Psychobiol. 1993, 26, 467–481. [Google Scholar] [CrossRef] [PubMed]
- Benus, R.F.; Henkelmann, C. Litter composition influences the development of aggression and behavioural strategy in male Mus domesticus. Behaviour 1998, 135, 1229–1249. [Google Scholar]
- Van Loo, P.L.P.; Kruitwagen, C.; Van Zutphen, L.F.M.; Koolhaas, J.M.; Baumans, V. Modulation of aggression in male mice: Influence of cage cleaning regime and scent marks. Anim. Welf. 2000, 9, 281–295. [Google Scholar]
- Van Loo, P.L.; Mol, J.A.; Koolhaas, J.M.; Van Zutphen, B.F.; Baumans, V. Modulation of aggression in male mice: Influence of group size and cage size. Physiol. Behav. 2001, 72, 675–683. [Google Scholar] [CrossRef] [Green Version]
- Garris, D.R. Aggression-associated changes in murine olfactory tubercle bioamines. Brain Res. 2003, 963, 150–155. [Google Scholar] [CrossRef]
- Sorensen, D.B.; Johnsen, P.F.; Bibby, B.M.; Bottner, A.; Bornstein, S.R.; Eisenhofer, G.; Pacak, K.; Hansen, A.K. PNMT transgenic mice have an aggressive phenotype. Horm. Metab. Res. = Horm.-Und Stoffwechs. = Horm. Et Metab. 2005, 37, 159–163. [Google Scholar] [CrossRef]
- Bisazza, A. Social organization and territorial behaviour in three strains of mice. Boll. Zool. 1981, 48, 157–167. [Google Scholar] [CrossRef]
- Mondragón, R.; Mayagoitia, L.; López-Luján, A.; Díaz, J.L. Social structure features in three inbred strains of mice, C57Bl/6J, Balb/cj, and NIH: A comparative study. Behav. Neural Biol. 1987, 47, 384–391. [Google Scholar] [CrossRef]
- Nicholson, A.; Malcolm, R.D.; Russ, P.L.; Cough, K.; Touma, C.; Palme, R.; Wiles, M.V. The response of C57BL/6J and BALB/cJ mice to increased housing density. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 740–753. [Google Scholar] [PubMed]
- Smith, A.L.; Mabus, S.L.; Muir, C.; Woo, Y. Effects of housing density and cage floor space on three strains of young adult inbred mice. Comp. Med. 2005, 55, 368–376. [Google Scholar] [PubMed]
- Babineau, B.A.; Yang, M.; Berman, R.F.; Crawley, J.N. Low home cage social behaviors in BTBR T+tf/J mice during juvenile development. Physiol. Behav. 2013, 114–115, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Fitchett, A.E.; Collins, S.A.; Mason, H.; Barnard, C.J.; Cassaday, H.J. Urinary corticosterone measures: Effects of strain and social rank in BKW and CD-1 mice. Behav. Process. 2005, 70, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Kempf, E.; Puglisi-Allegra, S.; Cabib, S.; Schleef, C.; Mandel, P. Serotonin levels and turnover in different brain areas of isolated aggressive or non-aggressive strains of mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1984, 8, 365–371. [Google Scholar] [CrossRef]
- Martinez-Cue, C.; Rueda, N.; Garcia, E.; Davisson, M.T.; Schmidt, C.; Florez, J. Behavioral, cognitive and biochemical responses to different environmental conditions in male Ts65Dn mice, a model of Down syndrome. Behav. Brain Res. 2005, 163, 174–185. [Google Scholar] [CrossRef]
- Trainor, B.C.; Workman, J.L.; Jessen, R.; Nelson, R.J. Impaired Nitric Oxide Synthase Signaling Dissociates Social Investigation and Aggression. Behav. Neurosci. 2007, 121, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Lewejohann, L.; Kloke, V.; Heiming, R.S.; Jansen, F.; Kaiser, S.; Schmitt, A.; Lesch, K.P.; Sachser, N. Social status and day-to-day behaviour of male serotonin transporter knockout mice. Behav. Brain Res. 2010, 211, 220–228. [Google Scholar] [CrossRef]
- Rodriguez-Arias, M.; Navarrete, F.; Daza-Losada, M.; Navarro, D.; Aguilar, M.A.; Berbel, P.; Minarro, J.; Manzanares, J. CB1 cannabinoid receptor-mediated aggressive behavior. Neuropharmacology 2013, 75, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Arias, M.; Navarrete, F.; Blanco-Gandia, M.C.; Arenas, M.C.; Aguilar, M.A.; Bartoll-Andres, A.; Valverde, O.; Minarro, J.; Manzanares, J. Role of CB2 receptors in social and aggressive behavior in male mice. Psychopharmacology 2015, 232, 3019–3031. [Google Scholar] [CrossRef] [PubMed]
- McNamara, G.I.; John, R.M.; Isles, A.R. Territorial Behavior and Social Stability in the Mouse Require Correct Expression of Imprinted Cdkn1c. Front. Behav. Neurosci. 2018, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Xiao, Y.; Yin, H.H.; Chen, A.I.; Soong, T.W.; Je, H.S. Postnatal TrkB ablation in corticolimbic interneurons induces social dominance in male mice. Proc. Natl. Acad. Sci. USA 2018, 115, E9909–E9915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguiz, R.M.; Chu, R.; Caron, M.G.; Wetsel, W.C. Aberrant responses in social interaction of dopamine transporter knockout mice. Behav. Brain Res. 2004, 148, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Bailoo, J.D.; Murphy, E.; Varholick, J.A.; Novak, J.; Palme, R.; Wurbel, H. Evaluation of the effects of space allowance on measures of animal welfare in laboratory mice. Sci. Rep. 2018, 8, 713. [Google Scholar] [CrossRef] [Green Version]
- Blankenberger, W.B.; Weber, E.M.; Chu, D.K.; Geronimo, J.T.; Theil, J.; Gaskill, B.N.; Pritchett-Corning, K.; Albertelli, M.A.; Garner, J.P.; Ahloy-Dallaire, J. Breaking up is hard to do: Does splitting cages of mice reduce aggression? Appl. Anim. Behav. Sci. 2018, 206, 94–101. [Google Scholar] [CrossRef]
- Welch, B.L.; Welch, A.S. Graded effect of social stimulation upon d-amphetamine toxicity, aggressiveness and heart and adrenal weight. J. Pharmacol. Exp. Ther. 1966, 151, 331–338. [Google Scholar]
- Barnard, C.J.; Behnke, J.M.; Sewell, J. Social behaviour and susceptibility to infection in house mice (Mus musculus): Effects of group size, aggressive behaviour and status-related hormonal responses prior to infection on resistance to Babesia microti. Parasitology 1994, 108 Pt 5, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Saldívar-González, J.A.; Posadas-Andrews, A.; Rojas, J.A.; Yoldi-Negrete, M.; Álvarez-Sekely, A.; Flores-Hernández, V.; Ortiz-León, S.; Foilloux-Morales, C.; Mayagoitia, L.; Mondragón-Ceballos, R. Effect of imipramine and electro convulsive stimulation in mice under social stress conditions. Curr. Top. Pharmacol. 2007, 11, 57–70. [Google Scholar]
- Bartolomucci, A.; Chirieleison, A.; Gioiosa, L.; Ceresini, G.; Parmigiani, S.; Palanza, P. Age at group formation alters behavior and physiology in male but not female CD-1 mice. Physiol. Behav. 2004, 82, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Kikusui, T.; Takeuchi, Y.; Mori, Y. Early weaning induces anxiety and aggression in adult mice. Physiol. Behav. 2004, 81, 37–42. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, I.; Alleva, E.; Branchi, I. Communal nesting, an early social enrichment, affects social competences but not learning and memory abilities at adulthood. Behav. Brain Res. 2007, 183, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.A.; Lister, R.G. Are there preexisting behavioral characteristics that predict the dominant status of male NIH Swiss mice (Mus musculus)? J. Comp. Psychol. 1992, 106, 184–189. [Google Scholar] [CrossRef]
- Kaliste-Korhonen, E.; Eskola, S. Fighting in NIH/S male mice: Consequences for behaviour in resident-intruder tests and physiological parameters. Lab. Anim. 2000, 34, 189–198. [Google Scholar] [CrossRef]
- Lee, W.; Khan, A.; Curley, J.P. Major urinary protein levels are associated with social status and context in mouse social hierarchies. Proceedings. Biol. Sci. 2017, 284, 20171570. [Google Scholar] [CrossRef]
- Williamson, C.M.; Romeo, R.D.; Curley, J.P. Dynamic changes in social dominance and mPOA GnRH expression in male mice following social opportunity. Horm. Behav. 2017, 87, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.; Hurst, J.L. The effects of cage cleaning on aggression within groups of male laboratory mice. Anim. Behav. 1995, 49, 821–826. [Google Scholar] [CrossRef]
- Smith, A.L.; Corrow, D.J. Modifications to husbandry and housing conditions of laboratory rodents for improved well-being. ILAR J. 2005, 46, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Koyama, S.; Kamimura, S. Influence of social dominance and female odor on the sperm activity of male mice. Physiol. Behav. 2000, 71, 415–422. [Google Scholar] [CrossRef]
- Wurbel, H.; Garner, J.P. Refinement of rodent research through environmental enrichment and systematic randomization. Natl. Cent. Replace. Refinement Reduct. Anim. Res. 2007, 9, 1–9. [Google Scholar]
- Ratuski, A.S.; Weary, D.M. Environmental Enrichment for Rats and Mice Housed in Laboratories: A Metareview. Animals 2022, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Olsson, I.A.S.; Westlund, K. More than numbers matter: The effect of social factors on behaviour and welfare of laboratory rodents and non-human primates. Appl. Anim. Behav. Sci. 2007, 103, 229–254. [Google Scholar] [CrossRef]
- Akre, A.K.; Bakken, M.; Hovland, A.L.; Palme, R.; Mason, G. Clustered environmental enrichments induce more aggression and stereotypic behaviour than do dispersed enrichments in female mice. Appl. Anim. Behav. Sci. 2011, 131, 145–152. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef] [PubMed]
- Azkona, G.; Caballero, J.M. Implementing strategies to reduce singly housed male mice. Lab Anim 2019, 53, 508–510. [Google Scholar] [CrossRef]
- Grífols, R.; Zamora, C.; Ortega-Saez, I.; Azkona, G. Postweaning Grouping as a Strategy to Reduce Singly Housed Male Mice. Animals 2020, 10, 2135. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Number of Articles | Number of Observations |
|---|---|---|
| Enrichment | 33 | 64 |
| Time in the group | 27 | 38 |
| Strain | 26 | 35 |
| Group formation | 23 | 30 |
| Housing condition | 16 | 23 |
| Other | 7 | 8 |
| Methodological Approach Used to Study Aggression | ||||||
|---|---|---|---|---|---|---|
| Less Aggressive | More Aggressive | Home Cage Behavior | Test For Aggression | Wounding | General Observation | Reference |
| C57BL/6 | BALB/c | x | [75] | |||
| C57BL/6 | BALB/c | x | x | [73] | ||
| C57BL/6 | BALB/c | x | [74] | |||
| C57BL/6 | 129S a | x | [23] | |||
| C57BL/6 | Swiss | x | x | [73] | ||
| C57BL/6 | NIH | x | [74] | |||
| BALB/c | CD-1 b | x | x | [20] | ||
| BALB/c | FVB | x | x | [76] | ||
| BALB/c | Swiss | x | x | [73] | ||
| BALB/c | NIH | x | [74] | |||
| BALB/c | C57BL/6 | x | x | x | [33] | |
| CBA/J | DBA2 | x | [32] | |||
| CD-1 | BKW | x | [78] | |||
| NOD | FVB | x | x | [76] | ||
| No Difference | Home Cage Behavior | Test for Aggression | Wounding | General Observation | Reference | |
| C57BL/6 | 129S c | x | [23] | |||
| C57BL/6 | BALB/c | x | [30] | |||
| C57BL/6 | BALB/c | x | [65] | |||
| C57BL/6 | BTBR | x | [77] | |||
| C57BL/6 | BTBR | x | [61] | |||
| C57BL/6 d | BTBR | x | [77] | |||
| C57BL/6 | DBA/2 | x | [79] | |||
| BALB/c | NOD | x | x | [76] | ||
| Group Size | Methodological Approach Used to Study Aggression | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | 2 | 3 | 4 | 5 | 6 | 8 | 9 | 10 | 12 | 20 | Home Cage Behavior | Test for Aggression | Wounding | General Observation | Reference |
| BALB/c | x | x | x | x | x | [70] | |||||||||
| BALB/c | x | x | x | x | [88] | ||||||||||
| Varying strains | x | x | x | x | x | x | [89] | ||||||||
| Randomly bred | x | x | x | x | x | x | [54] | ||||||||
| DD/S strain | x | x | x | [59] | |||||||||||
| MF1 | x | x | x | [38] | |||||||||||
| BALB/c | x | x | x | [92] | |||||||||||
| CFLP | x | x | x | x | [42] | ||||||||||
| CFLP | x | x | x | x | [91] | ||||||||||
| DUB/ICR Swiss | x | x | x | x | x | [90] | |||||||||
| DUB/ICR Swiss | x | x | x | x | x | x | [90] | ||||||||
| DUB/ICR Swiss | x | x | x | x | x | [90] | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, E.M.; Zidar, J.; Ewaldsson, B.; Askevik, K.; Udén, E.; Svensk, E.; Törnqvist, E. Aggression in Group-Housed Male Mice: A Systematic Review. Animals 2023, 13, 143. https://doi.org/10.3390/ani13010143
Weber EM, Zidar J, Ewaldsson B, Askevik K, Udén E, Svensk E, Törnqvist E. Aggression in Group-Housed Male Mice: A Systematic Review. Animals. 2023; 13(1):143. https://doi.org/10.3390/ani13010143
Chicago/Turabian StyleWeber, Elin M., Josefina Zidar, Birgit Ewaldsson, Kaisa Askevik, Eva Udén, Emma Svensk, and Elin Törnqvist. 2023. "Aggression in Group-Housed Male Mice: A Systematic Review" Animals 13, no. 1: 143. https://doi.org/10.3390/ani13010143