
miR-27a Targeting PIK3R3 Regulates the Proliferation and Apoptosis of Sheep Hair Follicle Stem Cells
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture of HFSCs
2.2. Cell Transfection
2.3. Determination of Gene Expression
2.4. Dual Luciferase Reporter Assay
2.5. EdU and CCK-8 Assays
2.6. Detection of Apoptosis by Flow Cytometry
2.7. Western Blot Assay
2.8. Data Analysis
3. Results
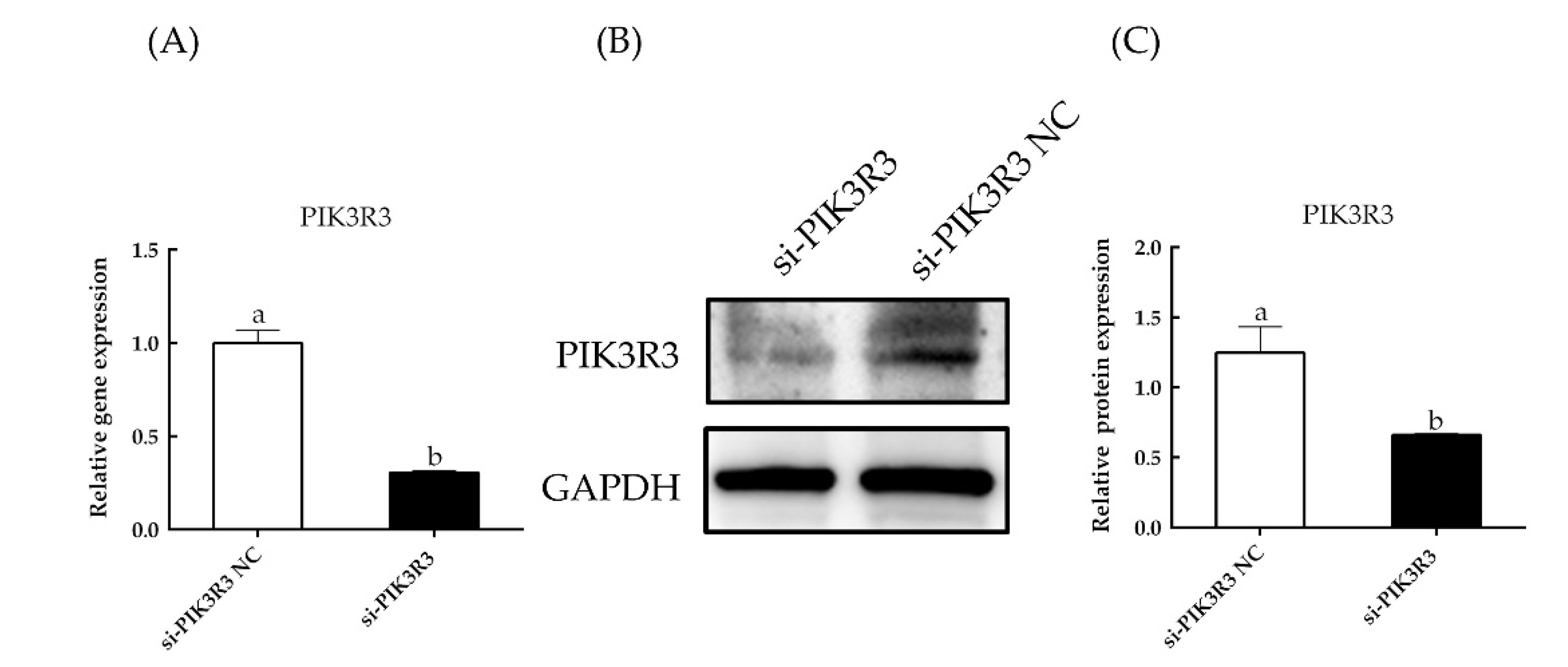
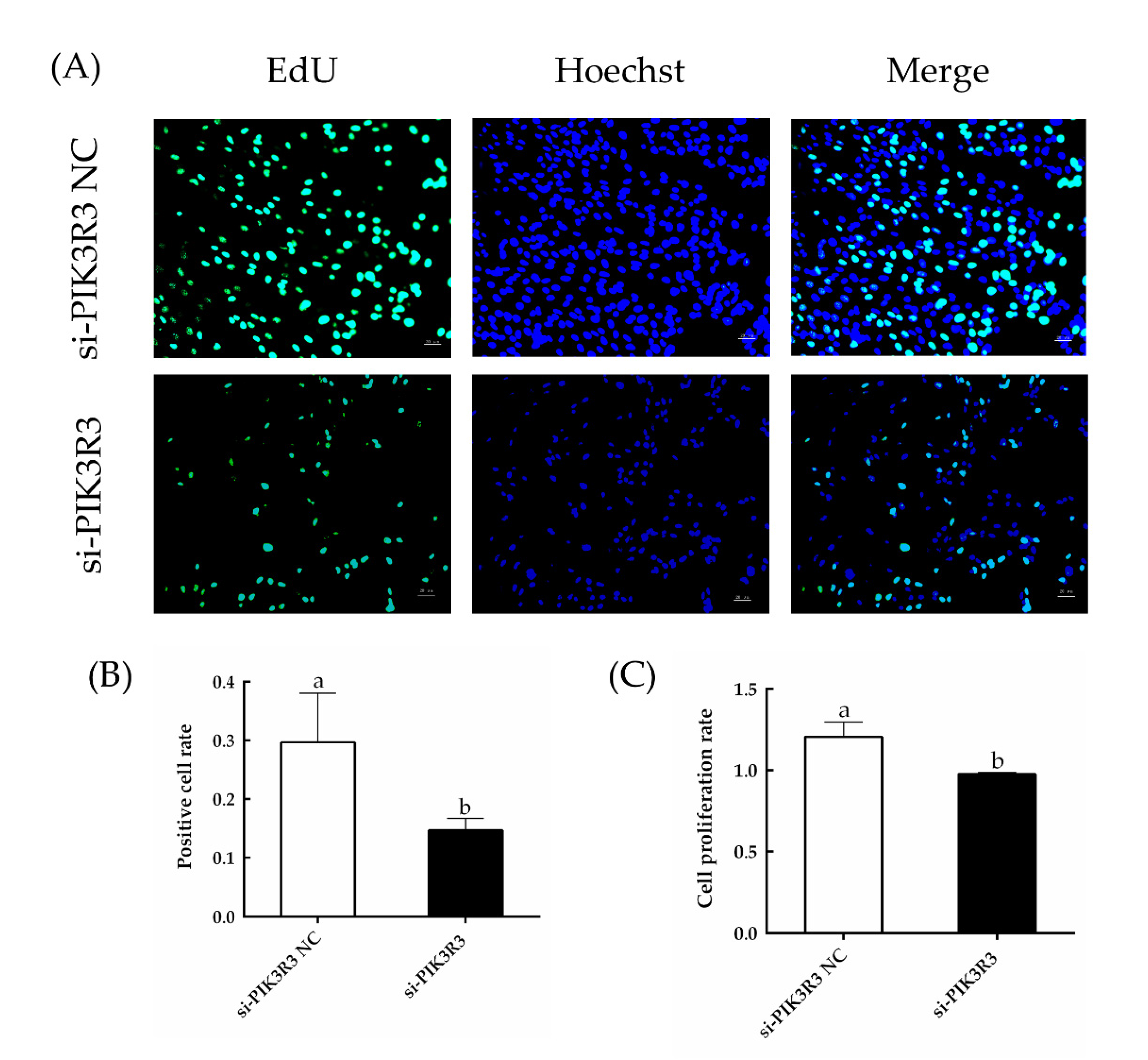
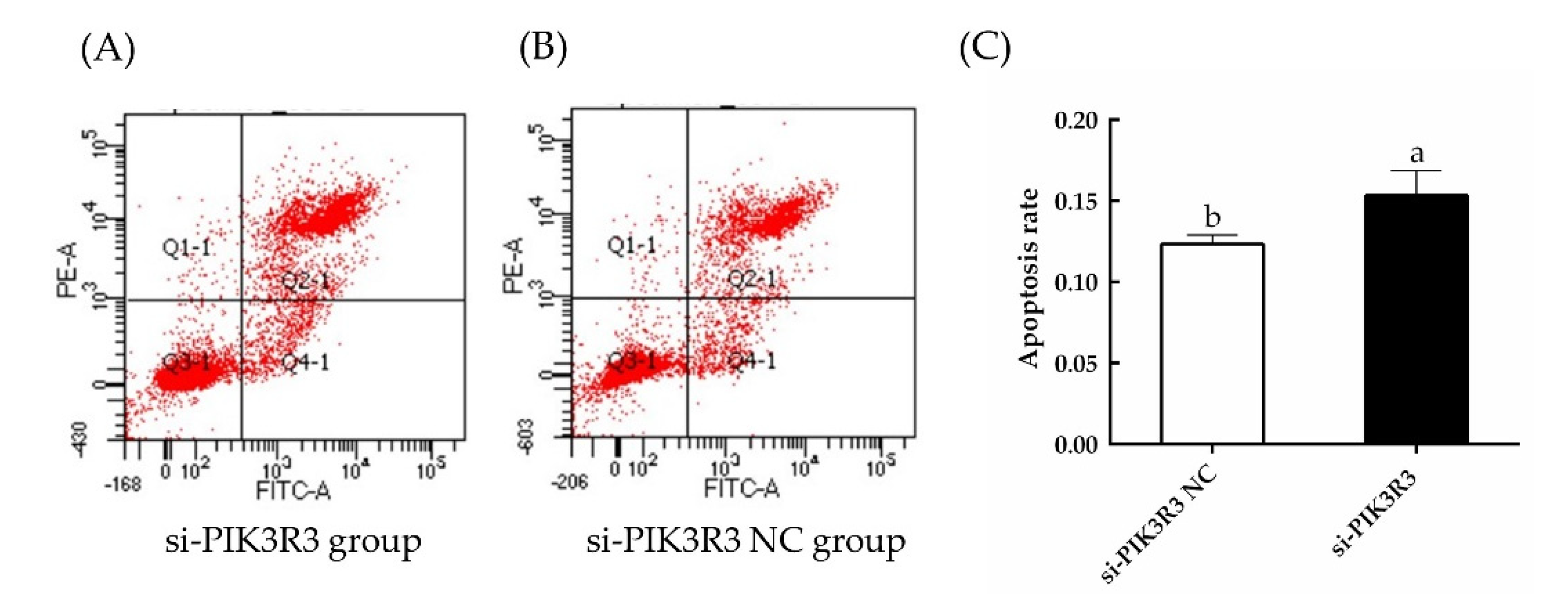
3.1. PIK3R3 Regulates HFSC Proliferation and Apoptosis
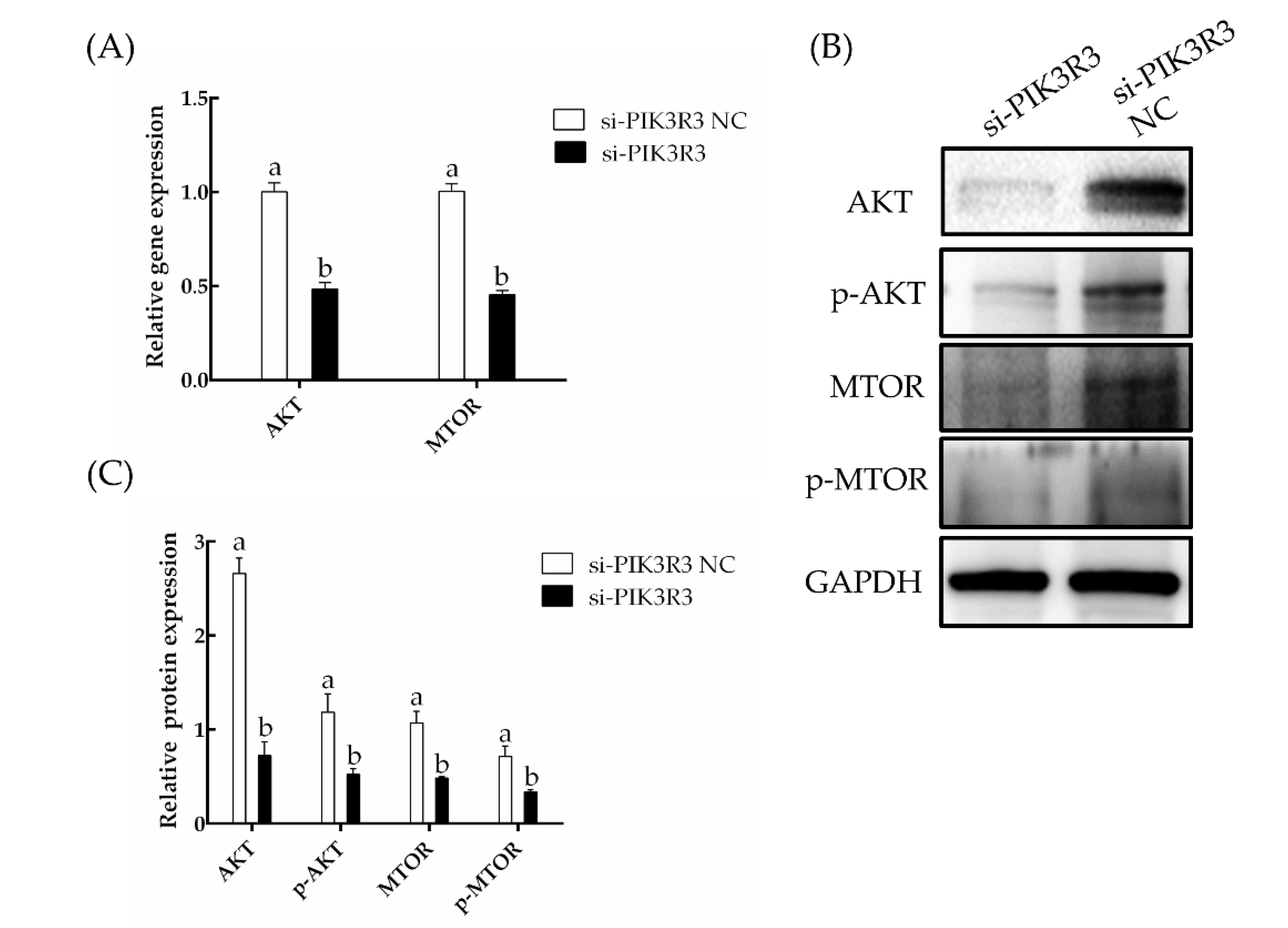
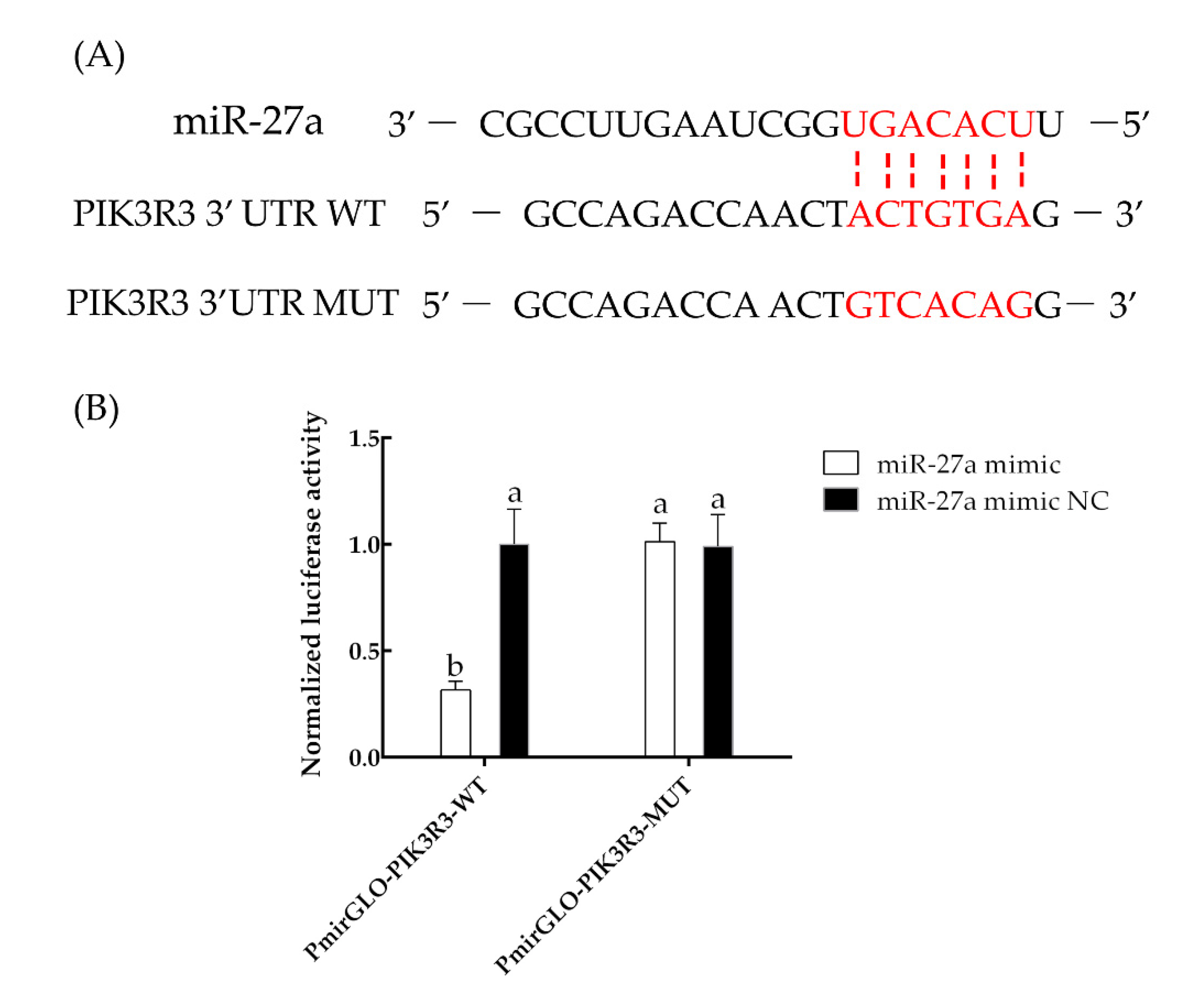
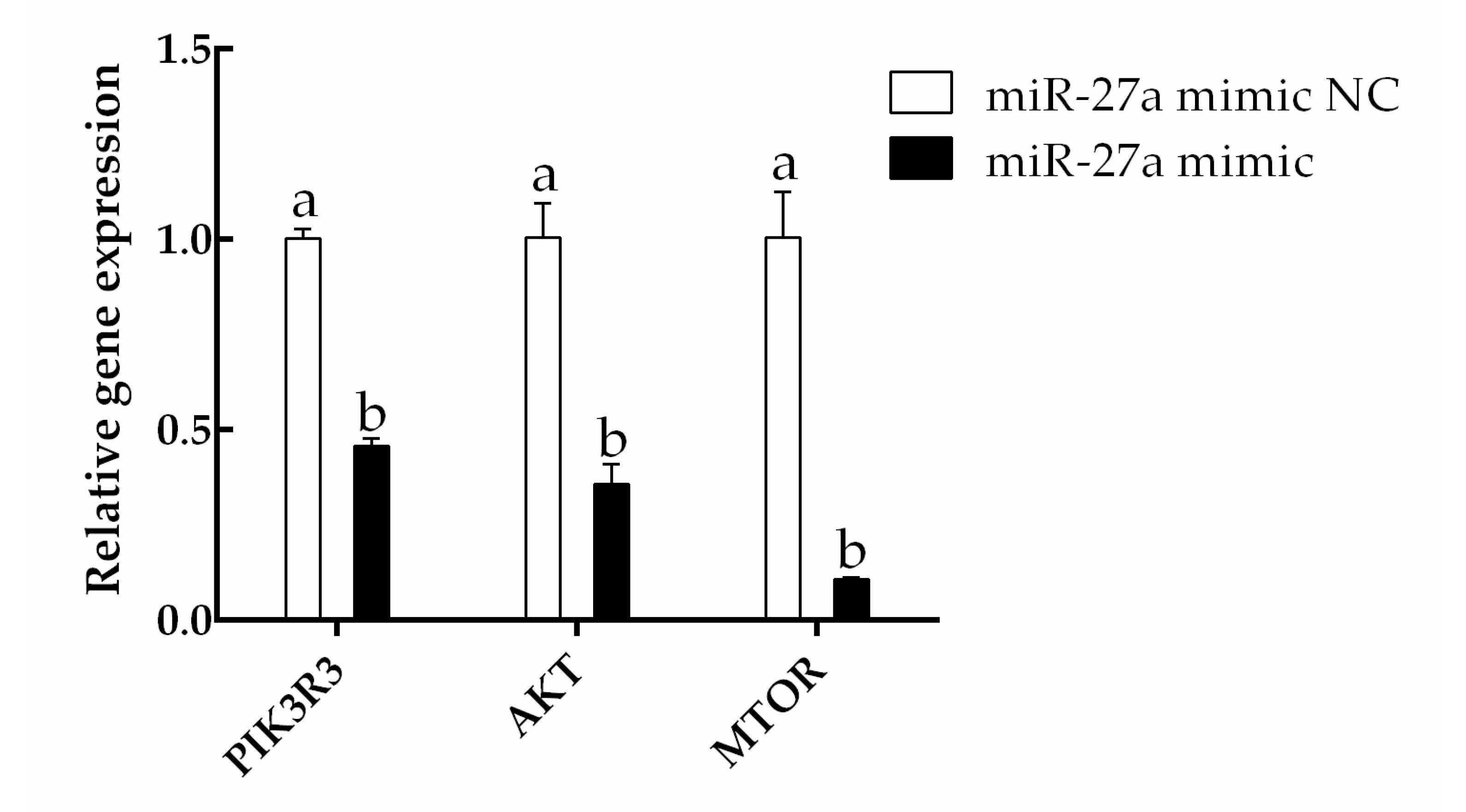
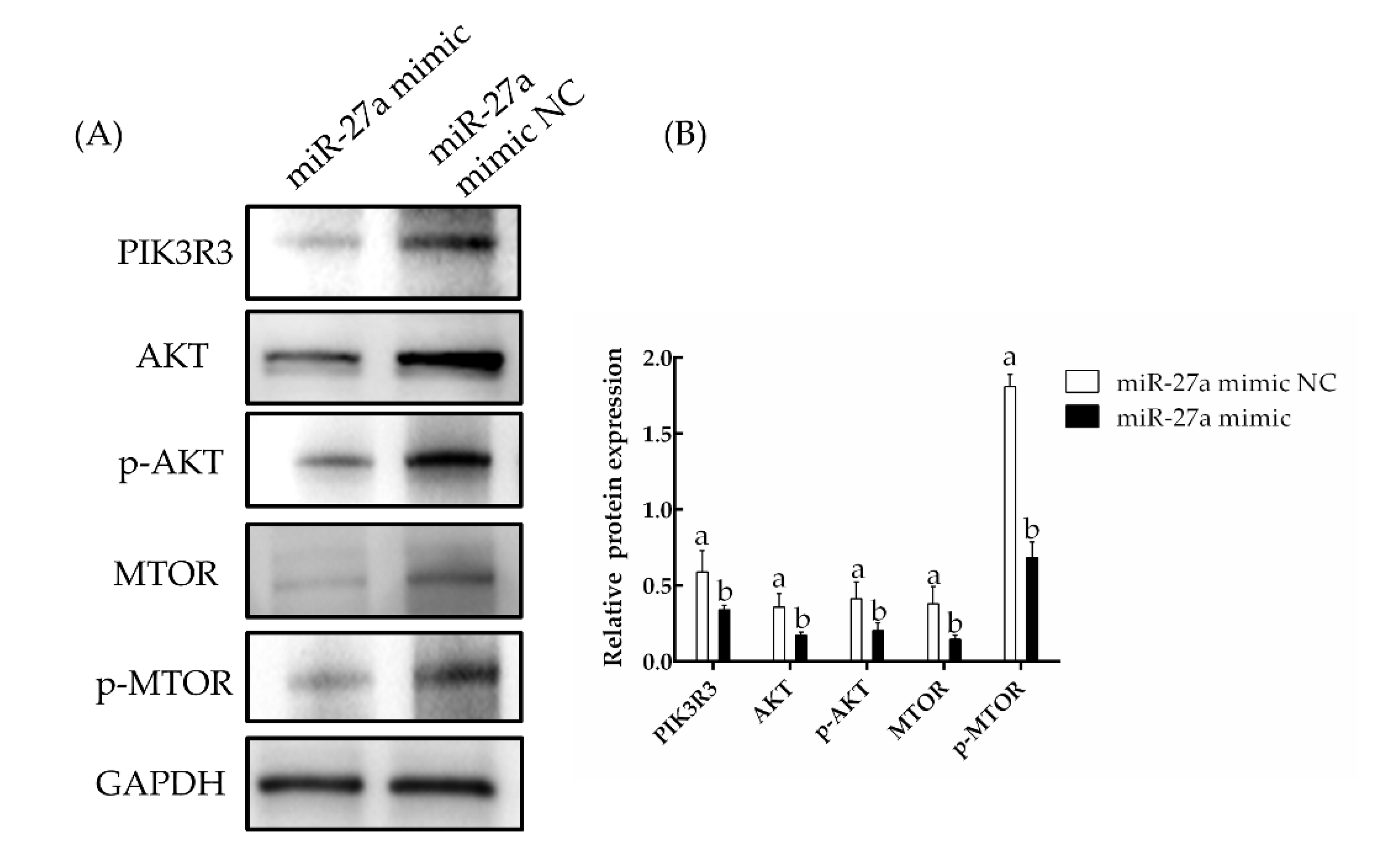
3.2. The miRNA miR-27a Targets PIK3R3 and Downregulates the AKT/MTOR Pathway
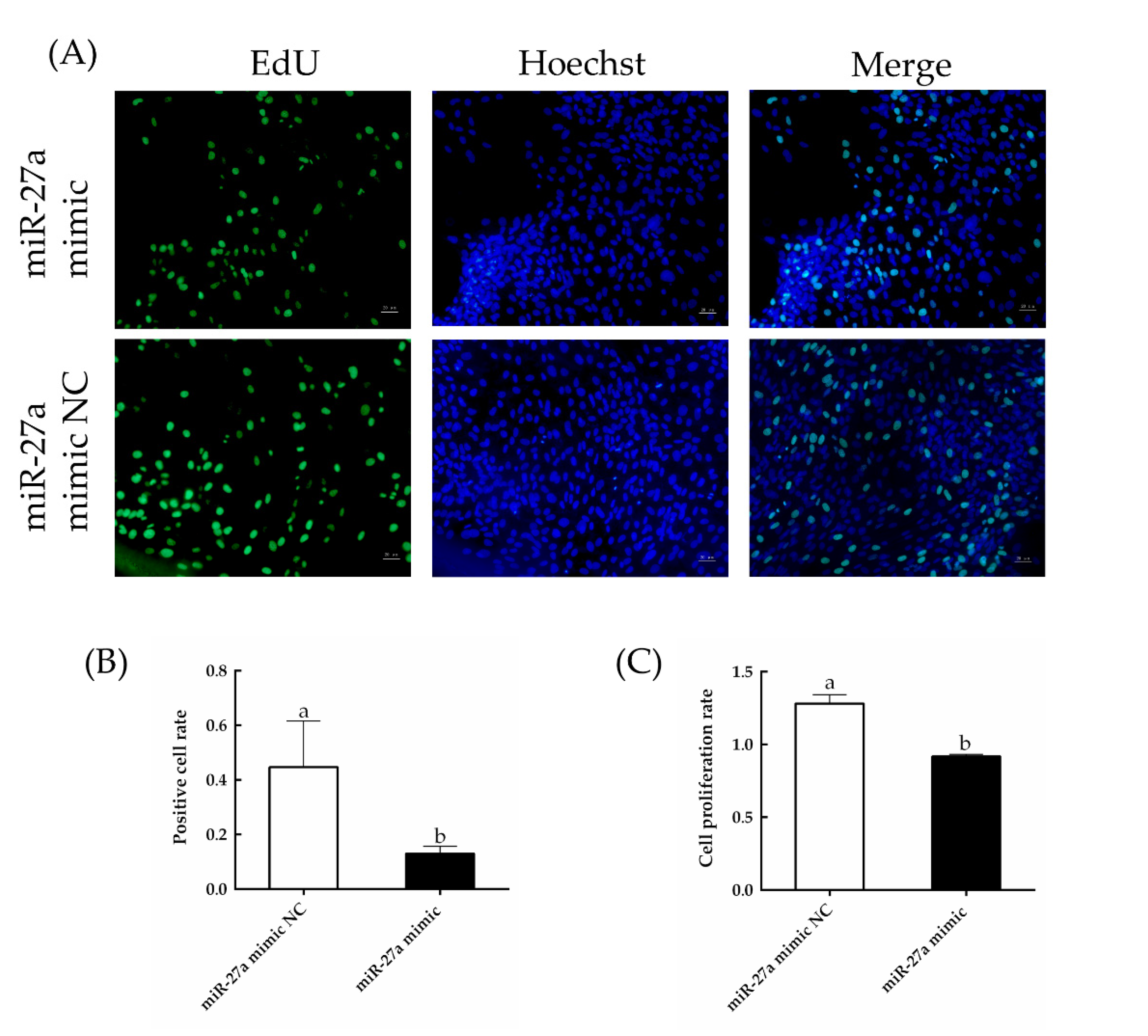
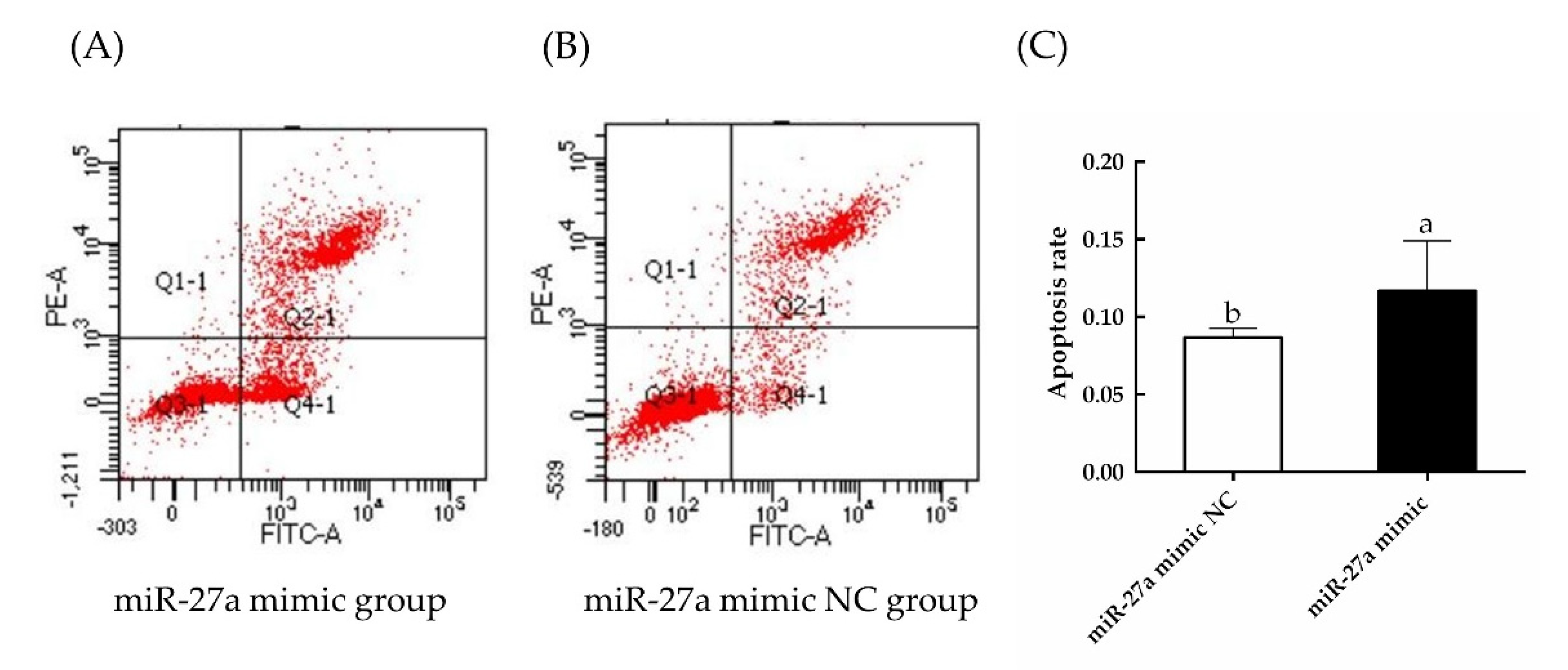
3.3. Overexpression of miR-27a Inhibits HFSC Proliferation
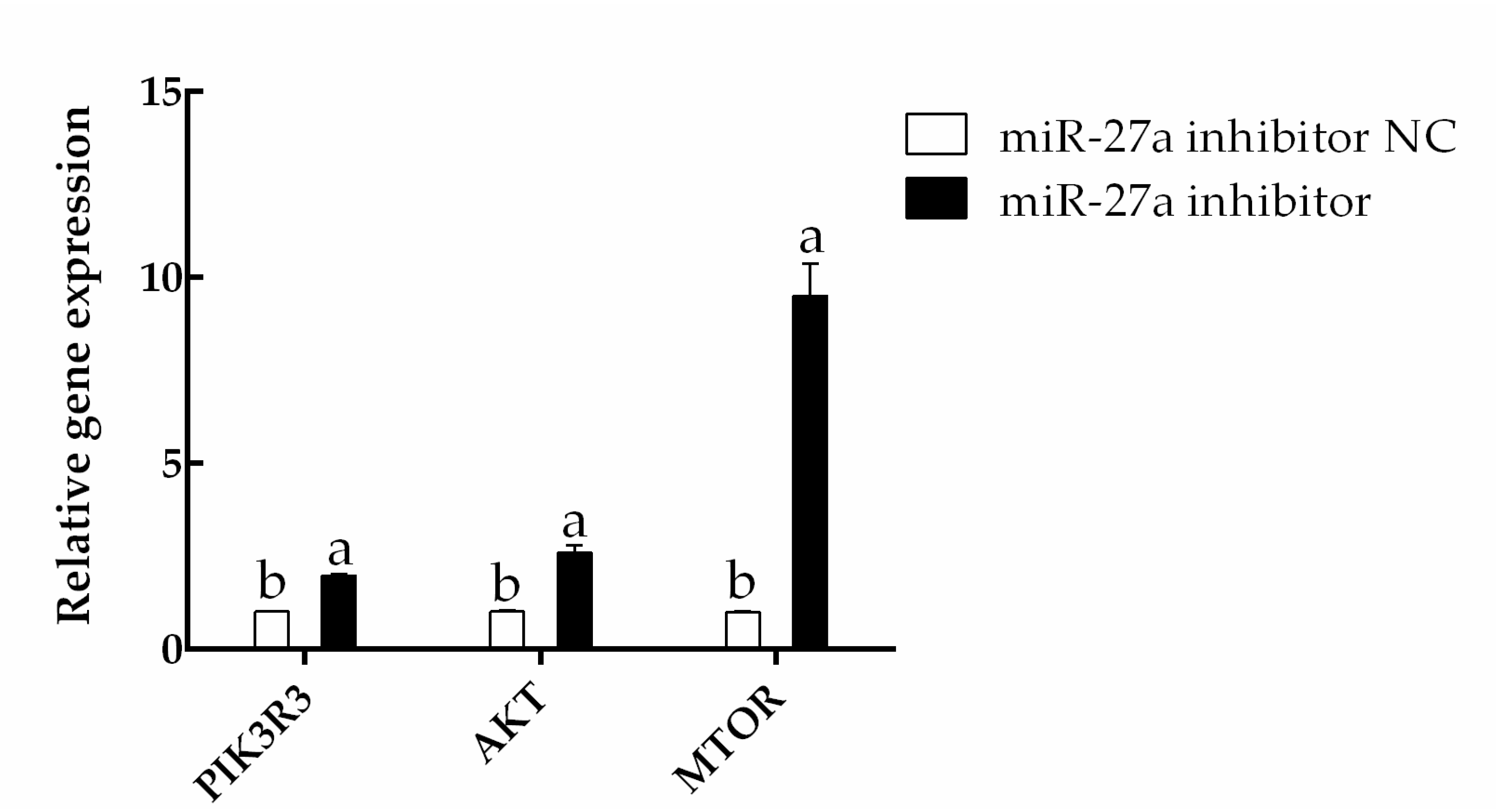
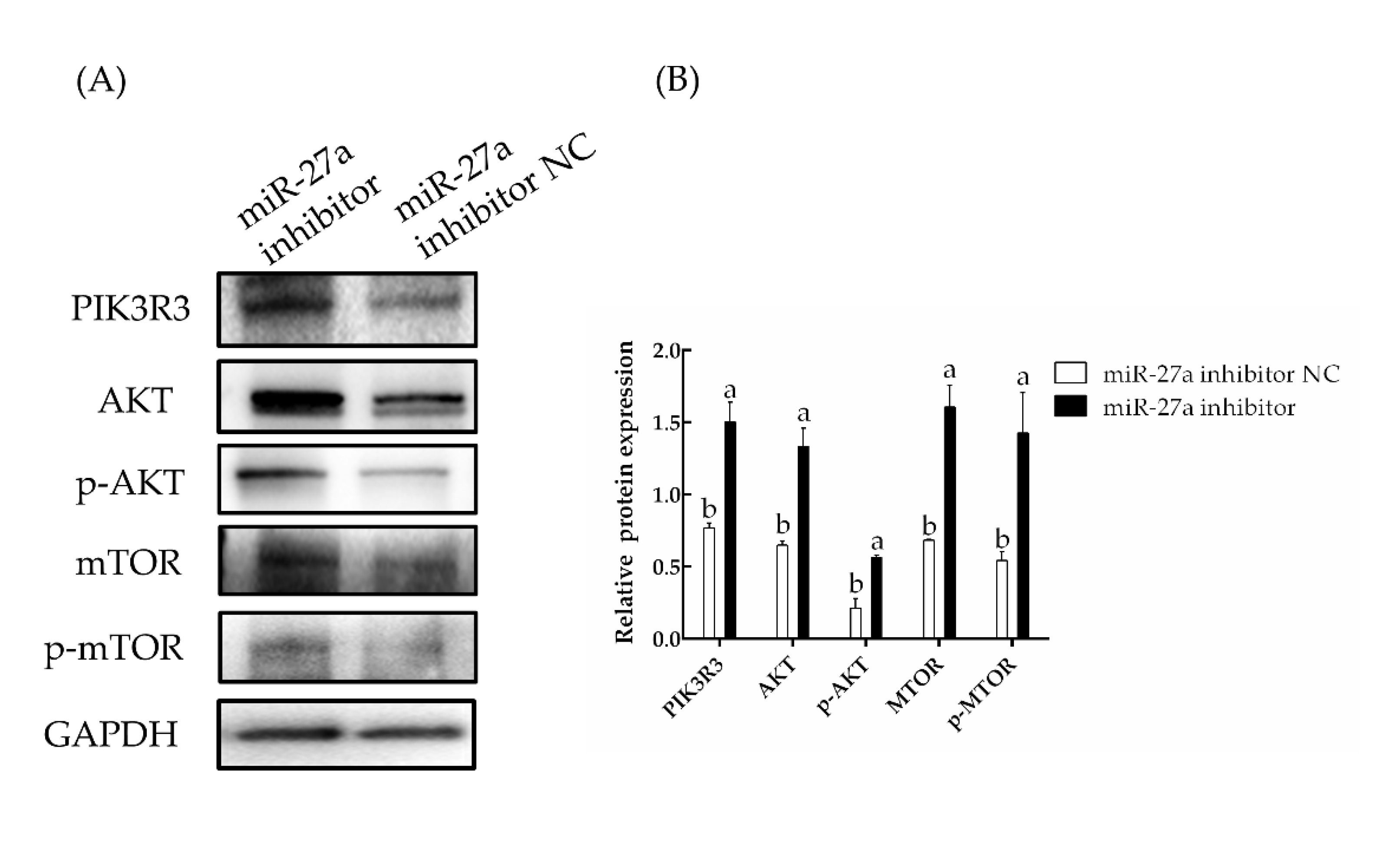
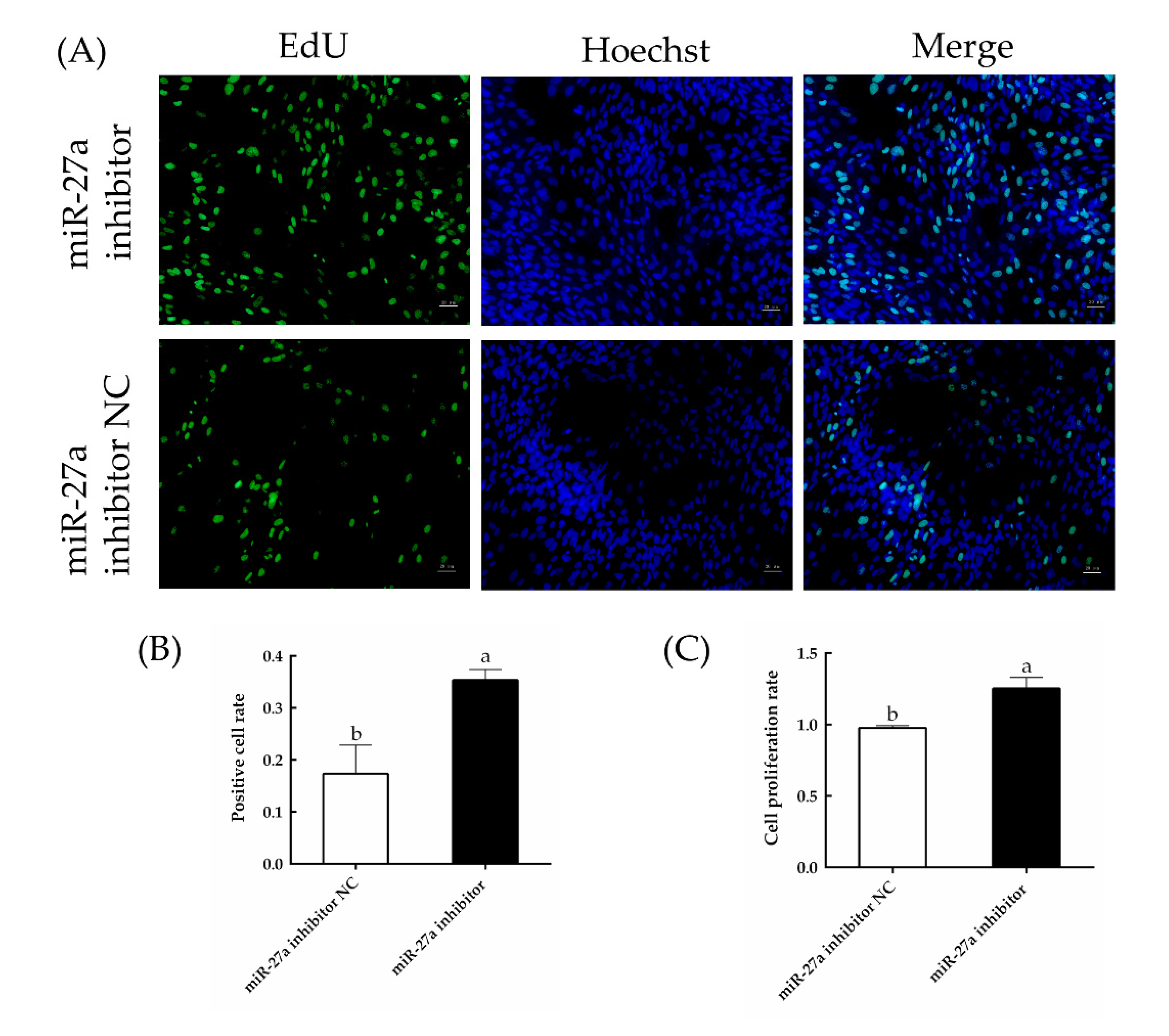
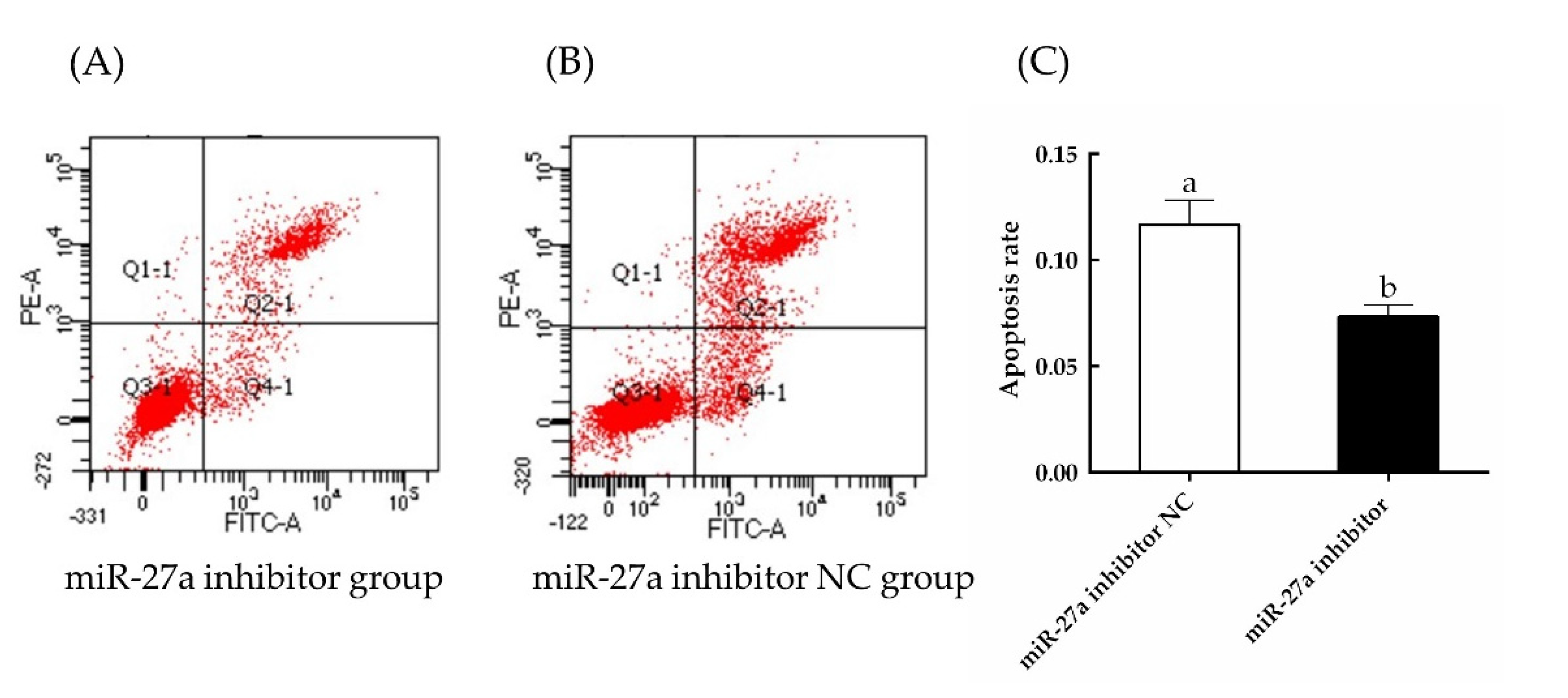
3.4. Interfering with miR-27a Promotes HFSC Proliferation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rogers, G.E. Biology of the wool follicle: An excursion into a unique tissue interaction system waiting to be re-discovered. Exp. Dermatol. 2006, 15, 931–949. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, S.; Zhao, H.; Qiao, J.; Liu, S.; Deng, Z.; Lei, X.; Ning, L.; Cao, Y.; Zhao, Y.; et al. Ovine Hair Follicle Stem Cells Derived from Single Vibrissae Reconstitute Haired Skin. Int. J. Mol. Sci. 2015, 16, 17779–17797. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Li, J.; Liu, N.; Li, H.; Liu, L.; Yang, F.; Li, L.; Wang, Y.; He, J. Transcriptomic Analysis Reveals the Involvement of lncRNA-miRNA-mRNA Networks in Hair Follicle Induction in Aohan Fine Wool Sheep Skin. Front. Genet. 2020, 11, 590. [Google Scholar] [CrossRef]
- Hwang, D.; Lee, H.; Lee, J.; Lee, M.; Cho, S.; Kim, T.; Kim, H. Micro-Current Stimulation Has Potential Effects of Hair Growth-Promotion on Human Hair Follicle-Derived Papilla Cells and Animal Model. Int. J. Mol. Sci. 2021, 22, 4361. [Google Scholar] [CrossRef]
- Huang, S.; Zhen, Y.; Yin, X.; Yang, Z.; Li, X.; Wang, R.; Wen, H.; Zhong, H.; Yan, J.; Sun, Q. KMT2C Induced by FABP5P3 Aggravates Keratinocyte Hyperproliferation and Psoriasiform Skin Inflammation by Upregulating the Transcription of PIK3R3. J. Investig. Dermatol. 2022, 143, 37–47.e8. [Google Scholar] [CrossRef]
- Ji, Z.H.; Chen, J.; Gao, W.; Zhang, J.Y.; Quan, F.S.; Hu, J.P.; Yuan, B.; Ren, W.Z. Cutaneous transcriptome analysis in NIH hairless mice. PLoS ONE 2017, 12, e0182463. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, Y.; Liu, Y.; Zhu, D.; Yu, J.; Li, G.; Sun, Z.; Wang, W.; Jiang, H.; Hong, Z. MiR-27a promotes insulin resistance and mediates glucose metabolism by targeting PPAR-γ-mediated PI3K/AKT signaling. Aging 2019, 11, 7510–7524. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Blackstone, B.N.; Wilgus, T.A.; Roy, S.; Wulff, B.C.; Powell, H.M. Skin Biomechanics and miRNA Expression Following Chronic UVB Irradiation. Adv. Wound Care 2020, 9, 79–89. [Google Scholar] [CrossRef]
- Cai, T.; Liu, Z.H.; Wang, Z.X.; Zhao, M.; Ju, H.L.; Li, J.Q. miRNA in regulation of skin and hair follicle development. Yi Chuan = Hered. 2013, 35, 1087–1094. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, M.; Liu, Y.; Zhang, J.; Lai, W.; Xu, Q.; Zheng, Y. Predicting miRNA-lncRNA-mRNA network in ultraviolet A-induced human skin photoaging. J. Cosmet. Dermatol. 2021, 20, 1875–1884. [Google Scholar] [CrossRef]
- Luo, Z.; Dou, J.; Xie, F.; Lu, J.; Han, Q.; Zhou, X.; Kong, J.; Chen, D.; Liu, A. miR-203a-3p promotes loureirin A-induced hair follicle stem cells differentiation by targeting Smad1. Anat. Rec. 2021, 304, 531–540. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, J.; Ma, J.; Zhang, L.; Chu, C.; Hu, H.; Wang, Y.; Li, Y. miR-31-5p promotes proliferation and inhibits apoptosis of goat hair follicle stem cells by targeting RASA1/MAP3K1 pathway. Exp. Cell Res. 2021, 398, 112441. [Google Scholar] [CrossRef]
- Wang, J.; Wu, X.; Zhang, L.; Wang, Q.; Qu, J.; Wang, Y.; Ji, D.; Li, Y. MiR-149-5p promotes β-catenin-induced goat hair follicle stem cell differentiation. Vitr. Cell. Dev. Biol. Anim. 2022, 58, 325–334. [Google Scholar] [CrossRef]
- Ge, M.; Liu, C.; Li, L.; Lan, M.; Yu, Y.; Gu, L.; Su, Y.; Zhang, K.; Zhang, Y.; Wang, T.; et al. miR-29a/b1 Inhibits Hair Follicle Stem Cell Lineage Progression by Spatiotemporally Suppressing WNT and BMP Signaling. Cell Rep. 2019, 29, 2489–2504.e2484. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, M.; Ding, L.; Tang, J. MiR-27a: A Novel Biomarker and Potential Therapeutic Target in Tumors. J. Cancer 2019, 10, 2836–2848. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Kang, N.; Ling, X.; Pan, M.; Du, W.; Gao, S. MiR-27a-3p Promotes Non-Small Cell Lung Cancer Through SLC7A11-Mediated-Ferroptosis. Front. Oncol. 2021, 11, 759346. [Google Scholar] [CrossRef]
- Li, W.; Zhu, Q.; Xu, X.; Hu, X. MiR-27a-3p suppresses cerebral ischemia-reperfusion injury by targeting FOXO1. Aging 2021, 13, 11727–11737. [Google Scholar] [CrossRef]
- Lu, H.; Liu, P.; Pang, Q. MiR-27a-3p/miR-27b-3p Promotes Neurofibromatosis Type 1 via Targeting of NF1. J. Mol. Neurosci. MN 2021, 71, 2353–2363. [Google Scholar] [CrossRef]
- Li, Y.; Ren, S.; Xia, J.; Wei, Y.; Xi, Y. EIF4A3-Induced circ-BNIP3 Aggravated Hypoxia-Induced Injury of H9c2 Cells by Targeting miR-27a-3p/BNIP3. Mol. Ther. Nucleic Acids 2020, 19, 533–545. [Google Scholar] [CrossRef]
- Cai, C.; Min, S.; Yan, B.; Liu, W.; Yang, X.; Li, L.; Wang, T.; Jin, A. MiR-27a promotes the autophagy and apoptosis of IL-1β treated-articular chondrocytes in osteoarthritis through PI3K/AKT/mTOR signaling. Aging 2019, 11, 6371–6384. [Google Scholar] [CrossRef]
- Tang, J.; Yu, H.; Wang, Y.; Duan, G.; Wang, B.; Li, W.; Zhu, Z. miR-27a promotes osteogenic differentiation in glucocorticoid-treated human bone marrow mesenchymal stem cells by targeting PI3K. J. Mol. Histol. 2021, 52, 279–288. [Google Scholar] [CrossRef]
- Li, C.; Lin, X.F.; Wang, J.N.; Ren, X.S. FBXW7 inhibited cell proliferation and invasion regulated by miR-27a through PI3K/AKT signaling pathway and epithelial-to-mesenchymal transition in oral squamous cell carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3701–3709. [Google Scholar] [CrossRef]
- Wu, S.; Li, J.; Ma, T.; Li, J.; Li, Y.; Jiang, H.; Zhang, Q. MiR-27a regulates WNT3A and KITLG expression in Cashmere goats with different coat colors. Anim. Biotechnol. 2021, 32, 205–212. [Google Scholar] [CrossRef]
- Wang, Q.; Qu, J.; Li, Y.; Ji, D.; Zhang, H.; Yin, X.; Wang, J.; Niu, H. Hair follicle stem cells isolated from newborn Yangtze River Delta White Goats. Gene 2019, 698, 19–26. [Google Scholar] [CrossRef]
- Li, B.; Huang, X.; Yang, C.; Ge, T.; Zhao, L.; Zhang, X.; Tian, L.; Zhang, E. miR-27a Regulates Sheep Adipocyte Differentiation by Targeting CPT1B Gene. Animals 2021, 12, 28. [Google Scholar] [CrossRef]
- Zhao, B.; Luo, H.; He, J.; Huang, X.; Chen, S.; Fu, X.; Zeng, W.; Tian, Y.; Liu, S.; Li, C.J.; et al. Comprehensive transcriptome and methylome analysis delineates the biological basis of hair follicle development and wool-related traits in Merino sheep. BMC Biol. 2021, 19, 197. [Google Scholar] [CrossRef]
- Snippert, H.J.; Haegebarth, A.; Kasper, M.; Jaks, V.; van Es, J.H.; Barker, N.; van de Wetering, M.; van den Born, M.; Begthel, H.; Vries, R.G.; et al. Lgr6 marks stem cells in the hair follicle that generate all cell lineages of the skin. Science 2010, 327, 1385–1389. [Google Scholar] [CrossRef] [Green Version]
- Coutte, L.; Dreyer, C.; Sablin, M.P.; Faivre, S.; Raymond, E. PI3K-AKT-mTOR pathway and cancer. Bull. Du Cancer 2012, 99, 173–180. [Google Scholar] [CrossRef]
- Hessam, S.; Gambichler, T.; Skrygan, M.; Scholl, L.; Sand, M.; Meyer, T.; Stockfleth, E.; Bechara, F.G. Increased expression profile of NCSTN, Notch and PI3K/AKT3 in hidradenitis suppurativa. J. Eur. Acad. Dermatol. Venereol. JEADV 2021, 35, 203–210. [Google Scholar] [CrossRef]
- Mercurio, L.; Albanesi, C.; Madonna, S. Recent Updates on the Involvement of PI3K/AKT/mTOR Molecular Cascade in the Pathogenesis of Hyperproliferative Skin Disorders. Front. Med. 2021, 8, 665647. [Google Scholar] [CrossRef]
- Strozyk, E.; Kulms, D. The role of AKT/mTOR pathway in stress response to UV-irradiation: Implication in skin carcinogenesis by regulation of apoptosis, autophagy and senescence. Int. J. Mol. Sci. 2013, 14, 15260–15285. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Cheng, A.; Akinmade, D.; Hamburger, A.W. The N-terminal 24 amino acids of the p55 gamma regulatory subunit of phosphoinositide 3-kinase binds Rb and induces cell cycle arrest. Mol. Cell. Biol. 2003, 23, 1717–1725. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.B.; Lu, Y.S.; Liu, T.; Li, L.M.; Wang, H.X.; Wu, Y.; Gao, X.H.; Chen, H.D. UVA influenced the SIRT1-miR-27a-5p-SMAD2-MMP1/COL1/BCL2 axis in human skin primary fibroblasts. J. Cell. Mol. Med. 2020, 24, 10027–10041. [Google Scholar] [CrossRef]
- Wang, H.; Wang, S.; Chen, F.; Zhang, C.; Zhang, X.; Sun, Y. Expression and clinical value of miR-27a in serum of patients with skin squamous cell carcinoma. J. BU ON. Off. J. Balk. Union Oncol. 2020, 25, 2515–2522. [Google Scholar]
- Banerjee, N.; Das, S.; Tripathy, S.; Bandyopadhyay, A.K.; Sarma, N.; Bandyopadhyay, A.; Giri, A.K. MicroRNAs play an important role in contributing to arsenic susceptibility in the chronically exposed individuals of West Bengal, India. Environ. Sci. Pollut. Res. Int. 2019, 26, 28052–28061. [Google Scholar] [CrossRef]
- Grabarek, B.O.; Dąbala, M.; Kasela, T.; Gralewski, M.; Gładysz, D. Changes in the Expression Pattern of DUSP1-7 and miRNA Regulating their Expression in the Keratinocytes Treated with LPS and Adalimumab. Curr. Pharm. Biotechnol. 2022, 23, 873–881. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, J.; Liu, Y.; Chen, Z.; Li, X.; Tang, L.; Li, J.; Duan, M.; Zhang, G. Human Amniotic Fluid Stem Cell-Derived Exosomes as a Novel Cell-Free Therapy for Cutaneous Regeneration. Front. Cell Dev. Biol. 2021, 9, 685873. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′-3′) | Gene ID | Product Size (bp) |
|---|---|---|---|
| PIK3R3 | F: AGACTGGAGGGAGGTGATG | 101123247 | 100 |
| R: AGTCATTGGCTTAGGTGGC | |||
| AKT | F: GTCGCCCCTCAACAACTTCT | 100294652 | 233 |
| R: CATCGTCTCCTCCTCCTCCTGCC | |||
| MTOR | F: AGCATCTCTCCCCAAAGAACCTCA | 100271659 | 221 |
| R: GGCCCTGGTCTCTTCATTCC | |||
| GAPDH | F: AAGTTCAACGGCACAGTCA | 443005 | 151 |
| R: ACCACATACTCAGCACCAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.; Li, L.; Liu, M.; Wang, L.; Gao, X.; Zhou, L.; Liu, N.; He, J. miR-27a Targeting PIK3R3 Regulates the Proliferation and Apoptosis of Sheep Hair Follicle Stem Cells. Animals 2023, 13, 141. https://doi.org/10.3390/ani13010141
Yu M, Li L, Liu M, Wang L, Gao X, Zhou L, Liu N, He J. miR-27a Targeting PIK3R3 Regulates the Proliferation and Apoptosis of Sheep Hair Follicle Stem Cells. Animals. 2023; 13(1):141. https://doi.org/10.3390/ani13010141
Chicago/Turabian StyleYu, Mengqi, Lanlan Li, Meng Liu, Lei Wang, Xiaoxiao Gao, Lisheng Zhou, Nan Liu, and Jianning He. 2023. "miR-27a Targeting PIK3R3 Regulates the Proliferation and Apoptosis of Sheep Hair Follicle Stem Cells" Animals 13, no. 1: 141. https://doi.org/10.3390/ani13010141